小麥 TaCP3基因的克隆及其參與非生物脅迫的表達(dá)分析
2019-06-14 07:10:26張炳慧楊明明王憲國(guó)何慶夢(mèng)姚曉露李曉燕趙萬(wàn)春
麥類作物學(xué)報(bào) 2019年5期
張炳慧,高 翔,2,楊明明,王憲國(guó),何慶夢(mèng),姚曉露,李曉燕,趙萬(wàn)春,2,董 劍,2
(1.西北農(nóng)林科技大學(xué)農(nóng)學(xué)院,陜西楊凌 712100; 2.陜西省小麥工程技術(shù)研究中心/陜西省小麥新品種培育工程研究中心,陜西楊凌 712100)
半胱氨酸蛋白酶(CP)是植物中重要的蛋白酶家族之一,廣泛參與植物的各種生理過(guò)程,比如貯存蛋白的沉積和降解、對(duì)非生物脅迫和生物脅迫的響應(yīng)、植物的衰老和細(xì)胞程序性死亡(PCD)等[1]。Rawlings等[2]把半胱氨酸蛋白酶大致分為15個(gè)家族,分屬于5大類。包括C1:Papain,木瓜蛋白酶;C2:Calpain,鈣依賴半胱氨酸蛋白酶;C12:催化蛋白去泛素化的半胱氨酸蛋白酶;C13:Legumain,豆類天冬氨酸蛋白內(nèi)切酶;C14:天冬氨酸特異性的半胱氨酸蛋白酶;C19:蛋白酶體。大多數(shù)植物半胱氨酸蛋白酶屬于Papain(木瓜蛋白酶)(C1)和Legumain(豆類天冬氨酸蛋白內(nèi)切酶)(C13)家族。Papain-like蛋白家族(C1A)是植物半胱氨酸蛋白酶的主要成員,也是研究得最為詳盡的一個(gè)家族[3],近年來(lái)在擬南芥[4-5]、大豆[6-7]煙草[8]和水稻[9]等植物中對(duì)這類基因已有大量研究,但在小麥中報(bào)道較少。Papain家族在酶前體中已經(jīng)形成大小相當(dāng)?shù)?個(gè)結(jié)構(gòu)域,在這2個(gè)結(jié)構(gòu)域構(gòu)成的裂隙底部是催化活性位點(diǎn)[10]。Papain家族的各個(gè)成員含有由保守氨基酸殘基組成的催化三聯(lián)體,它們?cè)陔逆溕系呐帕许樞蚴荂ys-his-Asn/Asp[11],Gln殘基對(duì)其催化作用的發(fā)揮也很重要。酶前體通過(guò)分子內(nèi)或分子間蛋白水解而激活。Zang等[12]的研究表明,小麥中TaCP為水分脅迫誘導(dǎo)表達(dá)的上調(diào)基因可以受脫水脅迫、鹽脅迫及低溫脅迫的誘導(dǎo)表達(dá)。馬巖巖等[13]研究發(fā)現(xiàn),柑橘半胱氨酸蛋白酶基因CsCysP也可以受脫水脅迫、鹽脅迫及低溫脅迫的誘導(dǎo)表達(dá)。
半胱氨酸蛋白酶家族基因目前在小麥中尚未被深入研究,主要功能還不明確。本課題組在前期工作中從小麥抗旱品種西農(nóng)538中克隆出一個(gè)半胱氨酸蛋白酶基因,將其命名為TaCP3,本試驗(yàn)對(duì)其進(jìn)行生物信息學(xué)分析、蛋白誘導(dǎo)表達(dá)和非生物脅迫處理,以期為在小麥中進(jìn)一步研究半胱氨酸蛋白酶的功能提供參考。
1 材料與方法
1.1 材料與處理
普通小麥品種西農(nóng)538由國(guó)家小麥改良中心楊凌分中心品質(zhì)實(shí)驗(yàn)室保存并提供。待小麥生長(zhǎng)至兩葉一心期,分別用20% PEG-6000、250 mmol·L-1NaCl進(jìn)行干旱和高鹽脅迫處理,并分別放置在4 ℃和40 ℃的光照培養(yǎng)箱進(jìn)行低溫和高溫脅迫處理。從脅迫0 h開(kāi)始取樣(小麥葉片),除高溫脅迫處理在0、0.25、0.5、1、2、3、4和6 h取樣外,其余脅迫均在0、1、3、6、12、24、48和 72 h取樣。所有樣品取完后均迅速放入液氮中,并在速凍后放入-80 ℃冰箱保存?zhèn)溆谩?/p>
