不同寄主植物對黏蟲體內能源物質的影響
2019-07-23 08:40:58韓海斌譚瑤李艷艷高書晶徐林波劉愛萍王丹陽
植物保護 2019年2期
韓海斌 譚瑤 李艷艷 高書晶 徐林波 劉愛萍 王丹陽
摘要黏蟲Mythimna separata Walker是一種世界性害蟲。為了明確不同寄主植物對黏蟲幼蟲體內能源物質的影響,選取了玉米、小麥、燕麥、老芒麥4種黏蟲的禾本科寄主植物對黏蟲進行飼喂,并測定黏蟲3、4、5齡期幼蟲體內的可溶性蛋白、可溶性糖及甘油酯的含量;測定了4種寄主植物粗蛋白及粗纖維的含量,并與黏蟲能源物質含量進行了相關性分析。結果表明,喂食燕麥的黏蟲體內可溶性蛋白含量達到263.84ug/mg,顯著高于其他3種植物(P<0.05);喂食4種植物黏蟲體內甘油酯含量為106.69~152.23ug/mg,沒有顯著差異(P>0.05);喂食玉米的黏蟲體內可溶性糖含量為68.64ug/mg,顯著低于其他3種植物(P<0.05)。寄主植物粗蛋白含量與黏蟲體內能源物質含量均呈正相關關系,寄主植物粗纖維含量與黏蟲體內能源物質含量均呈負相關關系。說明不同寄主對黏蟲幼蟲體內能源物質有顯著影響,且與飼喂的寄主植物所含物質含量具有顯著的相關性。
關鍵詞黏蟲;寄主植物;能源物質;相關分析
中圖分類號:S433.4文獻標識碼:A DOI:10.16688/j.zwbh.2018165
黏蟲Mythimna separata(Walker)是鱗翅目Lepidoptera夜蛾科Noctuidae的一種世界性害蟲,具有遷飛性、突發性、暴發性和毀滅性的為害特點,在我國及亞洲其他國家為害較為嚴重。黏蟲幼蟲常成群遷徙,啃食作物葉片,對遷入地作物造成危害。由于其危害的嚴重性,所以在我國很早就對其生物學特性、遷飛規律、飛行生物學等進行了初步研究。20世紀90年代以來,圍繞黏蟲的暴發及遷飛,學者們對其內在調控機制進行了探究,明確了幼蟲密度較高、幼蟲期饑餓、短光照和高溫等因素是誘導成蟲遷飛行為發生的主要原因。Jiang等發現在黏蟲羽化后24h的關鍵時期內,還存在遷飛型向居留型轉化的不可逆的第2次調控機制,從而系統揭示了黏蟲遷飛為害的內在原因與適應機制。同時,由于黏蟲繁殖能力強,較易人工飼養,所以常作為寄生及捕食性天敵昆蟲擴繁的替代寄主,用以人工大規模擴繁天敵昆蟲,對靶標害蟲進行防治。所以為了更好地人工飼養黏蟲,本文對黏蟲能源物質指標與寄主植物的關系進行了研究。
