虎斑烏賊生殖腺多肽的制備工藝優(yōu)化及其體外抗氧化活性
2019-06-26 09:30:42陳敏陳道海
食品與發(fā)酵工業(yè) 2019年11期
陳敏,陳道海
1(華南理工大學(xué) 食品科學(xué)與工程學(xué)院,廣東 廣州,510640)2(嶺南師范學(xué)院 生命科學(xué)與技術(shù)學(xué)院,廣東 湛江,524048) 3(廣東省粵西海鮮資源可持續(xù)利用工程技術(shù)研究中心,廣東 湛江,524048)
虎斑烏賊(SepiapharaonisEhrenberg, 1831),別稱(chēng)花旗、花西、墨姆,隸屬于軟體動(dòng)物門(mén)(Mollusca)、頭足綱(Cephalopoda)、烏賊目(Sepiida)、烏賊科(Sepiidae)、烏賊屬(Sepia),主要分布于印度洋至西太平洋海域,在我國(guó)南海分布廣泛[1-2]。作為重要的經(jīng)濟(jì)頭足類(lèi)物種,虎斑烏賊具有肉質(zhì)鮮美、營(yíng)養(yǎng)豐富、養(yǎng)殖潛力大、抗病力強(qiáng)、生長(zhǎng)迅速等特點(diǎn),受到國(guó)內(nèi)外研究學(xué)者的廣泛關(guān)注[3]。目前,對(duì)虎斑烏賊的研究主要集中在生物學(xué)特性[4-6]、養(yǎng)殖技術(shù)[7-8]及天然活性物質(zhì)的提取和應(yīng)用[9-11]等方面,而對(duì)于虎斑烏賊蛋白質(zhì)來(lái)源的抗氧化活性肽的研究還鮮見(jiàn)報(bào)道。生殖腺是虎斑烏賊副產(chǎn)物的重要組成部分,占體質(zhì)量的4%~7%,具有高蛋白、低脂肪的特點(diǎn),是一種非常理想的蛋白質(zhì)來(lái)源[12],極具開(kāi)發(fā)前景。
目前,國(guó)內(nèi)外關(guān)于抗氧化活性肽的制備主要集中于商業(yè)蛋白酶酶解法,而對(duì)于發(fā)酵法制備抗氧化活性肽的研究相對(duì)較少[13]。與酶解法相比,微生物蛋白酶來(lái)源廣泛,種類(lèi)繁多,且微生物代謝產(chǎn)生的胞外酶不用分離,降低了生產(chǎn)成本[14]。枯草芽孢桿菌(Bacillussubtilis)具有產(chǎn)酶能力強(qiáng)、發(fā)酵周期短、相對(duì)耐高溫、生產(chǎn)難度小等特點(diǎn),在各行各業(yè)中應(yīng)用廣泛[15]。目前,利用枯草芽孢桿菌及其蛋白酶水解蛋白質(zhì)制備抗氧化活性肽的研究,已取得了一定的成效[16-17]。本文以虎斑烏賊生殖腺作為枯草芽孢桿菌(Bacillussubtilis)的唯一發(fā)酵基質(zhì),以發(fā)酵液的蛋白酶活力和非蛋白氮含量為評(píng)價(jià)指標(biāo),研究底物濃度、培養(yǎng)基的初始pH值、發(fā)酵溫度和發(fā)酵時(shí)間對(duì)發(fā)酵過(guò)程的影響。
1 材料與方法
1.1 材料與試劑
虎斑烏賊:購(gòu)于廣東省湛江市霞山水產(chǎn)品批發(fā)市場(chǎng),解剖取生殖腺,經(jīng)凍干、粉碎后,于-18 ℃貯藏備用;枯草芽孢桿菌CICC 20030:購(gòu)于中國(guó)工業(yè)微生物菌種保藏管理中心;其他試劑:國(guó)產(chǎn)分析純。
1.2 儀器與設(shè)備