1.2 目的基因的克隆
以室內(nèi)培養(yǎng)至兩葉一心期的西農(nóng)538幼嫩葉片為材料,采用微量CTAB[14]法提取基因組DNA。根據(jù)擬南芥At1g47128序列,在NCBI上進(jìn)行序列比對(duì),結(jié)合URGI數(shù)據(jù)庫(kù)比對(duì)結(jié)果,選取小麥基因組中與At1g47128相似性最高的3B染色體上的序列,命名為TaCP3。根據(jù)其序列設(shè)計(jì)特異性引物CP3-F(ATGGGGCAGCTCAGC AAGAAG)/CP3-R(TCACTGCGACTCCCATG C,以西農(nóng)538的基因組DNA為模板進(jìn)行擴(kuò)增反應(yīng),體系與程序參考ZXEs Taq Master Mix說(shuō)明書(shū)(康為世紀(jì),北京),其中循環(huán)數(shù)為30,退火溫度為62 ℃,延伸時(shí)間為75 s。PCR產(chǎn)物用1%的瓊脂糖凝膠電泳進(jìn)行檢測(cè),用DNA凝膠回收純化試劑盒(天根,北京)回收目的片段。將回收的目的片段連接入pEASY-T1克隆載體(全式金,北京),轉(zhuǎn)化進(jìn)Transl-T1感受態(tài)(全式金,北京),涂于含有適量Amp的LB平板,37 ℃過(guò)夜培養(yǎng),以通用引物M13F/M13R對(duì)單克隆菌落進(jìn)行PCR擴(kuò)增,篩選陽(yáng)性克隆,并送北京奧科鼎盛生物科技有限公司進(jìn)行測(cè)序。
1.3 生物信息學(xué)分析
利用ExPASy-ProtParam(https://web.expasy.org/protparam/)進(jìn)行蛋白質(zhì)基本理化性質(zhì)預(yù)測(cè);用NPS-SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)預(yù)測(cè)TaCP3蛋白序列的二級(jí)結(jié)構(gòu);用ExPASy-SWISS MODELL(https://www.swissmodel.expasy.org/interactive)預(yù)測(cè)蛋白質(zhì)的三級(jí)結(jié)構(gòu);用CBS-SignalP 3.0 Server(http://www.cbs.dtu.dk/services/SignalP-3.0/)預(yù)測(cè)信號(hào)肽;用CBS-TargetP 1.1 Server(http://www.cbs.dtu.dk/services/TargetP/)進(jìn)行亞細(xì)胞定位。下載其他植物的半胱氨酸蛋白酶的蛋白序列,用DNAMAN進(jìn)行多重序列比對(duì);用MEGA 7軟件依據(jù)Neighbor-Joining法構(gòu)建系統(tǒng)進(jìn)化樹(shù)。
1.4 目的基因的誘導(dǎo)表達(dá)