昆蟲生命活動需要的能量大部分來自貯存在體內的能源物質中的化學能。昆蟲體內主要的能源物質包括可溶性蛋白質、可溶性糖、脂肪等。在正常情況下,昆蟲消耗的主要能源物質是糖類,而脂肪和氨基酸僅在昆蟲長期飛行或饑餓等特殊條件下,才能被進一步消耗。脂肪酸是昆蟲體內的主要儲備能源,是至關重要的營養物質,還是昆蟲各種細胞和亞細胞膜結構的重要組成部分。吳孔明和郭予元對棉鈴蟲飛翔的能源物質及消耗進行了研究,發現初羽化個體的能源儲備主要為甘油酯;甘油酯和糖原是棉鈴蟲飛翔的能源物質,蛋白質含量和飛行活動無明顯的相關關系。王夢圓等對傘裙追寄蠅飛行動態能耗進行了研究,得出傘裙追寄蠅飛行過程先消耗糖原,之后再利用甘油酯作為其能源提供。關于黏蟲的能源物質也有一些研究,李克斌和羅禮智研究了不同幼蟲密度對黏蟲成蟲能源物質的影響,得出幼蟲密度不僅影響到初羽化成蟲能源物質的含量,而且也可能影響到成蟲能源物質特別是甘油酯的代謝。蔡彬等分析了溫、濕度對黏蟲成蟲飛行能源物質利用的影響,發現溫、濕度對成蟲飛行能源物質消耗有顯著影響。昆蟲的行為與體內能源物質息息相關,對于植食性昆蟲來說,寄主植物是其獲取能源物質的直接來源。
昆蟲從植物獲得食料是最原始的生態關系,植物所含的營養成分是昆蟲生長發育和繁殖的必要條件。本試驗主要是探討不同齡期黏蟲體內能源物質含量,以及食物中蛋白質與纖維素的含量與黏蟲體內能源物質的關系,從而為深入開展黏蟲的營養生態學研究奠定基礎,為寄主昆蟲及天敵昆蟲大規模擴繁提供依據。
1材料與方法
1.1試驗材料
1.1.1供試昆蟲
黏蟲幼蟲均選用‘遼單青貯625玉米葉在實驗室內飼喂擴繁,選用連續培養的第9代幼蟲進行本試驗。采用不同寄主植物對黏蟲幼蟲進行飼喂,將孵化出的1齡幼蟲移至鋪有濾紙的幼蟲飼養盒中,每盒30頭,每日兩次往飼養盒中放置寄主植物并取出殘渣,幼蟲2齡以后每日清理盒中糞便。分別對3、4、5齡期的幼蟲能源物質進行提取。
1.1.2供試植物
選用無農藥包衣的青貯玉米Zea mays L.(‘遼單青貯625)、小麥Triticum aestivum L(‘內麥5號)、燕麥Arena sativa L(‘貝勒)、老芒麥Ely-mus sibiricus Linn.(野生品種)4種禾本科植物在室內育苗架上進行種植(溫度25℃±1℃,濕度為30%~40%),并飼喂不同齡期黏蟲。
1.2試驗方法
1.2.1黏蟲可溶性蛋白質含量的測定