紫外可見(jiàn)分光光度計(jì)(UV-3000型),上海美譜達(dá)儀器有限公司;冷凍干燥機(jī)(LGJ-18型),北京四環(huán)科學(xué)儀器廠(chǎng);中草藥粉碎機(jī)(FW135型),天津市泰斯特儀器有限公司;高速組織搗碎機(jī)(DS-1型),上海標(biāo)本模型廠(chǎng);恒溫培養(yǎng)振蕩器(HNY-200B型),天津歐諾儀器股份有限公司;生化培養(yǎng)箱(SPL-250型),天津市萊玻特瑞儀器設(shè)備有限公司;超凈工作臺(tái)(CJ-2S型),天津市泰斯特儀器有限公司;立式壓力蒸汽滅菌鍋(YXQ-LS-75SⅡ型),上海博訊實(shí)業(yè)有限公司;多參數(shù)分析儀(DZB-718-B型),上海儀電科學(xué)儀器股份有限公司;索氏提取器(SXT-06型),上海洪紀(jì)儀器設(shè)備有限公司;全自動(dòng)氨基酸分析儀(S-433D型),德國(guó)塞卡姆公司;全自動(dòng)凱氏定氮儀(K9860型),濟(jì)南海能儀器股份有限公司。
1.3 培養(yǎng)基
營(yíng)養(yǎng)瓊脂培養(yǎng)基:牛肉膏3 g/L,蛋白胨10 g/L,NaCl 5 g/L,瓊脂18 g/L,pH 7.0;種子培養(yǎng)基:牛肉膏3 g/L,蛋白胨10 g/L,NaCl 5 g/L,pH 7.0;液體發(fā)酵培養(yǎng)基:以虎斑烏賊生殖腺為底物的培養(yǎng)基。
1.4 方法
1.4.1 菌種的活化與種子液的制備
將枯草芽孢桿菌在營(yíng)養(yǎng)瓊脂培養(yǎng)基中活化,于37 ℃恒溫培養(yǎng)箱中培養(yǎng)24 h后,挑選形態(tài)正常、生長(zhǎng)良好的單菌落接種至種子培養(yǎng)基中,于37 ℃、180 r/min搖床培養(yǎng)24 h,即得枯草芽孢桿菌種子液。
1.4.2 烏賊生殖腺發(fā)酵工藝的優(yōu)化
在固定發(fā)酵總量50 mL、菌種接種量10%(V/V)、搖床轉(zhuǎn)速180 r/min的條件下,以蛋白酶活力和非蛋白氮含量作為評(píng)價(jià)指標(biāo),分別研究發(fā)酵底物質(zhì)量濃度(10、20、30、40、50 mg/mL)、培養(yǎng)基的初始pH值(5、6、7、8、9)、發(fā)酵溫度(30、34、37、40 ℃)和發(fā)酵時(shí)間(12、24、36、48、60 h)對(duì)發(fā)酵過(guò)程的影響。在上述單因素實(shí)驗(yàn)的基礎(chǔ)上,對(duì)發(fā)酵底物質(zhì)量濃度、初始pH值、發(fā)酵溫度和發(fā)酵時(shí)間進(jìn)行L9(34)正交實(shí)驗(yàn),優(yōu)化枯草芽孢桿菌發(fā)酵虎斑烏賊生殖腺的工藝流程。
1.4.3 烏賊生殖腺多肽的制備工藝
烏賊生殖腺原料粉→加水勻漿并調(diào)節(jié)pH值→高溫滅菌(121 ℃,20 min)→接種→發(fā)酵→沸水浴(100 ℃,15 min)→冷卻→離心(6 000 r/min,30 min)→上清液→冷凍干燥→虎斑烏賊生殖腺粗肽樣品(SPGPs)
1.4.4 理化指標(biāo)的檢測(cè)