根據(jù)測(cè)序得到的TaCP3基因序列,設(shè)計(jì)一對(duì)去除信號(hào)肽序列的表達(dá)引物RDd-F/RDd-R。以陽(yáng)性克隆菌液為模板,用RDd-F/RDd-R進(jìn)行PCR擴(kuò)增反應(yīng),體系和程序參考KOD FX說(shuō)明書(shū)(TOYOBO,日本),其中循環(huán)數(shù)為30,退火溫度為62 ℃,延伸時(shí)間為75 s。同1.2節(jié)的方法進(jìn)行目的片段的回收、連接和轉(zhuǎn)化,送測(cè)序。將上述測(cè)序正確的pEASY-T1/TaCP3載體中的目的片段用內(nèi)切酶BamHI和HindⅢ進(jìn)行雙酶切,瓊脂糖凝膠電泳后用回收試劑盒純化,插入同樣雙酶切的大腸桿菌目的表達(dá)載體pcold-TF(全式金,北京)中,經(jīng)T4 DNA連接酶16 ℃連接過(guò)夜,轉(zhuǎn)化至感受態(tài)大腸桿菌DH5α,在含有Amp抗性的LB平板上培養(yǎng),挑取陽(yáng)性克隆,PCR、酶切鑒定。正確后將插入順序正確的陽(yáng)性單克隆提取質(zhì)粒,轉(zhuǎn)化于EscherichiacoliBL21(DE3)表達(dá)感受態(tài)(全式金,北京),培養(yǎng)后挑選陽(yáng)性單克隆菌落,以備重組蛋白誘導(dǎo)表達(dá)所用。將上一步得到的陽(yáng)性單克隆菌落接種于LB液體培養(yǎng)基(含100 mg·mL-1Amp),于37 ℃、200 r·min-1過(guò)夜培養(yǎng)。將培養(yǎng)物按1∶100的比例接種于5 mL含LB的液體培養(yǎng)基(含100 mg·mL-1Amp)中震蕩培養(yǎng)至菌液OD600=0.5左右,取1 mL菌液作為陰性對(duì)照,向剩下的菌液中加入終濃度為0.6 mmol·L-1的IPTG,20 ℃誘導(dǎo)表達(dá)20 h。用12%的SDS-PAGE分析鑒定蛋白表達(dá)結(jié)果。
1.5 誘導(dǎo)表達(dá)產(chǎn)物的Western blot分析
誘導(dǎo)表達(dá)產(chǎn)物經(jīng)SDS-PAGE電泳后進(jìn)行轉(zhuǎn)膜。在60 V電壓下濕轉(zhuǎn)75 min,然后取出NC膜于干凈的玻璃容器中,TBS洗膜3次,每次15 min;用新配置的封閉液(含5%脫脂奶粉)室溫?fù)u床封閉2 h,棄封閉液,TBS和TBST洗膜各2次,每次15 min;以鼠抗his(1∶10 000封閉液稀釋)抗體為一抗(全式金,北京),室溫?fù)u床孵育1 h,棄一抗,TBS和TBST洗膜各2次,每次15 min;以HRP標(biāo)記的羊抗鼠IgG(1∶5 000封閉液稀釋)為二抗(全式金,北京),室溫?fù)u床孵育1 h,棄二抗,TBS和TBST洗膜各2次,每次15 min。
1.6 實(shí)時(shí)熒光定量PCR(qRT-PCR)
用RNAprep Pure植物總RNA提取試劑盒(天根,北京)提取RNA,用FastKing cDNA第一鏈合成試劑盒(天根,北京)進(jìn)行cDNA第一鏈的合成。將所得cDNA于-20 ℃保存。用SuperReal 彩色熒光定量預(yù)混試劑盒(天根,北京)在Q7熒光實(shí)時(shí)定量PCR儀(ABI,美國(guó))上進(jìn)行qRT-PCR,分析TaCP3基因在上述處理各個(gè)時(shí)間點(diǎn)的表達(dá)情況,反應(yīng)體系與程序參照說(shuō)明書(shū)。以 18S RNA為內(nèi)參基因,每個(gè)樣品設(shè)置3個(gè)生物學(xué)重復(fù),用2-ΔΔCt[15]法計(jì)算脅迫處理的樣品和對(duì)照組樣品中目標(biāo)基因的相對(duì)表達(dá)量。
2 結(jié)果與分析
2.1 目的基因的克隆與序列分析結(jié)果