參照Bradford的考馬斯亮藍G-250法測定蛋白質的含量。用牛血清蛋白制作標準曲線。分別取3齡幼蟲8頭,加入冰冷磷酸緩沖液2mL,4齡4頭、加入冰冷磷酸緩沖液4mL,5齡3頭,加入冰冷磷酸緩沖液3mL,于冰水浴中勻漿。勻漿液于高速離心機10000g,4℃離心10min,取上清液作提取液,加1%考馬斯亮藍試劑5mL,混合均勻,25℃水浴5min,室內平衡后與595nm測定OD值。每個處理3次重復。根據制作的標準曲線計算出蛋白質含量。
1.2.2黏蟲可溶性糖含量的測定
采用蒽酮比色法,用葡萄糖制作標準曲線。取3齡幼蟲8頭,加去離子水1mL;4齡幼蟲4頭,加去離子水1mL;5齡幼蟲3頭,加去離子水3mL。于冰水浴中勻漿。勻漿液于冷凍高速離心機上,15000r/min,25℃離心10min,取上清液作提取液。根據標準曲線計算糖含量。
1.2.3黏蟲幼蟲甘油酯含量測定
參照Bligh & Dyer的乙酰丙酮法測定甘油酯含量,以三油酸甘油酯標準液作標準曲線。按3齡幼蟲8頭,加去離子水1mL;4齡幼蟲4頭,加去離子水1mL;5齡幼蟲3頭,加去離子水3mL,勻漿。勻漿液于高速冷凍離心機上,15000r/min,25℃離心10min,取上清液作提取液。取0.05mL樣品加入3mL提取劑,充分混勻,加0.5mL的0.04mol/L硫酸溶液,靜置分層,吸取0.5mL上清液于另一試管中,加入1.5mL皂化劑,搖勻,65℃水浴保溫5min,再加入1mL氧化劑、1mL乙酰丙酮試劑,搖勻后再次置于65℃水浴10min。以事先配制好的標準液為對照,于紫外分光光度計中420nm處比色,測定OD值,計算甘油酯含量。
1.2.4寄主植物蛋白及粗纖維提取
委托內蒙古農牧漁業生物實驗研究中心對4種黏蟲寄主植物的粗蛋白及粗纖維進行測定,測定方法參照國家標準(GB/T6432-1994、GB/T、6434-20)。
1.2.5數據統計分析
對測定所得數據采用DPS軟件進行統計分析,經方差分析測定其顯著性并進行Duncan氏多重比較。
2結果與分析
2.1取食不同寄主植物黏蟲幼蟲體內蛋白含量
取食不同寄主植物的不同齡期黏蟲幼蟲體內蛋白含量測定結果(表1)表明,取食燕麥的3、4、5齡幼蟲體內蛋白含量均高于取食其他寄主植物,含量分別為224.87、303.95、262.70ug/mg,且差異顯著(P<0.05)。3齡黏蟲取食老芒麥時體內蛋白含量最低,為129.27tug/mg;4齡及5齡黏蟲取食玉米時體內的蛋白含量最低,分別為163.73ug/mg和188.80ug/mg。說明喂食燕麥能夠顯著增加黏蟲幼蟲體內蛋白含量。
2.2取食不同寄主植物黏蟲幼蟲體內甘油酯含量