蛋白質(zhì)含量的測(cè)定參照GB 5009.5—2016《食品中蛋白質(zhì)的測(cè)定》,采用凱氏定氮法;脂肪含量的測(cè)定參照GB 5009.6—2016《食品中脂肪的測(cè)定》,采用索氏提取法;水分含量的測(cè)定參照GB 5009.3—2016《食品中水分的測(cè)定》,采用直接干燥法;灰分的測(cè)定參照GB 5009.4—2016《食品中灰分的測(cè)定》,采用高溫灼燒法。
1.4.5 發(fā)酵液蛋白酶活力的測(cè)定
蛋白酶活力的測(cè)定參照GBT 23527—2009《蛋白酶制劑》中的福林法進(jìn)行測(cè)定。酶活力單位定義為:1 mL酶液在40 ℃、pH值為7.5的條件下,每分鐘水解酪蛋白產(chǎn)生1 μg酪氨酸,即為1個(gè)酶活力單位,以U/mL表示。
酶活力標(biāo)準(zhǔn)曲線(xiàn)的繪制:取質(zhì)量濃度為0、10、20、30、40、50、60 mg/mL的L-酪氨酸標(biāo)準(zhǔn)溶液各1 mL,分別加入5 mL Na2CO3溶液(42.4 g/L)、1 mL福林試劑,混勻后,于40 ℃水浴顯色20 min,以不含酪氨酸的0管為空白,測(cè)定其在680 nm處的吸光值。以吸光度A為縱坐標(biāo),酪氨酸濃度為橫坐標(biāo),繪制標(biāo)準(zhǔn)曲線(xiàn)y=0.010 6x-0.002(R2=0.999 9)。
1.4.6 發(fā)酵液中非蛋白氮含量的測(cè)定
取發(fā)酵后的上清液,加入等體積三氯乙酸(trichloroacetic acid, TCA)溶液(150 g/L),混勻后,室溫靜置30 min,于6 000 r/min離心20 min,用凱氏定氮法測(cè)定上清液中非蛋白(包括多肽和氨基酸)氮的含量。
1.4.7 氨基酸組分分析
氨基酸含量的測(cè)定參照GB 5009.124—2016《食品中氨基酸的測(cè)定》,采用氨基酸分析儀測(cè)定樣品中酸水解氨基酸的組成和含量。
1.4.8 羥自由基清除能力的測(cè)定
參照文獻(xiàn)[18]的方法并作適當(dāng)修改。向試管中依次加入0.5 mL FeSO4溶液(6 mmol/L)、0.5 mL水楊酸-乙醇溶液(6 mmol/L)及0.5 mL樣液,最后加入0.5 mL H2O2溶液(6 mmol/L)啟動(dòng)反應(yīng),并用蒸餾水補(bǔ)至總體積為5 mL。其中,對(duì)照管不加樣液,底管不加H2O2,混勻后于37 ℃水浴中保溫30 min,測(cè)定其在波長(zhǎng)510 nm處的吸光值。以抗壞血酸作為陽(yáng)性對(duì)照,按式(1)計(jì)算清除率:

(1)
式中:A0,對(duì)照管的吸光值;A,樣品管的吸光值;AD,底管的吸光值。
1.4.9 超氧陰離子自由基清除能力的測(cè)定
參照文獻(xiàn)[19]的方法并作適當(dāng)修改。向試管中依次加入1 mL樣液、5 mL Tris-HCl緩沖液(0.1 mol/L, pH 8.2),混勻后,于25 ℃水浴中保溫10 min,取出后立即加入0.3 mL鄰苯三酚溶液(3 mmol/L),迅速搖勻后倒入比色皿,每隔30 s測(cè)定其在波長(zhǎng)320 nm處的吸光值,持續(xù)4 min。以蒸餾水代替樣液作為空白對(duì)照,以抗壞血酸作為陽(yáng)性對(duì)照,計(jì)算線(xiàn)性范圍內(nèi)每分鐘吸光度的增加量。按式(2)計(jì)算清除率:

