乙酸脅迫下巴氏醋酸桿菌發酵過程中微環境水平的應答分析
2019-06-26 09:30:32史偉高玲夏小樂
食品與發酵工業 2019年11期
史偉,高玲,夏小樂*
1(工業生物技術教育部重點實驗室(江南大學),江蘇 無錫,214122) 2(江南大學 生物工程學院,江蘇 無錫,214122)
醋酸菌是一種革蘭氏陰性專性好氧菌,屬于α-變形桿菌的Acetobacteraceae[1]。醋酸菌具有“氧化發酵”的特殊能力,在發酵過程中會將乙醇逐漸氧化為乙酸。醋酸發酵是底物和產物抑制發酵進行的典型例子。隨著發酵過程的進行,乙酸逐漸積累,在質量濃度5 g/L下產生較強的細胞毒性[2-3],對菌體細胞施加嚴重的酸脅迫壓力,這極大地降低了醋酸發酵生產效率,因此改善醋酸桿菌的耐酸特性具有重要的意義。

本研究曾通過酸激復合紫外線誘變組合獲得1株名為A.pasteurianusCICIM B7003-02,具有較高乙酸耐受性的突變菌株,在不同脅迫條件下突變株超過了親本株的發酵速率和乙酸產量[16]。巴氏醋酸桿菌是嚴格的好氧菌,利用7.5 L發酵罐保證其溶氧,采用不同的發酵方式,研究突變菌株和親本菌株的發酵特性,以及不同發酵階段酸脅迫下菌體細胞膜脂肪酸、胞內氨基酸含量、胞內ATP含量和乙醇呼吸鏈活性,以更好地了解巴氏醋酸桿菌耐受酸脅迫的機理,解析其適應特定發酵環境的優勢。
1 材料與方法
1.1 材料與試劑
1.1.1 菌株
巴氏醋酸桿菌親本菌株A.pasteurianusCICIM B7003,一種典型的工業用醋酸發酵菌株,分離自釀造工廠,保存于本實驗室,巴氏醋酸桿菌突變菌株A.pasteurianusCICIM B7003-02保存于本實驗室。
1.1.2 培養基與試劑
種子培養基(g/L):葡萄糖10,酵母粉10,121 ℃滅菌20 min,接種前加無水乙醇至24。
發酵培養基(g/L):葡萄糖10,酵母粉10,KH2PO40.6,無水MgSO40.4,121 ℃滅菌20 min,接種前加入過濾除菌的初始乙酸10,無水乙醇至40。
葡萄糖、無水乙醇、冰醋酸、三氯乙酸:分析純;甲醇、正己烷:色譜純,國藥集團;有機酸標準品:TCI公司;輔酶Q9標準品:Sigma公司;ATP測定試劑盒:碧云天生物技術有限公司。
1.2 儀器與設備
臺式高速離心機,德國Sigma公司;超聲細胞破碎機,寧波新芝生物科技股份有限公司;BioFlo115 發酵罐,美國NBS公司;Enspire多標記檢測系統(酶標儀),美國珀金埃爾默有限公司;Agilent1260高效液相色譜儀,美國安捷倫公司;GC-MS TSQ8000三重四極桿氣質聯用儀、LC-MS TSQ quantum Ultra EMR,美國賽默飛世爾科技公司。
1.3 實驗方法
1.3.1 發酵培養方法
搖瓶活化和種子培養參照文獻[17]。
分批發酵:將培養24 h的種子接種到發酵罐,總發酵體積為4 L,初始乙醇質量濃度為40 g/L,初始乙酸為10 g/L,接種量10%,培養溫度30 ℃,通氣速率設定為1 L/min (0.25 vvm),轉速400 r/min,溶氧下降到20%時,調節通氣速率為1.6 L/min (0.4 vvm)。