取食不同寄主植物的不同齡期黏蟲幼蟲體內甘油酯含量測定結果(表1)表明,3齡幼蟲取食老芒麥時體內甘油酯含量最高,為212.16ug/mg,飼喂玉米時體內甘油酯含量最低,為78.33ug/mg;4齡黏蟲取食小麥時體內甘油酯含量最高,為175。34ug/mg,與取食其他植物有顯著差異(P<0.05),取食老芒麥時黏蟲體內甘油酯含量最低,為90.05ug/mg;5齡黏蟲取食老芒麥時體內甘油酯含量最高,為154.48ug/mg,取食燕麥時黏蟲體內甘油酯含量最低,為143.17ug/mg。取食不同植物對黏蟲體內甘油酯含量沒有顯著影響(P>0.05)。
2.3取食不同寄主植物黏蟲幼蟲體內可溶性糖含量
取食不同寄主植物的不同齡期黏蟲幼蟲體內可溶性糖含量測定結果(表1)表明,3齡黏蟲幼蟲取食老芒麥時體內可溶性糖含量最高,為268.11ug/mg,顯著高于取食其他植物(P<0.05),取食玉米時體內可溶性糖含量最低,為71.94ug/mg,與取食其他植物差異顯著(P<0.05);4齡黏蟲幼蟲取食小麥時體內可溶性糖含量最高,為136.66ug/nag,取食玉米時體內可溶性糖含量最低,為46.55ug/mg;5齡黏蟲幼蟲取食老芒麥時體內可溶性糖含量最高,為298.96ug/mg,取食玉米時體內可溶性糖含量最低,為87.43ug/mg。取食玉米的黏蟲幼蟲體內可溶性糖含量均值為68.64ug/mg,顯著低于喂食其他植物時幼蟲體內的可溶性糖含量(P<0.05)。
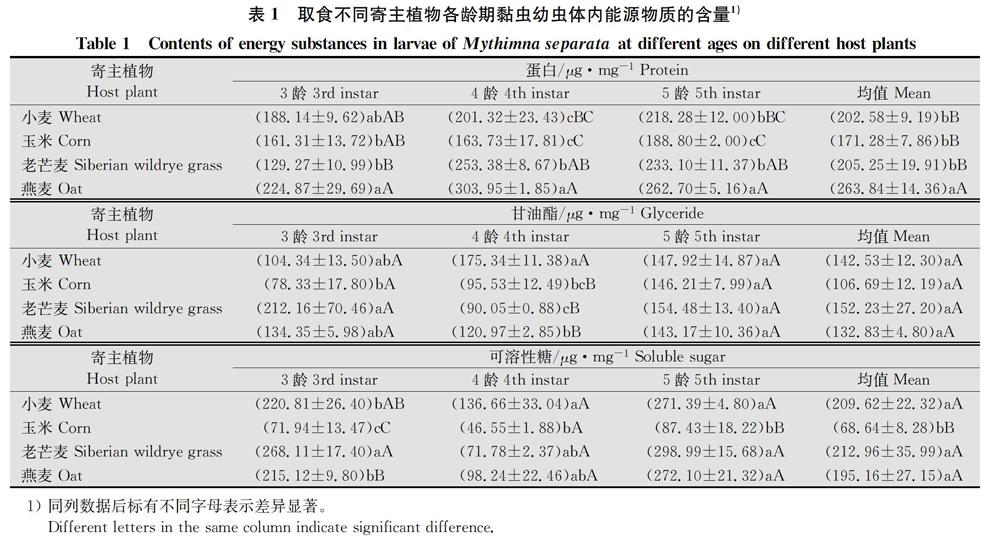
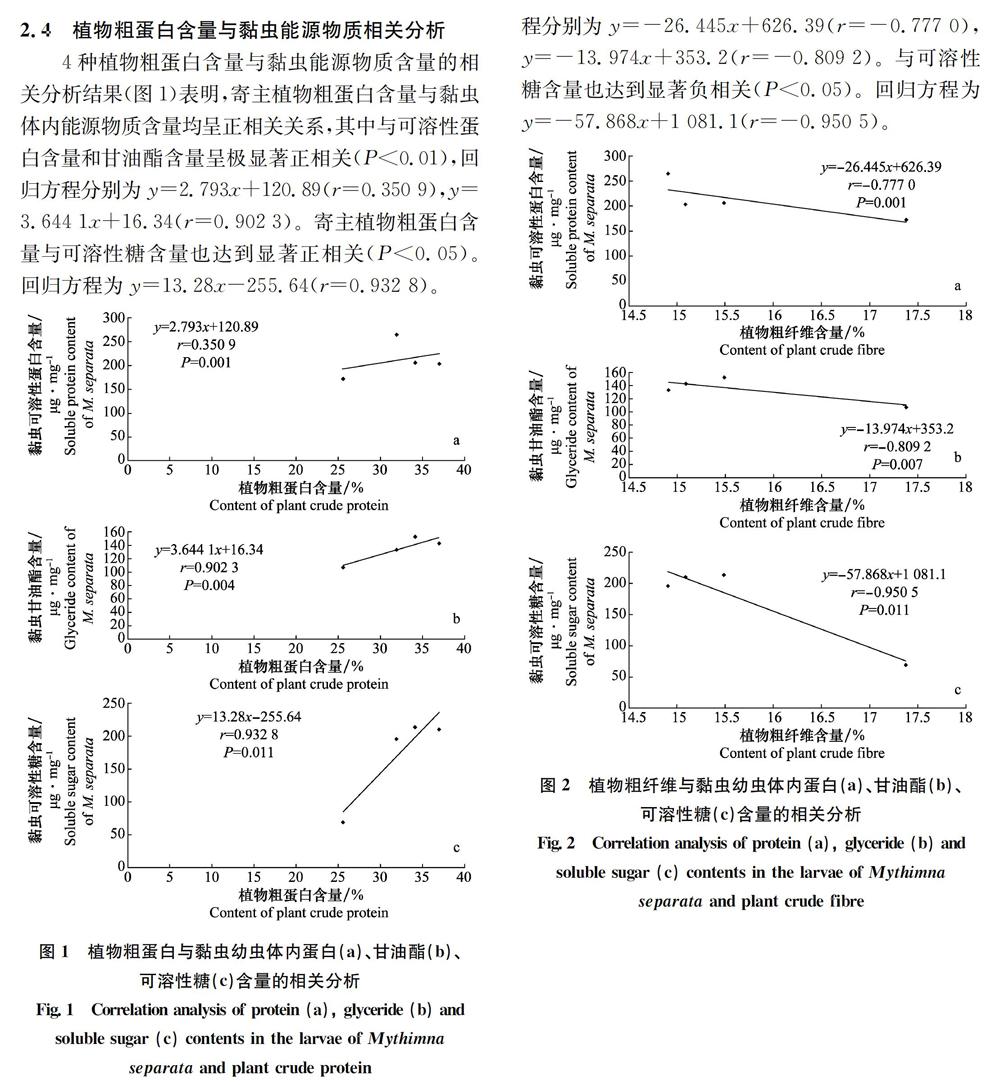
2.5植物粗纖維含量與黏蟲能源物質相關分析
4種植物粗纖維含量與黏蟲能源物質的相關分析結果(圖2)表明,寄主植物粗纖維含量與黏蟲體內能源物質含量均呈負相關關系,其中與可溶性蛋白含量和甘油酯含量呈極顯著負相關(P<0.01),回歸方
3討論
昆蟲從植物獲得食料是最原始的生態關系,植物所含的營養成分是昆蟲生長發育和繁殖的必要條件,同時植物為昆蟲提供生境,影響昆蟲對食料的選擇,還對昆蟲有生態保護作用。植物能為昆蟲提供充分的營養成分。昆蟲通過復雜的代謝作用,不僅把營養物質轉變成自身身體的構成成分,而且還要儲存能源物質,以便提供昆蟲在繁殖、越冬及其遠距離遷飛等生命過程中所需的能量。