(2)
式中:A0,空白對(duì)照的吸光值;A,樣品組的吸光值。
1.4.10 亞鐵離子螯合能力的測(cè)定
參照文獻(xiàn)[20]的方法并作適當(dāng)修改。向試管中依次加入1 mL樣液、3.7 mL蒸餾水及0.1 mL FeSO4溶液(2 mmol/L),混勻后室溫靜置5 min,加入0.2 mL菲洛嗪溶液(5 mmol/L),混勻后室溫靜置10 min。以蒸餾水代替樣液作為空白對(duì)照,以乙二胺四乙酸(EDTA)作為陽(yáng)性對(duì)照,測(cè)定其在波長(zhǎng)562 nm處的吸光值。按式(3)計(jì)算螯合率:
(3)
式中:A0,空白對(duì)照的吸光值;A,樣品組的吸光值。
2 結(jié)果與分析
2.1 烏賊生殖腺原料粉的基本成分分析
由表1可知,虎斑烏賊生殖腺原料粉中粗蛋白質(zhì)和粗脂肪含量分別為62.71 g/100g和0.52 g/100g,是一種高蛋白、低脂肪的優(yōu)質(zhì)蛋白質(zhì)來(lái)源。其灰分含量相對(duì)較高,達(dá)6.64 g/100g,這可能與海洋環(huán)境有關(guān)。戴宏杰等[12]的研究也表明,雌性虎斑烏賊纏卵腺的粗蛋白質(zhì)和粗脂肪含量分別為18.40 g/100g和0.16 g/100g(以鮮重計(jì)),且其氨基酸種類(lèi)豐富,組成和比例合理,具有很高的營(yíng)養(yǎng)價(jià)值和開(kāi)發(fā)潛力。
表1 虎斑烏賊生殖腺原料粉的基本成分單位:g/100g

Table 1 Elementary composition in cuttlefish(Sepia pharaonis) gonads power
2.2 烏賊生殖腺發(fā)酵工藝的優(yōu)化
2.2.1 發(fā)酵底物濃度對(duì)蛋白酶活力和非蛋白氮含量的影響
由圖1可知,在一定的范圍內(nèi),隨著底物濃度的增加,發(fā)酵液的蛋白酶活力不斷增加,當(dāng)?shù)孜镔|(zhì)量濃度達(dá)到30 mg/mL時(shí),發(fā)酵液的蛋白酶活力達(dá)到最大值,而后逐漸下降。這可能是因?yàn)榈孜餄舛冗^(guò)低時(shí),培養(yǎng)基營(yíng)養(yǎng)不足,導(dǎo)致菌株的生長(zhǎng)或產(chǎn)酶受到限制;而底物濃度過(guò)高時(shí),培養(yǎng)基的黏度過(guò)高影響通氣,從而導(dǎo)致菌株的蛋白酶活力降低。而底物質(zhì)量濃度在10~50 mg/mL,發(fā)酵液中的非蛋白氮含量與底物濃度始終呈正相關(guān)。結(jié)合成本和節(jié)約等因素,選擇30 mg/mL作為最適的底物濃度。

圖1 底物濃度對(duì)酶活力和非蛋白氮含量的影響Fig.1 Effect of substrate concentration on enzyme activity and non-protein nitrogen contents
2.2.2 初始pH值對(duì)蛋白酶活力和非蛋白氮含量的影響