半連續發酵:(1)發酵啟動階段。3 L新鮮發酵培養基,初始醋酸和乙醇質量濃度分別為10 g/L和30 g/L,接種量10%,培養24 h的種子接種到發酵罐,培養溫度為30 ℃,轉速400 r/min,通氣速率設定為0.865 L/min (0.25 vvm),當發酵罐中殘余乙醇質量濃度降到5 g/L時,泵入260 g/L乙醇的新鮮培養基0.54 L,共計4 L培養液。培養溫度為30 ℃,通氣速率上調至1.2 L/min (0.3 vvm)繼續培養。(2)重復補料發酵階段:當發酵罐中殘余乙醇質量濃度降到5 g/L時,排出總發酵體積(4 L)43% (1.72 L)的發酵液,緩慢補入81.4 g/L乙醇的新鮮培養基1.72 L,培養溫度為30 ℃,通氣速率1.2 L/min(0.3 vvm)。
1.3.2 發酵過程參數的測定
乙醇、乙酸、細胞干重的測定參照文獻[18]。
1.3.3 發酵過程ATP含量的測定
利用ATP檢測試劑盒(ATP Assay Kit)檢測發酵過程菌體內部的ATP,向 96 孔板中加入100 μL ATP檢測工作液,放置3~5 min,加50 μL樣品或者標準品,檢測時間設置10 s,測Lum值,圖1為ATP測定的標準曲線。

圖1 Lum值與ATP濃度的線性關系Fig.1 Linear relationship between Lum value and ATP concentration
1.3.4 ADH和ALDH酶活的測定
采用比色法,具體操作參考文獻[19]。
1.3.5 輔酶Q9的測定
輔酶Q9的提取:取適量發酵液,離心得菌體,加入10 mL丙酮重懸,冰浴條件下超聲波破碎20 min (240 W, 3 s/3 s),破碎完全后,5 000×g、4 ℃下離心15 min;取上清液,在45 ℃的旋轉蒸發儀中濃縮,上清液不再減少時再加入50 mL石油醚萃取;充分混勻,分層后將含有輔酶的萃取液用旋轉蒸發儀45 ℃條件下濃縮至干,最后加入無水乙醇10 mL溶解,放入冰箱冷凍,析出膽固醇等雜質,過濾后用無水乙醇定容到50 mL,待測。
輔酶Q9含量的測定條件:純甲醇為洗脫液,流速設置為0.3 mL/min,柱溫45 ℃,色譜柱為BEH chemistry C18柱,進樣量2 μL,圖2為輔酶Q9含量的標準曲線。
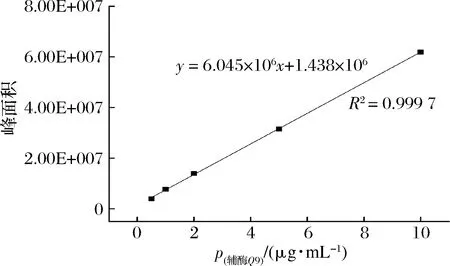
圖2 峰面積與輔酶Q9濃度的線性關系Fig.2 Linear relationship between peak area and Q9 concentration
1.3.6 細胞膜脂肪酸含量的測定
收集發酵一定時間的菌體,經過皂化、甲基化、萃取、堿洗等前處理步驟得到脂肪酸[19],用于氣質聯用分析。GC-MS分析條件參考文獻[19]。
1.3.7 胞內氨基酸的測定