以西農(nóng)538基因組DNA為模板,利用引物RD-F/RD-R進(jìn)行PCR擴(kuò)增。擴(kuò)增產(chǎn)物經(jīng)1%瓊脂糖凝膠電泳檢測(cè)后,得到約1 100 bp的單一條帶。將目的條帶回收、克隆和測(cè)序后,得到長(zhǎng)度為1 125 bp的序列,比對(duì)后確定成功擴(kuò)增了TaCP3的序列全長(zhǎng)。序列分析表明,TaCP3編碼374個(gè)氨基酸,相對(duì)分子質(zhì)量約為40.5 kD,等電點(diǎn)為6.76。在NCBI蛋白數(shù)據(jù)庫(kù)中進(jìn)行保守結(jié)構(gòu)域分析,結(jié)果顯示TaCP3的N端包含一個(gè)組織蛋白酶前肽抑制劑結(jié)構(gòu)域,位于TaCP3的第46~102 位氨基酸上,在部分木瓜蛋白酶家族中也存在這一結(jié)構(gòu)域[16];含有一個(gè)木瓜蛋白酶家族(papain C1) 保守結(jié)構(gòu)域,位于TaCP3的第138~354位氨基酸。NPS-SOPMA預(yù)測(cè)結(jié)果表明,該蛋白主要以無(wú)規(guī)則卷曲(47.33%)和α-螺旋(33.16%)為主,含少量β-轉(zhuǎn)角(5.08%)。ExPASy-SWISS MODELL分析結(jié)果表明,5egw.1.B序列與TaCP3氨基酸序列的第33~360個(gè)殘基的一致性達(dá)46%,且5egw.1.B序列編碼蛋白也是半胱氨酸蛋白酶家族蛋白,具備同源建模的條件。得到TaCP3基因所編碼蛋白的三維結(jié)構(gòu)模型。CBS-SignalP 3.0預(yù)測(cè)發(fā)現(xiàn),TaCP3在N-端含有信號(hào)肽,且切割位點(diǎn)位于第28位和第29位氨基酸之間。結(jié)合CBS-TargetP 1.1 Server的亞細(xì)胞定位分析,表明這個(gè)蛋白可能是分泌型蛋白、胞外蛋白。在Uniprot蛋白序列庫(kù)中對(duì)TaCP3的氨基酸序列進(jìn)行NCBI-BLASTP同源序列比對(duì)分析,下載來(lái)自不同植物的半胱氨酸蛋白酶蛋白序列9條,用DNAMAN進(jìn)行蛋白質(zhì)多重序列比對(duì)(圖1),用MEGA7軟件構(gòu)建系統(tǒng)進(jìn)化樹(shù)。結(jié)果顯示,TaCP3與同族禾本科的大麥(GenBank登錄號(hào)M0Z3Q3)和山羊草(GenBank登錄號(hào)N1R0h7)同源關(guān)系最近(圖2),序列相似性分別為88%和89%。
2.2 TaCP3原核蛋白表達(dá)
通過(guò)優(yōu)化表達(dá)條件,在0.6 mmol·L-1IPTG、20 ℃誘導(dǎo)條件下誘導(dǎo)20 h目的蛋白表達(dá)達(dá)到高峰。表達(dá)蛋白經(jīng)12% SDS-PAGE電泳顯示相對(duì)分子質(zhì)量約為92 kD(40 kD+52 kD,因?yàn)榈鞍踪|(zhì)標(biāo)簽的融合表達(dá),因此所得的表達(dá)產(chǎn)物的分子量比目的蛋白大),與推測(cè)蛋白分子質(zhì)量大小相符(圖3,圖4)。
粗體下劃線為半胱氨酸蛋白酶保守區(qū),箭頭指向TaCP3蛋白質(zhì)序列 。
Multiple sequence alignment of proteins,underlined as a conserved region of cysteine protease; arrows point to TaCP3 protein sequence.