由圖2可知,隨著培養(yǎng)基初始pH值的升高,發(fā)酵液的蛋白酶活力和非蛋白氮含量均有所增加。當(dāng)初始pH值為6.0時(shí),蛋白酶活力和非蛋白氮含量均達(dá)到最大值,繼續(xù)升高pH值,蛋白酶活力迅速下降,非蛋白氮含量也開(kāi)始緩慢降低。這主要是因?yàn)榕囵B(yǎng)基的初始pH值會(huì)影響菌株的生長(zhǎng)繁殖及相關(guān)代謝產(chǎn)物的生物合成途徑,初始pH值過(guò)高或者過(guò)低,都會(huì)影響蛋白酶的合成和積累,并進(jìn)一步影響非蛋白氮含量的增加。因此,選擇初始pH 6.0作為培養(yǎng)基的最適pH值。

圖2 初始pH值對(duì)酶活力和非蛋白氮含量的影響Fig.2 Effect of initial pH on enzyme activity and non- proteinlnitrogen contents
2.2.3 發(fā)酵溫度對(duì)蛋白酶活力和非蛋白氮含量的影響
發(fā)酵溫度是影響微生物生長(zhǎng)及代謝產(chǎn)物積累的重要因素。由圖3可知,隨著發(fā)酵溫度的升高,發(fā)酵液的蛋白酶活力增加,34 ℃時(shí),蛋白酶活力達(dá)到最大,此后迅速下降。而在30~40 ℃,發(fā)酵液的非蛋白氮含量與發(fā)酵溫度始終呈正相關(guān)。主要原因可能是該菌株的最適產(chǎn)酶溫度(34 ℃)與酶解反應(yīng)的最適溫度不一致。隨著溫度的升高,酶解反應(yīng)的速率逐漸增加,使發(fā)酵液的非蛋白氮含量不斷增加。在本次實(shí)驗(yàn)中,超過(guò)34 ℃后,發(fā)酵液的蛋白酶活力迅速下降,而非蛋白氮含量的增加相對(duì)緩慢。因此,選擇34 ℃作為最適的發(fā)酵溫度。
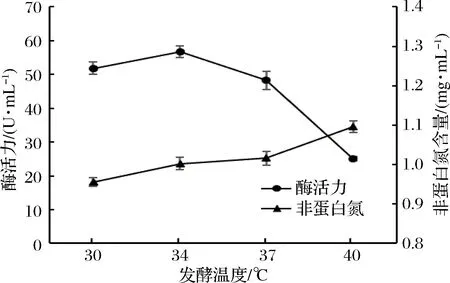
圖3 發(fā)酵溫度對(duì)酶活力和非蛋白氮含量的影響Fig.3 Effect of incubation temperature on enzyme activity and non-protein nitrogen contents
2.2.4 發(fā)酵時(shí)間對(duì)蛋白酶活力和非蛋白氮含量的影響
由圖4可知,隨著發(fā)酵時(shí)間的增加,發(fā)酵液的蛋白酶活力和非蛋白氮含量均有所增加,在發(fā)酵時(shí)間達(dá)到48 h時(shí),發(fā)酵液的蛋白酶活力達(dá)到最大值,之后又迅速下降。發(fā)酵液的非蛋白氮含量在60 h時(shí)達(dá)到最大,此后緩慢下降。原因可能是隨著發(fā)酵時(shí)間的增加,有害產(chǎn)物的不斷積累,菌種逐步進(jìn)入衰退期,使發(fā)酵液的酶活力降低,并進(jìn)一步導(dǎo)致發(fā)酵液的非蛋白氮含量降低。考慮到發(fā)酵效率等因素,選擇48 h作為最適的發(fā)酵時(shí)間。
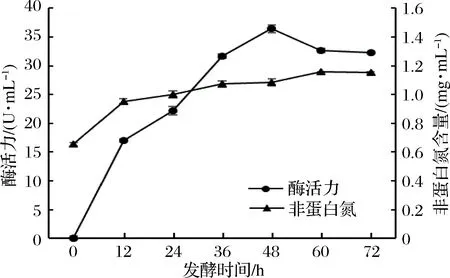
圖4 發(fā)酵時(shí)間對(duì)酶活力和非蛋白氮含量的影響Fig.4 Effect of fermentation time on enzyme activity and non-protein nitrogen contents
2.2.5 正交試驗(yàn)
在上述單因素實(shí)驗(yàn)的基礎(chǔ)上,以蛋白酶活力和非蛋白氮含量為評(píng)價(jià)指標(biāo),選取發(fā)酵底物濃度(A)、培養(yǎng)基的初始pH值(B)、發(fā)酵溫度(C)和發(fā)酵時(shí)間(D)進(jìn)行L9(34)正交試驗(yàn),結(jié)果見(jiàn)表2。