取適量發酵液,8 000×g,離心10 min收集菌體,用PBS緩沖液(pH 7.0)洗滌2次。重懸于1 mL 10 g/L的三氯乙酸中,37 ℃放置10 min,煮沸15 min,8 000×g離心10 min,除去細胞碎片,上清液經0.22 μm水膜過濾,取400 μL上清液用于氨基酸分析[20]。
2 結果與分析
2.1 兩株菌分批發酵過程比較
在7.5 L發酵罐中進行A.pasteurianusCICIM B7003和CICIM B7003-02的分批發酵。圖3-A為A.pasteurianusCICIM B7003的發酵過程曲線,親本菌株接入發酵罐后存在20 h左右的延滯期,隨后進入快速產酸階段,20~70 h左右進入快速產酸期,發酵80 h停止產酸,34 h之后溶氧需求達到最大,發酵期間因底物乙醇和產物乙酸的雙重抑制作用,菌株進入對數生長期中期,最大比生長速率μmax僅為0.106 h-1,發酵結束,生物量為0.62 g/L,比突變株高37.8%,最終酸產量達51.6 g/L,發酵過程平均產酸速率為0.645 g/(L·h),發酵結束乙醇殘留5.69 g/L,乙醇對乙酸的轉化率為87.7%。相比于親本菌株,如圖3-C所示,突變株種子液接入發酵罐后迅速開始菌體生長和乙酸發酵過程,無延滯期,8 h左右溶氧下降到4%左右,隨后保持不變,58 h左右溶氧逐漸上升,最終發酵66 h結束,生物量為0.45 g/L,乙酸產量達55.2 g/L,發酵過程產酸速率較平穩,平均產酸速率為0.836 g/(L·h),發酵期間最大比生長速率μmax為0.121 h-1,發酵結束幾乎無乙醇殘留,乙醇對乙酸的轉化率達86.3%。

A-CICIM B7003發酵過程曲線;B-CICIM B7003能量代謝曲線;C-CICIM B7003-02發酵過程曲線;D-CICIM B7003-02能量代謝曲線圖3 A. pasteurianus CICIM B7003和CICIM B7003-02的分批發酵過程比較Fig.3 Comparison of batch fermentation process of A. pasteurianus CICIM B7003 and CICIM B7003-02
進一步對其胞內能量代謝分析可知,如圖3-B所示,親本株胞內ATP含量10 h后快速上升,29 h達到最大值16.25 μmol/g (菌體干重),如圖3-A所示,此時比生長速率在10 h達到最大值0.106 h-1,產酸速率為0.9 g/(L·h),發酵結束前ATP含量為0.055 μmol/g (菌體干重),如圖3-D所示,突變株胞內ATP在8 h左右達到最大值18 μmol/g(菌體干重),此時的比生長速率為最大值0.121 h-1,產酸速率為1.215 g/(L·h),隨后ATP水平迅速下降,在20 h后出現短暫的上升后又迅速下降,發酵結束前ATP含量為0.08 μmol/g(菌體干重),發酵初期乙酸濃度較低,主要利用三羧酸循環來供能,能量以ATP的形式儲存,乙醇快速氧化階段,主要利用乙醇呼吸鏈產能[18],產生的能量不僅需要供給菌體生長,還需供給蛋白抵抗酸脅迫下的不利環境。當乙酸擴散到細胞中時,ATP依賴性ABC轉運蛋白泵出乙酸,維持胞內的pH穩態,與GrpE和DnaJ類似,DnaK等分子伴侶蛋白消耗ATP以確保蛋白質在酸性壓力下正確折疊,ATP被大量消耗[21-23]。