本試驗對不同寄主植物對黏蟲幼蟲體內能源物質的影響進行了分析,發現飼喂寄主植物對黏蟲體內蛋白含量的影響為燕麥>老芒麥>小麥>玉米,喂食燕麥的黏蟲體內蛋白含量達到263.84ug/mg,顯著高于喂食其他3種植物(P<0.05);飼喂寄主植物對黏蟲體內甘油酯含量的影響為老芒麥>小麥>燕麥>玉米,甘油酯含量在106.69ug/mg到152.23ug/mg之問,喂食4種植物之問沒有顯著差異;飼喂寄主植物對黏蟲體內可溶性糖含量的影響為老芒麥>小麥>燕麥>玉米,喂食玉米的黏蟲體內可溶性糖含量為68.64ug/mg,顯著低于喂食其他3種植物(P<0.05)。結果得出不同寄主植物對黏蟲幼蟲體內能源物質有顯著影響,這與曹玲對飼喂不同寄主植物甜菜夜蛾體內能源物質變化的研究結果一致。
目前對黏蟲進行大規模擴繁都是采用人工飼料,但多代擴繁以后容易出現世代退化嚴重、體內能源物質變化明顯、發育不一致等問題,如果想要得到世代性狀相近,發育一致度較高的黏蟲進行精確試驗的時候,還是需要使用寄主植物直接對黏蟲幼蟲進行飼喂擴繁的方式。而目前使用寄主植物對黏蟲進行飼養都是使用玉米植株作為黏蟲產卵、幼蟲飼養的場所及飼料。本試驗結果得出飼喂玉米的黏蟲幼蟲體內所含的能源物質含量是最低的,可溶性蛋白為171.28ug/mg、甘油酯含量為106.69ug/mg、可溶性糖含量為68.64ug/mg。所以玉米不是最佳的黏蟲擴繁植物;在黏蟲幼蟲4齡時,只有小麥能夠提供給其充足的甘油酯和可溶性糖。根據本試驗結果,我們提出一種較為科學的黏蟲幼蟲飼養方法設想,在黏蟲1~3齡時使用燕麥加老芒麥進行飼喂;到4齡時使用燕麥加小麥飼喂;在5齡以后再用燕麥加老芒麥飼喂。使黏蟲在整個幼蟲發育期都有充分的能源物質供其正常活動、生長及發育。
昆蟲的寄主植物由于營養物質構成不同對昆蟲的生長發育及繁殖具有重要的影響,本試驗對黏蟲4種寄主植物粗蛋白及纖維素進行了測定,并與黏蟲體內能源物質進行了相關性分析,得出寄主植物粗蛋白含量與黏蟲體內能源物質含量均呈正相關關系,其中與可溶性蛋白含量和甘油酯含量呈極顯著正相關(P<0.01),結果與曹玲利用3種不同寄主植物對甜菜夜蛾能源物質的影響的研究一致;寄主植物粗纖維含量與黏蟲體內能源物質含量均呈負相關關系,其中與可溶性蛋白含量和甘油酯含量呈極顯著負相關(P<0.01)。說明黏蟲幼蟲體內能源物質含量與飼喂的寄主植物所含不同物質含量具有顯著的相關性。
本試驗在測定寄主植物中主要營養物質粗蛋白和纖維素含量的基礎上,分析不同寄主對黏蟲體內能源物質含量的影響及植物營養物質與黏蟲體內能源物質的關系,提出黏蟲幼蟲精準飼養的方法,以供更好地闡明寄主植物與黏蟲生長發育、運動遷移的關系,為黏蟲的飼養生態學及可持續治理提供依據。