表2 正交試驗(yàn)結(jié)果Table 2 Results of orthogonal experiment
由表2可知,發(fā)酵過(guò)程中,各因素對(duì)蛋白酶活力影響的主次順序?yàn)镈>C>B>A,最優(yōu)組合為A2B2C2D3。各因素對(duì)非蛋白氮含量影響的主次順序?yàn)锳>D>C>B,最優(yōu)組合為A3B2C3D3。由于上述2個(gè)指標(biāo)獨(dú)立分析后得出的最佳發(fā)酵條件不同,因此,需要對(duì)2個(gè)指標(biāo)進(jìn)行綜合分析以確定最終的發(fā)酵工藝。在本次實(shí)驗(yàn)中,蛋白酶活力作為主要的評(píng)價(jià)指標(biāo),能夠更加直觀而準(zhǔn)確地反映菌種的生長(zhǎng)及產(chǎn)酶情況,并進(jìn)一步影響多肽的產(chǎn)量。而非蛋白氮含量側(cè)面反映了發(fā)酵過(guò)程中大分子蛋白質(zhì)降解為小分子肽和氨基酸的動(dòng)態(tài)變化,可作為輔助性指標(biāo)監(jiān)視蛋白質(zhì)的水解程度。底物濃度對(duì)非蛋白氮含量影響較大,對(duì)酶活力影響較小,但考慮到發(fā)酵成本以及底物濃度過(guò)高時(shí),底物粘壁嚴(yán)重等問(wèn)題,選擇A2作為最適的底物濃度水平。發(fā)酵溫度對(duì)蛋白酶活力的影響大于非蛋白氮含量,因此,選擇C2作為最適的發(fā)酵溫度。初始pH值和發(fā)酵時(shí)間均為B2和D3最佳。
據(jù)中正輿情中心博客提供的數(shù)據(jù)顯示,該事件輿論發(fā)酵的路徑經(jīng)歷了從自媒體介質(zhì)到傳統(tǒng)媒體平臺(tái)的過(guò)程。 事件起始,微信平臺(tái)開(kāi)始流傳事件消息,內(nèi)容為死者母親見(jiàn)到尸體哭泣、當(dāng)日下午拍攝的訴說(shuō)“孩子在學(xué)校沒(méi)有人身安全”的視頻片段與截圖、T中學(xué)群眾聚集的照片等。 這些內(nèi)容隨后迅速擴(kuò)散到網(wǎng)絡(luò)空間,傳播內(nèi)容以趙某“被校霸打死的‘內(nèi)幕’”為主,甚至出現(xiàn)了多種針對(duì)政府部門(mén)包庇兇手的流言。 自媒體的極速傳播迅速帶動(dòng)了傳統(tǒng)平面媒體的介入,包括人民網(wǎng)、中國(guó)新聞網(wǎng)、中國(guó)青年網(wǎng)、中國(guó)經(jīng)濟(jì)網(wǎng)、中國(guó)廣播網(wǎng)、澎湃新聞、騰訊、網(wǎng)易、搜狐、新浪等眾多新聞網(wǎng)站都對(duì)該事件進(jìn)行了報(bào)道。[4]
根據(jù)正交試驗(yàn)結(jié)果進(jìn)行綜合分析,選擇A2B2C2D3作為發(fā)酵的最佳組合條件,即發(fā)酵底物濃度30 mg/mL,培養(yǎng)基的初始pH值6.0,發(fā)酵溫度34 ℃,發(fā)酵時(shí)間60 h。在此條件下,發(fā)酵液的蛋白酶活力為(66.66±0.64) U/mL,非蛋白氮含量為(1.10±0.02) mg/mL。
2.3 烏賊生殖腺發(fā)酵物的抗氧化活性
2.3.1 羥自由基清除能力