2.2 發酵過程中細胞膜脂肪酸的變化
細胞膜與菌體能量傳導與維持細胞內微環境的穩定性密切相關,細胞膜脂肪酸組成的變化通常被認為是微生物細胞對外界毒性化合物的適應性反應[24]。實驗測定了24,48和72 h的細胞膜脂肪酸組成,如圖4所示。
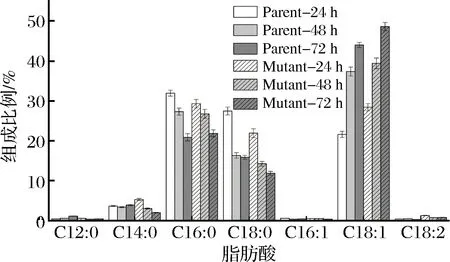
圖4 發酵過程中細胞膜脂肪酸組成變化Fig.4 Membrane fatty acids change of A. pasteurianusCICIM B7003 and CICIM B7003-02 during fermentation process
A.pasteurianusCICIM B7003和CICIM B7003-02發酵過程中的飽和脂肪酸主要由C12∶0 (月桂酸),C14∶0 (肉豆蔻酸),C16∶0 (棕櫚酸),C18∶0 (硬脂酸)組成,不飽和脂肪酸主要是C16∶1 (棕櫚油酸),C18∶1 (十八碳烯酸),C18∶2 (十八碳二烯酸)。棕櫚酸,硬脂酸和十八碳烯酸占據了A.pasteurianusCICIM B7003和CICIM B7003-02細胞膜脂肪酸大部分的比例。隨著發酵過程的進行,醋酸不斷積累,飽和脂肪酸,主要是棕櫚酸和硬脂酸的含量逐漸降低,不飽和脂肪酸,主要是十八碳一烯酸的比例不斷升高。發酵72 h棕櫚酸含量由24 h的31.92%(29.28%)降低到最終的20.86%(21.86%),硬脂酸的含量由發酵24 h的27.45% (21.91%)降低到發酵72 h的15.59%(11.88%)。十八碳一烯酸的含量由24 h的21.59% (28.4%)上升到發酵結束前的43.58% (48.7%),此外,發酵過程中A.pasteurianusCICIM B7003-02的棕櫚酸和硬脂酸等飽和脂肪酸比A.pasteurianusCICIM B7003低25.2%~33.8%,突變株中十八碳烯酸等不飽和脂肪酸比親本株高10.4%~31.5%,除此之外的其他脂肪酸幾乎無明顯變化。乙酸存在下,不飽和脂肪酸(順式異油酸)的增加形成更加流動的細胞膜,因為異油酸積聚會導致細胞膜流化[7]。
2.3 菌體胞內氨基酸含量變化
在高酸度的惡劣環境下,谷氨酸、天冬氨酸、脯氨酸、丙氨酸等氨基酸在醋酸菌細胞生長代謝和細胞存活中起重要作用,其增強酸脅迫損傷引起的DNA合成和DNA修復,促進細胞膜不飽和脂肪酸合成穩定性和完整性,改善細胞膜的流動性,穩定性和完整性[13-14],氨基酸脫羧形成氨降低胞內pH[25]。本實驗考察了突變菌株和親本菌株在發酵過程中的胞內氨基酸含量的變化,如圖5所示。

