不同葉面積指數對“黃果柑”光合特性及果實品質的影響
2019-07-24 12:29:28孫國超汪志輝
四川農業大學學報 2019年3期
關鍵詞:影響
邱 霞,孫國超,熊 博,廖 玲,汪志輝,*,王 迅,祝 進
(1.四川農業大學園藝學院,成都 611130;2.四川農業大學果蔬研究所,成都 611130;3.四川省園藝作物技術推廣總站,成都 610041)
葉面積指數(leaf area index,LAI),即植物葉片表面積之和與土地表面積的比率[1],是定量描述群體水平上葉片的空間分布和密度變化的特征參數[2],受植物大小、年齡、株行距和其他因子的影響[3]。LAI能直接影響植物群體對光能的截獲及利用[4]、水分的吸收與蒸騰[5]、潛熱和感熱通量等[6],進而影響群體的生長發育[7]、光合作用[8]和蒸騰作用[9]等生理過程,是反映作物產量[10]、植被冠層結構及生長健康狀況[11]的重要指標[12],同時也是水文、生態、氣候等模型的重要輸入參數[13],其值大小對農業生產具有重要意義。
光合作用是作物產量和品質形成的基礎,LAI作為重要的功能性狀,與植物植被冠層的截獲、冠層表明能量交換和凈初級生產力具有較強的相關關系[14],對作物群體的光合作用以及整個生長過程有重要的影響[15]。冠層LAI 大小能反映植物利用太陽輻射能量進行光合作用的能力,在一定范圍內,植物群體光合作用能力會著LAI 的增加而提升,而當LAI 達到一定限度后,植物葉片交叉面積過大,易導致田間郁閉,光照強度減弱,光合作用速率減小,進而作物產量降低[16]。目前關于LAI 的研究主要在森林、草地和大田作物等生態系統[17],研究方向集中在LAI 的估算、LAI 與產量的關系上,而LAI 對果樹光合作用和果實品質的影響鮮有研究,所以研究LAI 對柑橘葉片光合作用能力及果實品質的影響,對指導農業生產具有重要意義。
同等樹齡的柑橘的LAI 主要受栽植密度和修剪程度的影響,目前栽植密度對植物光合作用的影響研究較多[18],但栽植密度不能完全定量描述冠層葉片數量,其忽略了修剪程度對葉片數量的影響,故研究LAI 對光合作用的影響更為全面而精確。本試驗以黃果柑為試材,設置 LAI 為 2.0、2.4、2.8、3.2和3.6 共5 個處理,對各處理的葉片光合特性及果實品質指標進行測定與分析,篩選出適宜黃果柑的LAI,使黃果柑獲得高光效,進而提高其果實品質,并可為其他柑橘品種LAI 的選擇提供理論基礎。
1 材料和方法
1.1 試驗材料與地點
試驗材料為7年生黃果柑果樹,供試植株樹勢基本一致,生長良好,無病蟲害。試驗園區在四川省雅安市石棉縣安順片區的黃果柑栽培示范園,海拔850 m 左右,年均溫 13~17 ℃,1月均溫 5~8 ℃,絕對低溫-1 ℃,年有效積溫 4 000~6 500 ℃,無霜期250~300 d,沙壤土,有機質含量 1.79%,pH 值為 6.5左右。
1.2 試驗設計