羥自由基(·OH)是體內(nèi)最活潑、進(jìn)攻性最強(qiáng)的活性氧自由基,能夠殺死紅細(xì)胞,降解DNA、細(xì)胞膜和多糖化合物,具有很高的危害性[24]。
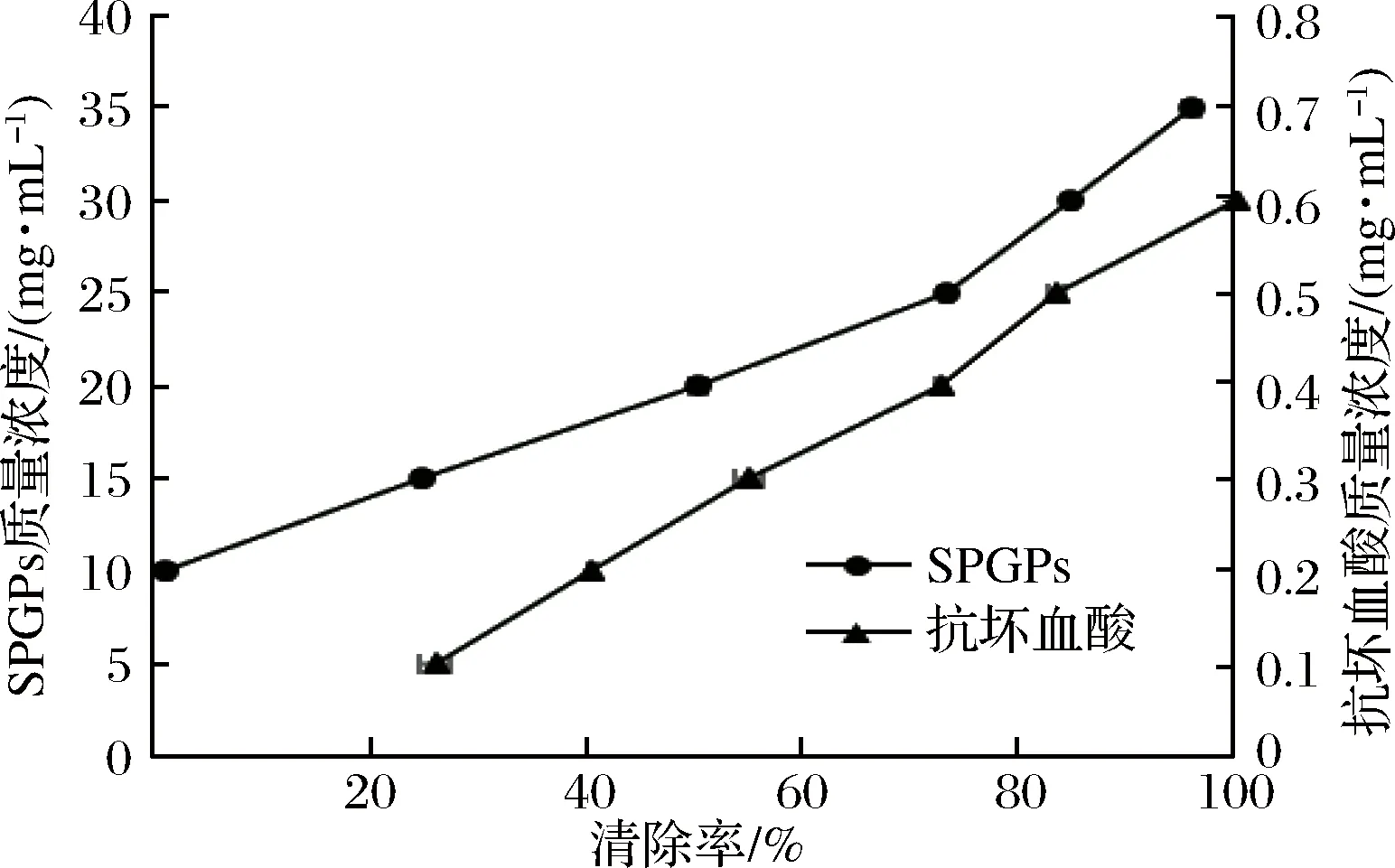
圖5 SPGPs和抗壞血酸的羥自由基清除能力Fig.5 Hydroxyl free radical scavenging abilities of SPGPs and ascorbic acid
由圖5可知,抗壞血酸具有很強(qiáng)的·OH清除能力,SPGPs的清除能力相對(duì)較弱。隨著SPGPs濃度的增加,其·OH清除能力逐漸增加,當(dāng)SPGPs的濃度達(dá)到35 mg/mL時(shí),其·OH清除率可達(dá)92.02%。對(duì)濃度和清除率之間作回歸方程并計(jì)算半抑制濃度(IC50),得到SPGPs清除·OH的IC50值為19.40 mg/mL,高于金槍魚(yú)多肽(IC50=28.81 mg/mL)[25]。總的來(lái)說(shuō),SPGPs具有一定的·OH清除能力。
2.3.2 超氧陰離子自由基清除能力
超氧陰離子自由基是人體內(nèi)產(chǎn)生的活性氧自由基,過(guò)高會(huì)導(dǎo)致機(jī)體損傷和疾病發(fā)生[23]。由圖6可知,與抗壞血酸相比,SPGPs的清除能力相對(duì)較弱,其清除率隨SPGPs濃度的增加而增加。當(dāng)SPGPs濃度為15 mg/mL時(shí),其清除率為61.10%,隨后清除率變化減慢。對(duì)濃度和清除率之間作回歸方程,計(jì)算得到SPGPs清除的IC50值為8.90 mg/mL。楊濤等[24]用堿性蛋白酶水解海參內(nèi)臟制備了海參多肽,其清除的IC50值為21.6 mg/mL。胡小軍等[25]用復(fù)合蛋白酶酶解魷魚(yú)蛋白制備抗氧化肽,其酶解產(chǎn)物清除的IC50值為10.3 mg/mL,均低于SPGPs。綜上分析表明,SPGPs具有良好的清除能力。
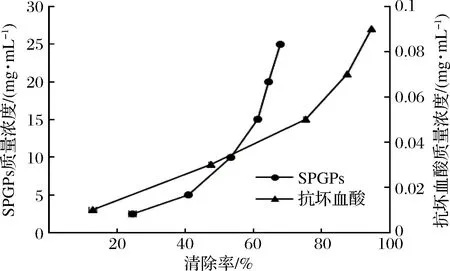
圖6 SPGPs和抗壞血酸的超氧陰離子自由基清除能力Fig.6 Superoxide anion radical scavenging abilities of SPGPs and ascorbic acid
2.3.3 亞鐵離子螯合能力
過(guò)渡金屬離子,如Fe2+和Cu2+,能夠參與體內(nèi)的許多氧化反應(yīng),催化·OH等自由基的產(chǎn)生并介導(dǎo)脂質(zhì)過(guò)氧化[26]。由圖7可知,SPGPs具有一定的Fe2+螯合能力,且螯合能力隨SPGPs濃度的增加而增加,具有很好的量效關(guān)系。

圖7 SPGPs和EDTA的亞鐵離子螯合能力Fig.7 Ferrous ion chelating power of SPGPs and EDTA
當(dāng)SPGPs質(zhì)量濃度達(dá)到2.5 mg/mL時(shí),其Fe2+螯合率達(dá)到78.00%。對(duì)濃度和螯合率之間作回歸方程,計(jì)算得到SPGPs螯合Fe2+的IC50值為1.75 mg/mL。但與陽(yáng)性對(duì)照EDTA相比,SPGPs螯合Fe2+的能力相對(duì)較弱,這可能與SPGPs的純度有關(guān)。ZHOU等[27]利用木瓜蛋白酶水解光棘球海膽(Strongylocentrotusnudus)生殖腺,酶解產(chǎn)物經(jīng)10 kDa超濾膜過(guò)濾后,其螯合Fe2+的IC50值為7.26 mg/mL。鄭杰等[28]用木瓜蛋白酶酶解海蜇生殖腺,得到酶解產(chǎn)物Fe2+螯合能力的IC50值為1.73 mg/mL,這與本次實(shí)驗(yàn)結(jié)果相近。綜上所述,SPGPs具有良好的Fe2+螯合能力。