圖1 TaCP3 蛋白的多重序列比對(duì)
Fig.1 Multiple sequence alignment of TaCP3 proteins
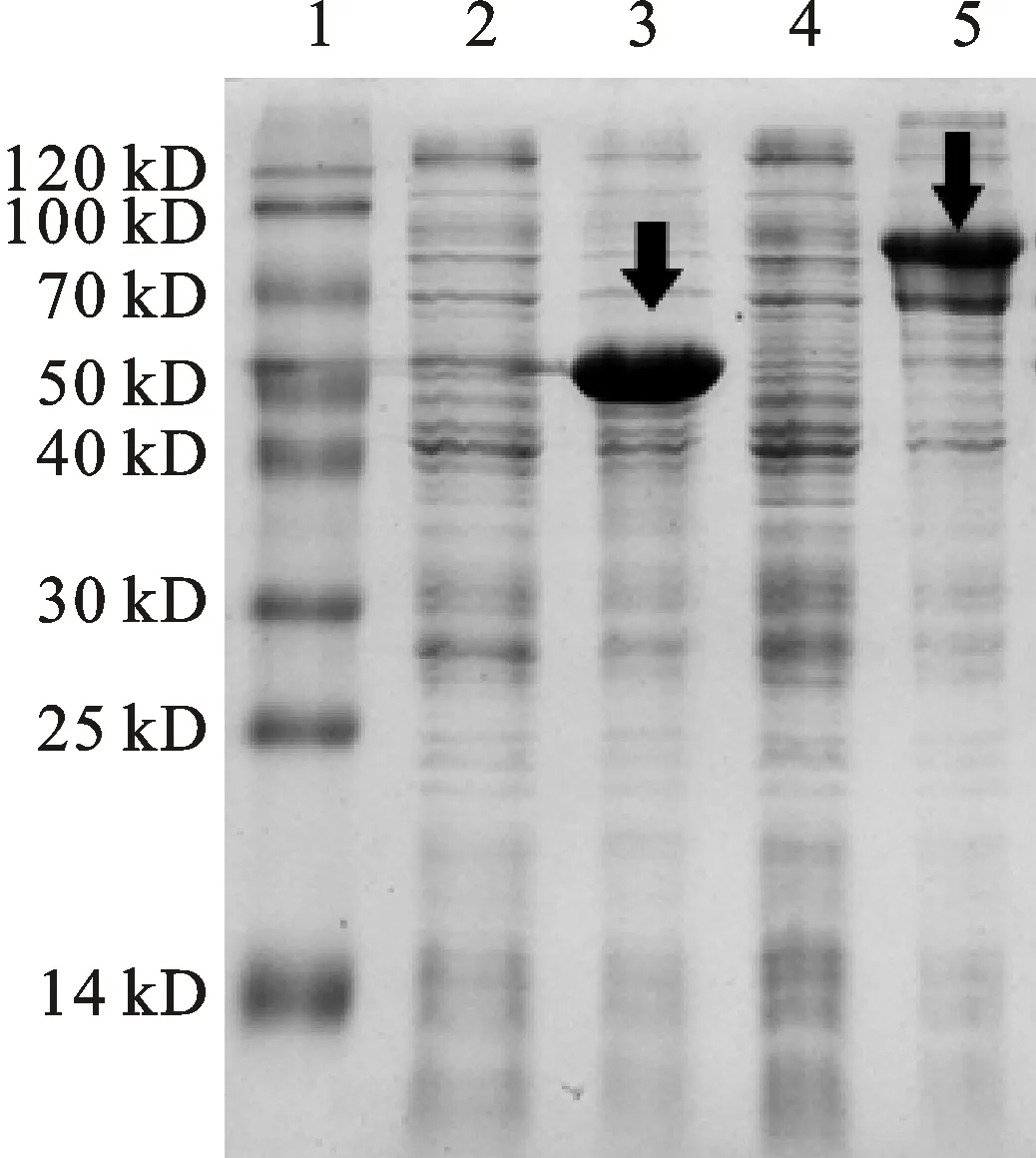
1:蛋白質(zhì)分子質(zhì)量標(biāo)準(zhǔn);2:未誘導(dǎo)的空載體;3:IPTG誘導(dǎo)的空載體;4:未誘導(dǎo)的TaCP3重組質(zhì)粒表達(dá)產(chǎn)物;5:IPTG誘導(dǎo)的TaCP3重組質(zhì)粒表達(dá)產(chǎn)物。
1:Blue Plus II Protein Marker; 2:Protein of non-recombinant plasmid without induction; 3:Protein of non-recombinant plasmid after induced by IPTG; 4:Recombinant plasmid of TaCP3 without induction; 5:Recombinant plasmid of TaCP3 induced by IPTG.