按LAI 的不同,設置表1所示的5 個處理,LAI的設置通過栽植密度和修剪水平的調整實現,各處理于夏季和冬季各修剪1 次,使全年LAI 變化穩定在正負0.1 之間,LAI 用LAI-2200 植物冠層分析儀測定[19]。小區面積 667 m2,3 次重復,小區間隔 10 m以上。各處理除修剪外的田間管理一致,施肥量以單株計算,以減少肥水條件對試驗的影響。

表1 試驗處理列表Table 1 The lest of treatment
1.3 測定方法
1.3.1 光合氣體交換參數測定
在幼果期、膨大期、轉色期和成熟期這4 個關鍵物候期進行黃果柑葉片光合氣體交換參數測定。測定時間為晴朗天氣的 8:00~18:00,測定儀器為Li-COR 6400XTP 型便攜式光合作用測量系統,選擇樹冠南面新梢頂部第3~5 片健康功能葉進行測定,每2 h 測定一組,每個數據重復測定3 次,自動記錄數據。測定參數主要包括凈光合速率(Pn)、蒸騰速率(Tr)、胞間 CO2濃度(Ci)和氣孔導度(Gs)。記錄數據并繪制成圖。
1.3.2 光響應曲線、CO2響應曲線測定
參照邱霞[20]的方法對光響應曲線和CO2響應曲線進行測定,測定時間為葉幕穩定期晴朗天氣的9:00~11:00,測定儀器同 1.3.1。光響應曲線采用 LED紅藍光源控光測定,光強依次設定為2 000、1 800、1 500、1 200、1 000、800、600、400、200、100、50、30、0 μmol/m2·s,樣品室CO2濃度設定為400 μmol/m2·s,采用LightCurve2 程序自動測定,記錄,繪制成圖。CO2響應曲線采用CO2壓縮鋼瓶緩釋控制測定,樣品室 CO2濃度依次設定為 400、300、200、100、50、400、400、600、800、1 000、1 200、1 500、2 000 μmol/m2·s,光強設定為 1 200 μmol/m2·s,采用 A-Ci Curve2 程序自動測定,每個處理重復3 次,記錄,繪制成圖。
1.3.3 葉綠素熒光參數的測定
葉綠素熒光參數的測定儀器為PAM 2500 型便攜式調制熒光儀,測定時間為葉幕穩定期無雨天氣的黎明前,植株經過充分的暗適應,參照隆春艷[21]的操作測定初始熒光(F0)和最大熒光值(Fm),每個處理重復 3 次。依照公式 Fv/F0=(Fm-F0)/F0計算PSII潛在活性,并依照公式 Fv/Fm=(Fm-F0)/Fm計算 PSII最大光化學轉化效率Fv/Fm,整理記錄數據。
規劃核實測量成果數據是進行規劃核實的依據,具有嚴肅的法定性。為此,核實測量的全過程從外業數據采集、內業數據處理及質量檢查都必須細致、準確。核實測量的各個工序都完成之后,需要每一個作業班組織本班的人員進行自己負責工序的自檢、互檢;再由核實測量的技術負責人和專職的檢查人員對班組提交的成果進行細致全面的檢查,并進行外業的質量檢查工作。為確保質量,還準備適時實施注冊測繪師簽字制度。
1.3.4 黃果柑果實品質測定