圖5 發酵過程中胞內氨基酸含量變化Fig.5 Changes in intracellular pools of amino acids changes in intracellular pools of amino acids during fermentation
發酵初期,胞內氨基酸含量普遍較高,隨著酸度的提升,胞內氨基酸含量不斷下降,在17種氨基酸中,天冬氨酸、谷氨酸、丙氨酸占據了大部分。在發酵的初始階段,24 h時突變株的胞內氨基酸大部分含量都比親本株高,例如突變株中谷氨酸和天冬氨酸的含量分別為2.58和1.03 μg/mg (蛋白),相比于親本株的2.33和0.87 μg/mg(蛋白)高10.7%和18.3%;但在72 h時,親本株中含量為0.56和1.02 μg/mg(蛋白),突變株中為0.37和0.80 μg/mg(蛋白)。因此在低酸度下氨基酸可能主要用于供給生長代謝,到發酵中后期,細胞活性相對較弱,發酵液中酸度較高,突變株細胞利用胞內氨基酸增強脫氨作用,增加細胞內氨濃度,從而維持細胞內pH的穩定性[14]。
2.4 ADH、ALDH酶活以及輔酶Q9含量變化
醋酸菌利用吡咯喹啉依賴性醇脫氫酶(PQQ-ADH)和乙醛脫氫酶(ALDH)將乙醇轉化為乙酸,它們是乙醇呼吸鏈中的關鍵酶,同時,PQQ-ADH的活性與乙酸耐受性有關[4]。如圖6-A所示,ADH酶活性和ALDH酶活性在發酵過程中都呈現先上升后下降的趨勢,其活性與產酸速率正相關。在親本株中,由于延滯期的存在,發酵初始酶活水平較低,隨著發酵的進行,最高酶活出現在45 h,ADH活性為7.21 U/mg,ALDH活性為5.84 U/mg,突變株的最高酶活出現在36 h,ADH活性為9.16 U/mg,ALDH活性為6.73 U/mg。

圖6 發酵過程中ADH和ALDH酶活以及輔酶Q9含量的變化Fig.6 Changes of ADH and ALDH activity and coenzyme Q9 content during fermentation process
輔酶Q9是乙醇呼吸鏈中可自由穿梭的電子傳遞載體,其接受ADH的輔酶PQQ的電子被還原成Q9H2,隨后被末端氧化酶Cyto氧化,其含量與乙醇呼吸鏈的強度有直接關系,如圖6-B所示,整個發酵過程中突變株的輔酶Q9含量一直高于親本株,發酵24 h時由于剛進入生長期,親本株的輔酶Q9含量只有0.91 mg/g菌體干重,突變株處于對數生長期,輔酶Q9含量為3.71 mg/g (菌體干重),發酵48 h,輔酶Q9含量在2株菌中含量都達到了最大值,分別為3.09和5.24 mg/g(菌體干重),發酵72 h時,此時處于發酵后期,發酵液中酸積累較多,乙醇呼吸鏈活性減弱,輔酶Q9含量相比48 h降低,分別為2.36和4.19 mg/g (菌體干重)。由此可見,ADH,ALDH酶活以及輔酶Q9含量的高低直接反映了乙醇呼吸鏈在發酵過程中的動態調控過程。
2.5 半連續發酵特性比較