圖3 TaCP3重組質(zhì)粒的SDS-PAGE分析
Fig.3 Analysis TaCP3 of recombinant by SDS-PAGE
2.3 脅迫處理下 TaCP3基因的表達(dá)
在干旱脅迫(20% PEG-6000)處理下,TaCP3在前6 h內(nèi)表達(dá)量下降,于12 h表達(dá)量驟升至初始表達(dá)量的近10倍,24 h時(shí)回落至2倍初始表達(dá)量,并持續(xù)下降到72 h。在高鹽脅迫(NaCl 250 mmol·L-1)處理下,TaCP3在1 h時(shí)表達(dá)量略降低,3 h時(shí)表達(dá)量上升至最大值,然后逐漸降低,至48 h時(shí)表達(dá)量低于初始表達(dá)量,在72 h時(shí)又突升接近最高表達(dá)量。冷害脅迫(4 ℃)處理下,除48 h表達(dá)量高至初始表達(dá)量,其余時(shí)間均低于初始表達(dá)量,并在24 h時(shí)達(dá)到最低,總體呈現(xiàn)“下降-上升-下降-上升-下降”趨勢(shì)。高溫脅迫(40 ℃)處理下,TaCP3表達(dá)量先下降后上升至初始表達(dá)量,然后再下降再上升至初始表達(dá)量,在 6 h 升至最高表達(dá)量。
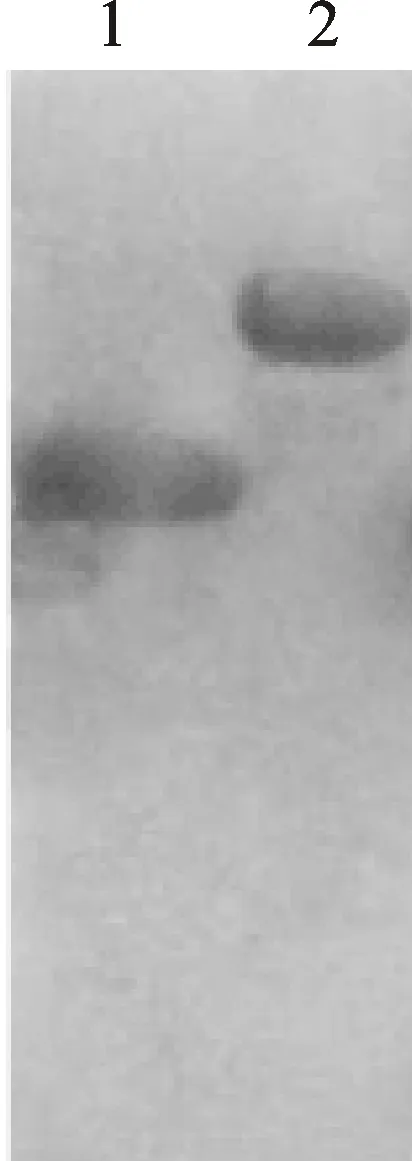
1:IPTG誘導(dǎo)的空載體;2:IPTG誘導(dǎo)的TaCP3重組質(zhì)粒表達(dá)產(chǎn)物。
1:Protein of non-recombinant plasmid induced by IPTG; 2:Recombinant plasmid of TaCP3 induced by IPTG.