用游標卡尺測量果實縱、橫徑,計算果形指數;用AL204 型精密電子天平測定單果重;用TD-45型數顯式手持測糖儀測定可溶性固形物含量;用蒽酮比色法測定糖總含量[22];用酸堿中和滴定法測定可滴定酸含量[22];用鉬藍比色法測定Vc 含量[22]。每個處理每個指標重復測定3 次。
1.4 數據分析
用光合計算軟件以直角雙曲線的修正模型[23]對光響應和CO2響應曲線的實驗數據進行處理,用SPSS 22.0 軟件以 Duncar 新復極差法做 P<0.05 水平的方差差異顯著性分析,并用Microsoft Excel 2016對數據進行整理和作圖。
2 結果與分析
2.1 葉面積指數對黃果柑光合氣體交換參數的影響
由圖1可知,黃果柑葉片凈光合速率(Pn)在幼果期和成熟期時呈升-降-升-降的雙峰曲線,因為這兩個時期時溫度較高光照較強,存在不同程度的光合抑制;果實膨大期和轉色期呈先升后降的單峰曲線,沒有明顯的光合抑制現象,在早上10:00 或12:00 達到峰值。不同 LAI 處理下黃果柑的 Pn相比,處理5 的Pn在4 個物候期中均較低,且日變化相對較穩定;處理1 在4 個物候期的Pn最大值分別為12.75、7.49、8.29 和 3.5 μmol/m2·s,比同期處理 5 的Pn高 1.02、1.76、2.16 和 2.89 倍,光合速率顯著提升。
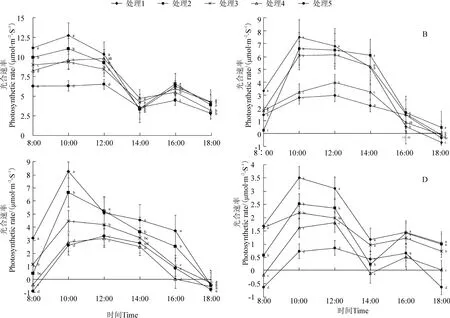
圖1 不同葉面積指數處理的黃果柑葉片光合速率日變化Figure 1 Photosynthetic rate of Huangguogan leaves with different leaf area index
由圖2可知,黃果柑葉片氣孔導度(Gs)日變化規律與Pn相似,幼果期和成熟期呈雙峰曲線,果實膨大期和轉色期呈單峰趨勢,在早上10 點左右達到最高值,12 點下降,較Pn變化早。Gs的年均值表現為處理1>處理2>處理4>處理3>處理5,依次為0.082、0.067、0.060、0.059 和 0.050 mmol/m2·s,整體表現為隨著LAI 的升高而降低,表明低葉片密度的光環境更利于葉片氣孔開張。

圖2 不同葉面積指數處理下黃果柑葉片氣孔導度日變化Figure 2 Stomatal conductance of Huangguogan leaves with different leaf area index
由圖3可知,黃果柑葉片胞間 CO2濃度(Ci)與Pn呈相反規律,幼果期及成熟期呈降-升-降-升趨勢,果實膨大期及轉色期呈先降后升趨勢。不同LAI處理下黃果柑葉片Ci相比,低LAI 處理葉片的Ci較高LAI 處理的Ci低,隨著LAI 的降低,葉片光合作用增強,Pn上升,消耗CO2增多,使Ci顯著低于高LAI處理。處理 1 的 Ci年均值僅為 310.78 μmol/mol,較處理5 低13.1%,但果實膨大期和成熟期的中午,光合抑制現象較強時,處理5、處理4 和處理3 這3 個處理的 Ci較處理 1 和處理 2 的 Ci低,可能是因為葉片密度較大,相互遮陰嚴重,光照和溫度上升較慢,光合抑制現象較弱,所以導致Ci上升不明顯。