2.4 SPGPs的氨基酸組成分析
由表3可知,SPGPs的氨基酸組成中Lys的含量最高,占總氨基酸含量的11.1%,其次分別是Asp(10.7%)、His(9.3%)、Glu(8.8%)、Thr(8.7%)和Phe(7.3%)。其必需氨基酸(essential amino acid,EAA)含量豐富,組成合理,占總氨基酸(total amino acid,TAA)含量的42.1%,具有較高的營(yíng)養(yǎng)價(jià)值。

表3 SPGPs的氨基酸組成分析Table 3 Amino acid composition of SPGPs
注:★表示必需氨基酸;抗氧化氨基酸(antioxidant amino acid,AAA)包括:Asp、Glu、Pro、Met、Tyr、Phe、His、Lys;疏水性氨基酸(Hydrophobic amino acids,HAA)包括:Pro、Ala、Val、Met、Ile、Leu、Tyr、Phe。
相關(guān)研究表明,多肽的抗氧化活性與氨基酸的組成和序列密切相關(guān),疏水性氨基酸及His、Lys等氨基酸含量較高的多肽往往具有更高的抗氧化活性[29]。在SPGPs中,疏水性氨基酸的含量為107.2 mg/g,占總氨基酸含量的39.1%。其抗氧化氨基酸的含量為173.4 mg/mL,占總氨基酸含量的63.3%,這也側(cè)面印證了SPGPs具有較好的抗氧化潛力。另外,研究表明,His和酸性氨基酸(Asp、Glu)是參與金屬螯合作用的重要氨基酸[30]。在SPGPs中Asp、His和Glu的含量都很高,分別為29.3、25.6和24.0 mg/g,這可能是SPGPs具有良好的Fe2+螯合能力的原因。
3 結(jié)論
以虎斑烏賊生殖腺作為枯草芽孢桿菌的唯一發(fā)酵基質(zhì),以發(fā)酵液的蛋白酶活力和非蛋白氮含量為評(píng)價(jià)指標(biāo),研究發(fā)酵的底物濃度、培養(yǎng)基初始pH值、發(fā)酵溫度和發(fā)酵時(shí)間對(duì)發(fā)酵結(jié)果的影響。枯草芽孢桿菌發(fā)酵虎斑烏賊生殖腺的最佳條件為:底物質(zhì)量濃度30 mg/mL,初始pH值6.0,發(fā)酵溫度34 ℃,發(fā)酵時(shí)間60 h。在此條件下,發(fā)酵液的蛋白酶活力及非蛋白氮含量分別為(66.66±0.64) U/mL和(1.10±0.02) mg/mL。SPGPs具有一定的體外抗氧化活性,其羥自由基清除能力、超氧陰離子自由基清除能力及亞鐵離子螯合能力的IC50值分別為19.40、8.90和1.75 mg/mL。氨基酸組成分析表明,SPGPs中Lys含量最高,其抗氧化氨基酸和疏水性氨基酸的含量分別占總氨基酸含量的63.3%和39.1%,具有很高的抗氧化潛力。采用枯草芽孢桿菌發(fā)酵虎斑烏賊生殖腺,制備得到的多肽具有良好的體外抗氧化活性,可以作為天然抗氧化劑或功能性食品添加劑應(yīng)用于食品加工中。