圖4 基因誘導(dǎo)表達(dá)產(chǎn)物蛋白質(zhì)印跡法檢測(cè)結(jié)果
Fig.4 Analysis of recombinant plasmids by Western blot
3 討 論
本研究首次在小麥中克隆得到TaCP3基因,其氨基酸序列N端有28個(gè)氨基酸殘基編碼的信號(hào)肽,C端具有木瓜蛋白酶亞家族的保守結(jié)構(gòu)域。與其他的木瓜蛋白酶相似,TaCP3在保守區(qū)域內(nèi)也具有木瓜蛋白酶亞家族共有的活性位點(diǎn)Gln-Cys-his-Asn/Asp[17]。其與禾本科植物大麥和山羊草中半胱氨酸蛋白酶的相似性最高,分別為88%和89%,進(jìn)化分析表明,其與大麥和山羊草中半胱氨酸蛋白酶在進(jìn)化上的同源性也最高。Ao等[18]的研究也表明,禾本科的半胱氨酸蛋白酶基因是非常親近的,在進(jìn)化過(guò)程中某些物種中半胱氨酸蛋白酶基因的保守性以及這一特征可用于基因工程以改善植物的抗逆性。重組蛋白誘導(dǎo)表達(dá)結(jié)果顯示,含有終濃度為0.6 mmol·L-1IPTG的菌液,在20 ℃下誘導(dǎo)20 h是最佳的誘導(dǎo)條件。以上為進(jìn)一步研究TaCP3的酶學(xué)特征和生理功能打下了堅(jiān)實(shí)的基礎(chǔ)。
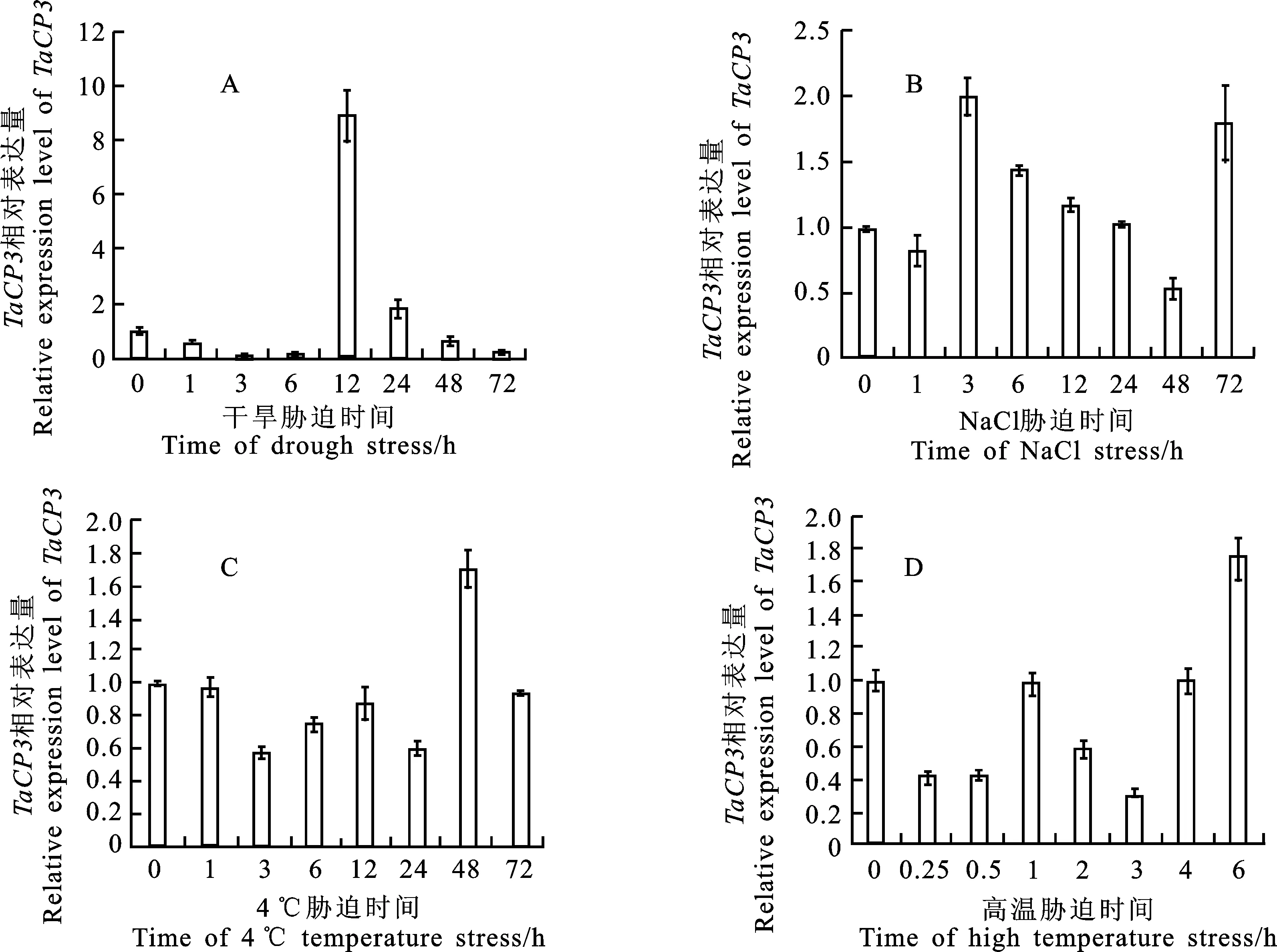
A:干旱脅迫(20% PEG-6000處理);B:高鹽脅迫(250 mmol·L-1NaCl處理);C:冷害脅迫(4 ℃);D:高溫脅迫(40 ℃)。
A:Drought stress(treated with 20% PEG6000); B:High salt stress(treated with 250 mmol·L-1NaCl);C:Cold stress(treated with 4 ℃); D:High temperature stress(treated with 40 ℃).