由圖4可知,黃果柑葉片蒸騰速率(Tr)日變化整體呈先升后降的趨勢,僅在果實膨大期12-14 點出現了較明顯抑制。Tr年均值表現為處理1>處理2>處理 4>處理 3>處理 5,依次為 1.57、1.25、1.15、1.12 和 1.03 mmol/m2·s,整體表現為隨著 LAI 的降低而升高。可見隨著LAI 的降低,葉片密度下降,葉片間遮擋程度下降,更多的葉片直接暴露在光照條件下,葉溫升高,使葉片蒸騰速率上升。
2.2 葉面積指數對黃果柑葉片光和CO2響應特征的影響

圖3 不同葉面積指數處理下黃果柑Ci日變化Figure 3 Intercellular CO2 concentration of Huangguogan leaves with different leaf area index

圖4 不同葉面積指數處理下黃果柑葉片Tr日變化Figure 4 Transpiration rate of Huangguogan leaves with different leaf area index

圖5 不同葉面積指數處理下黃果柑葉片光響應曲線Figure 5 Light response curve of Huangguogan leaves with different leaf area index
不同LAI 處理下黃果柑葉片光合氣體交換參數對CO2濃度梯度變化的響應關系如圖6所示,5個處理葉片的CO2響應曲線變化速率及最大光合速率均存在較大差異,與光響應曲線相似。所有處理的Pn均隨著CO2濃度的增大整體呈先上升后減緩趨勢,Gs和 Tr隨 PAR 升高逐漸降低,Ci隨 PAR 升高呈線性升高趨勢。在CO2濃度小于500 μmol/mol的區段,Pn隨著CO2濃度的上升迅速升高;當CO2濃度大于500 μmol/mol 時,保持在較穩定水平或以較低速率持續上升,與光響應曲線不同的是,幾乎未出現因CO2濃度過高而受到抑制的現象。對數據擬合分析得到CO2飽和點最大光合速率(CSPn),本試驗測定的 CSPn 值為光強控制在1 200 μmol/m2·s條件下,CO2濃度達到該品種飽和時,葉片的最大光合速率擬合值,能一定程度上反映該品種的光合潛力。不同處理的 CSPn在 24.52~32.26 μmol/m2·s 之間,表現為處理 1>處理 2>處理 3>處理 4>處理 5,依次為 32.26、29.39、28.71、27.52 和 24.54 μmol/m2·s,可見隨著LAI 降低,黃果柑光合潛力提高。
2.3 葉面積指數對黃果柑光合熒光參數的影響