底物乙醇初始濃度過高時,會對菌體產生抑制作用,所以工業中一般采用半連續發酵工藝來進行醋酸發酵。主要分為發酵啟動,半連續發酵適應階段,半連續發酵生產階段。如圖7所示,圖7-A為突變株的半連續發酵,發酵啟動時間為46 h,經過1次乙醇和新鮮培養基補料,達到最終酸度70.6 g/L,啟動后的第1次補料延滯期較長,為15 h左右,隨后進入快速產酸階段,經過一兩個不穩定的批次后進行穩定發酵,發酵周期33 h,平均產酸速率1.01 g/(L·h),乙醇對醋酸的轉化率為81.4%。
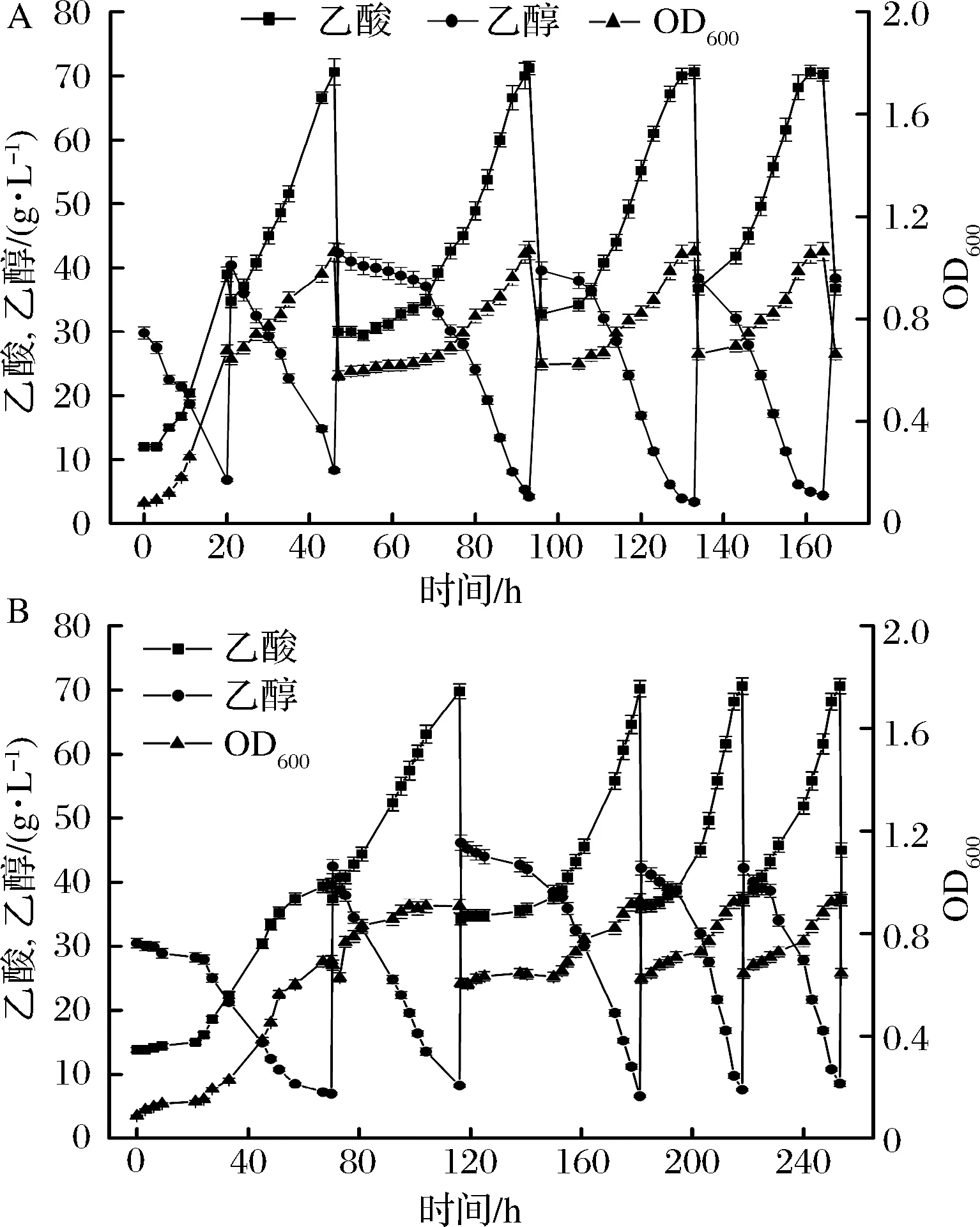
A-突變株的半連續發酵;B-親本株的半連續發酵圖7 兩株菌半連續發酵特性比較Fig.7 Fermentation characteriatics of A. pasteurianusCICIM B7003-02 and CICIM B7003 in repeated batch method
圖7-B中,種子液接入發酵罐后延滯期很長,116 h才結束了發酵啟動過程,經過一兩次適應階段后,發酵周期為35 h,平均產酸速率為0.87 g/(L·h),乙醇對醋酸的轉化率為76.7%,轉化率較低可能是由于半連續發酵中底物乙醇的不斷積累脅迫菌體導致利用乙醇不充分,另外發酵時間過長,乙醇的揮發降低了利用效率,誘變菌種充分縮短了發酵周期,特別是半連續發酵啟動階段。后續可以利用營養鹽進一步縮短發酵啟動時間。
3 討論
醋酸發酵是產物抑制底物轉化的典型模型,醋酸菌的一個突出和必要的生理特征是它們能夠在醋酸發酵的高醋酸條件下存活,本文從2個方面解析醋酸菌發酵耐酸特性,一方面通過親本菌株和突變菌株的發酵特性對比,包括分批發酵和重復補料分批發酵,親本株的延滯期較長,但后期產酸速率較快,并且分批發酵中親本株最終生物量高出突變株37.8%;醋酸發酵中,菌體生長狀態直接影響產酸速率,突變株中的生長代謝的基因突變可能限制了菌株的過度生長。突變株在發酵初始就擁有較高胞內ATP水平,初始乙酸可能誘導了其發酵啟動過程,導致大量ATP積累[22],保證了其快速發酵啟動過程,親本株依靠生物量的積累完成胞內ATP積累,達到最高ATP水平進入對數生長期和高產酸速率階段。
另一方面,親本株對發酵環境適應度差,考慮到可能是胞內微環境的影響,因此又對發酵過程中菌體細胞膜脂肪酸組成,胞內ATP含量,胞內氨基酸含量,乙醇脫氫酶(ADH),乙醛脫氫酶(ALDH)酶活以及輔酶Q9含量進行了分析,乙醇呼吸鏈ADH、ALDH酶活以及輔酶Q9含量,與發酵過程中產酸速率呈正相關,ATP含量與比生長速率呈正相關,突變株處于最高活性時的生理指標都明顯優于親本菌株。不飽和脂肪酸,特別是十八碳烯酸在發酵過程中不斷上升,飽和脂肪酸包括棕櫚酸和硬脂酸含量不斷下降,不飽和度的升高增強了細胞膜的流動性[26]。谷氨酸和天冬氨酸也被報道參與耐酸性,突變株胞內谷氨酸和天冬氨酸多10.7%和18.3%,其在發酵過程中不斷減少,主要參與增強磷酸戊糖和NADPH的生成,維持細胞內氧化還原平衡和細胞活力,增強氨基酸的脫氨作用,增加細胞內氨濃度,維持細胞內pH的穩定性[14],這不僅為食品工業選擇耐酸菌株提供了有效途徑,而且提供新的策略來提高該巴氏醋酸桿菌的工業實用性。