由圖7可知,隨著LAI 的上升,黃果柑葉片F0、Fm和Fv/F0均呈下降趨勢,不同處理間差異顯著,Fv/Fm略有下降,但差異不顯著。Fv/Fm反映了PSII 的最大光化學效率[24],黃果柑的Fv/Fm在不同處理間無顯著差異,穩定在0.75~0.78 之間,但Fv/F0呈顯著下降趨勢,Fv/F0反映了PSII 的潛在活性,處理1 的Fv/F0為3.56,較處理5 高20.6%,說明LAI 變化顯著影響了黃果柑葉片的潛在光合能力,與CO2響應曲線測定結果一致。
2.4 葉面積指數對黃果柑果實品質的影響
2.4.1 葉面積指數對黃果柑果實商品質量的影響
由表2可知,不同LAI 處理的黃果柑果實單果重、果實橫徑、縱徑差異顯著,果形指數變化差異不顯著,維持在 0.97~1.04 之間。不同 LAI 處理下黃果柑單果重在168.3~231.6 g 之間,處理1 黃果柑單果重達231.6 g,較處理5 高37.6%;橫縱徑分別為8.15 和 7.89 cm,較處理 5 高 19.50%和 10.81%,說明低LAI 條件更利于黃果柑果實生長發育和商品質量的提高。
2.4.2 葉面積指數對黃果柑果實營養價值的影響
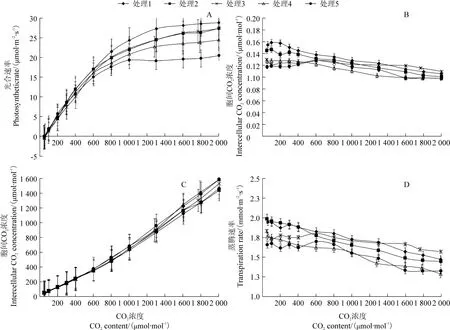
圖6 不同葉面積指數處理下黃果柑葉片CO2 響應曲線Figure 6 CO2 response curve of Huangguogan leaves with different leaf area index
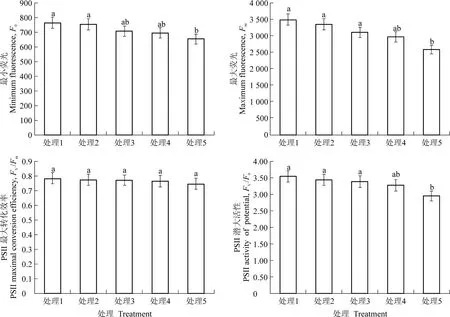
圖7 不同葉面積指數處理下黃果柑葉片葉綠素熒光參數Figure 7 Chlorophyll fluorescence parameters of Huangguogan leaves with different leaf area index
由表3可知,不同LAI 的黃果柑果實可溶性固形物(TSS)、總糖含量和糖酸比隨LAI 降低呈上升趨勢,處理1 和處理2 的黃果柑果實TSS、總糖含量和糖酸比均較高,其中處理1 的這3 個指標分別較處理5 高22.67%、16.89%和36.05%。不同處理間VC 含量差異不顯著。可滴定酸含量(TA)隨 LAI 降低而顯著降低,處理1 的TA 最低,僅0.73 g/100 mL,較處理5 低16.44%。
綜合分析不同LAI 對黃果柑果實商品屬性和營養價值的影響可知,處理1(LAI=2.0)的黃果柑果實最大,單果重最高,縱橫徑最大,果形指數為0.97,果實TSS、總糖含量和糖酸比最高,酸含量最低,整體果實品質最佳。
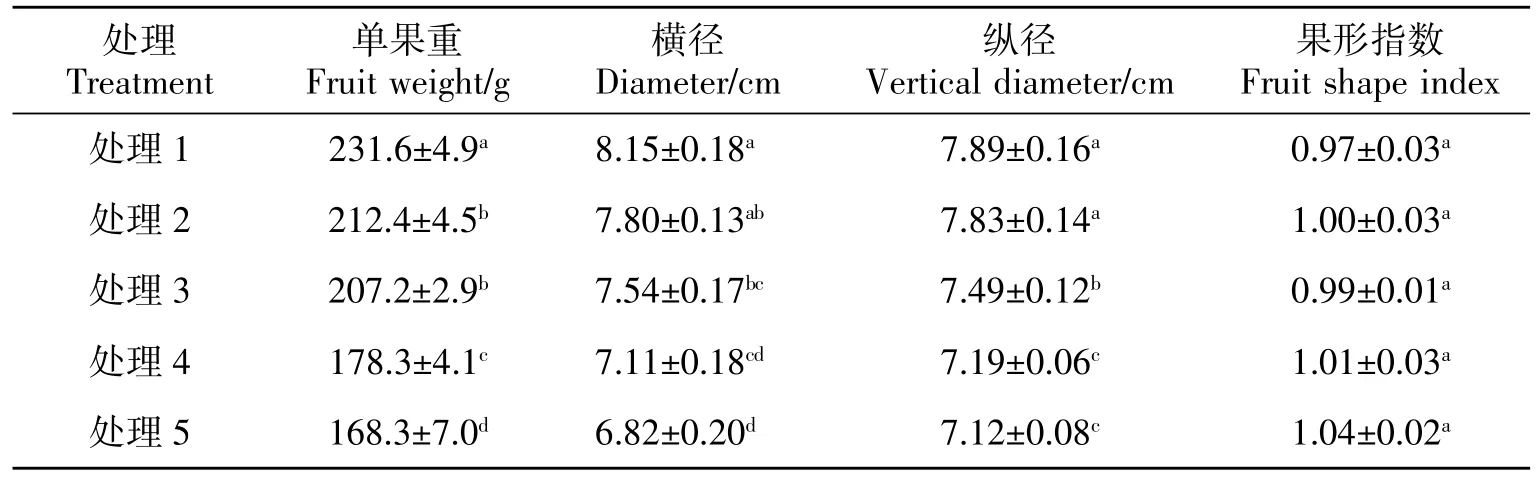
表2 不同葉面積指數處理下黃果柑果實商品質量Table 2 Fruit external quality of Huangguogan with different leaf area index
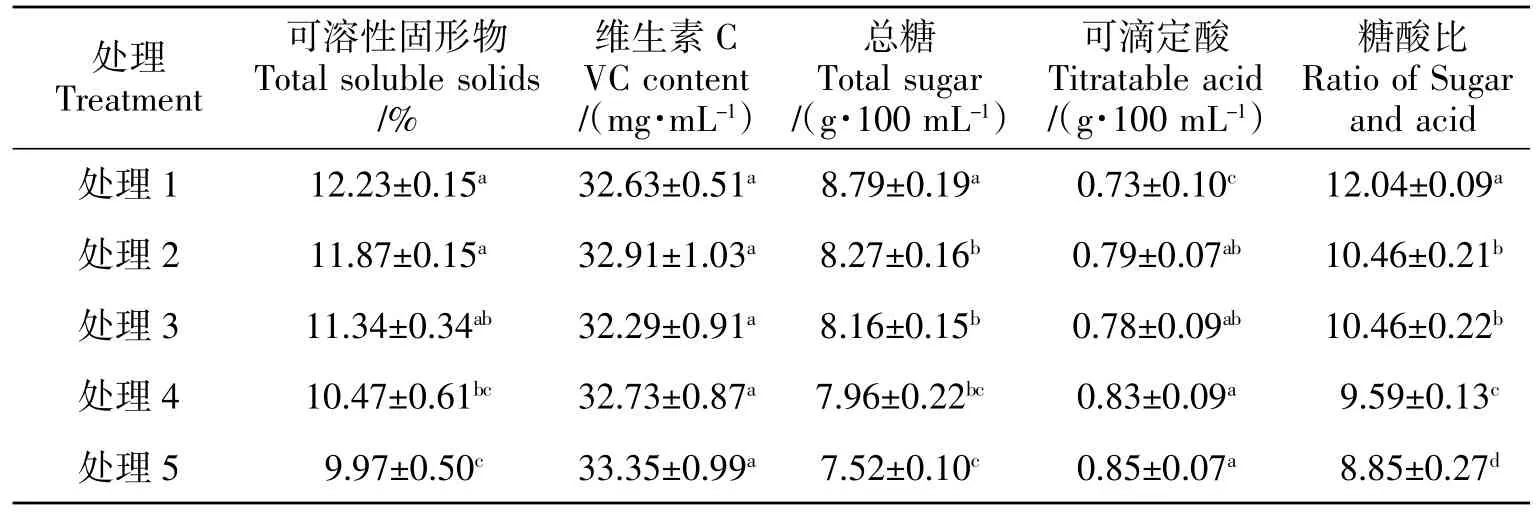
表3 不同葉面積指數處理下黃果柑果實營養價值Table 3 Fruit inner quality of Huangguogan with different leaf area index
3 討論與結論
光合氣體交換參數是對光合作用強弱的直觀體現。本研究結果表明,隨著LAI 降低,黃果柑在4個物候期的 Pn、Gs和 Tr均有不同程度的升高,Ci顯著降低,說明LAI 降低后光照增強、溫度升高使蒸騰作用增強,同時也促進氣孔的開張,使Gs升高,以促進光合作用,使Pn升高。在溫度較高光照較強的幼果期和成熟期出現了較明顯的光抑制現象。光抑制一般與PS II 有關,在出現光抑制時,會積累大量活性氧攻擊光合膜,如果PS II 較強會引起D1 蛋白的降解,以PSII 的可逆與失活應對光抑制,如果沒有PS II 的調節,則可能出現光破壞現象[25]。黃果柑的光抑制表現為可恢復性光抑制,在下午光強減弱的情況下能及時恢復,這說明低LAI 處理下黃果柑有較強PS II 活性。
不同LAI 處理的 PRA-Pn和 CO2-Pn響應曲線變化規律相似,低LAI 處理的LSPn 和CSPn 顯著升高。植物出現光飽和是因為在強光下碳同化速率跟不上光反應速度,而限制Pn的增加[25]。此時,低LAI黃果柑的LSPn 大于高LAI,是因為其在長期的光照良好的環境中,已表現出對強光照的適應性,進而提高了LSPn。而植物出現CO2飽和點是因為RuBP的再生速率受到限制[25],此時低LAI 處理的CSPn 的升高不僅體現了碳同化關鍵酶活性升高,同時也表現出光反應活性的增強,反映出低LAI 條件下黃果柑光合電子傳遞和光合磷酸化活性速率加快。
葉綠素熒光是植物體內發出的天然探針,可快速、靈敏和非破壞性地分析環境因子對光合作用的影響[24]。F0大小與葉綠素含量和PSⅡ反應中心活性相關[26],本研究發現,隨著LAI 的降低黃果柑葉片F0呈上升趨勢,且Fm和Fv/F0也顯著上升,說明低LAI條件下黃果柑PS II 活性增強。
LAI 能直接影響植物群體對光能的截獲與利用,進而影響植株的生長發育[4],最終導致果實品質的不同。本研究結果顯示,隨著LAI 的降低,黃果柑果實單果重、橫徑、縱徑、可溶性固形物、總糖和糖酸比顯著上升,可滴定酸含量顯著下降。可能因為LAI 降低時,冠層內葉片間遮擋程度降低,葉片光合能力增強,干物質積累增多,使果實發育質量整體上升。
綜上所述,LAI 的降低有利于黃果柑光合作用和果實品質的提高,LAI=2.0 條件下,黃果柑光合能力和光合潛力較好,果實品質最佳,是黃果柑適宜的葉面積指數。
猜你喜歡
英語文摘(2022年4期)2022-06-05 07:45:18
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
知識經濟·中國直銷(2016年3期)2016-02-27 16:15:49
現代檢驗醫學雜志(2014年6期)2014-02-02 03:02:04