圖5 非生物脅迫下TaCP3的相對(duì)表達(dá)量
Fig.5 Relative expression level ofTaCP3gene under various abiotic stresses
實(shí)時(shí)熒光定量PCR結(jié)果顯示,TaCP3基因在干旱、高鹽、低溫和高溫脅迫下均可以被誘導(dǎo),其中能夠被干旱脅迫強(qiáng)烈誘導(dǎo)。雖然四種不同的脅迫條件下,TaCP3的響應(yīng)程度和時(shí)間有所不同,但是大體呈現(xiàn)為“下降-上升-下降-上升”的波動(dòng)表達(dá)模式。同時(shí),不論什么脅迫處理下,在0 h后時(shí)間節(jié)點(diǎn)的TaCP3表達(dá)量總是先降低。Ao等[18]的研究指出,在48 h干旱脅迫期間,MwCP在4 h表達(dá),并且表達(dá)水平在下降至6 h之前達(dá)到峰值;Jennifer和John[19]的研究發(fā)現(xiàn),當(dāng)豌豆植株受到水分脅迫時(shí),Cyp15a的表達(dá)增加,在不同的脅迫條件下Cyp15a的表達(dá)量不同。在方正武等[20]的研究中,F(xiàn)eRD21在干旱脅迫12 h的表達(dá)量也達(dá)到了最大值;而擬南芥中的RD19A和RD21A能夠被干旱和高鹽強(qiáng)烈誘導(dǎo)[3]。雖然在不同物種中,干旱處理下CP家族基因響應(yīng)達(dá)到表達(dá)量峰值的時(shí)間會(huì)有所不同,但是都是正向調(diào)控,會(huì)達(dá)到一個(gè)表達(dá)量的最大值。就抗旱性而言,Grudkowska和Zagdan的[21]試驗(yàn)結(jié)果表明,缺水時(shí),偶氮酪蛋白活性增強(qiáng),但在Kobra栽培品種(耐受性基因型)中活性增強(qiáng)顯著較低。以往的研究表明,對(duì)脫水反應(yīng),抗旱性不同的小麥品種在水分脅迫時(shí)的表現(xiàn)不同。半胱氨酸蛋白酶活性優(yōu)先增強(qiáng),但其活性與10個(gè)春小麥品種的脫水耐性水平呈負(fù)相關(guān)[22]。此外,已有許多試驗(yàn)證明,干旱誘導(dǎo)基因也可被高鹽和低溫所誘導(dǎo),表明這3種脅迫響應(yīng)之間存在相似的機(jī)制[3]。
本試驗(yàn)從模擬非生物脅迫入手,證明了TaCP3是可以響應(yīng)干旱脅迫、高鹽脅迫、低溫脅迫、高溫脅迫的。但是逆境的響應(yīng)是由許多基因調(diào)控的,因此半胱氨酸蛋白酶在響應(yīng)逆境時(shí)的具體作用機(jī)制仍需進(jìn)行進(jìn)一步的研究。同時(shí)本試驗(yàn)中TaCP3能夠響應(yīng)干旱脅迫,也能夠間接證明西農(nóng)538是一個(gè)抗旱品種。另外試驗(yàn)只是證明了TaCP3可以響應(yīng)四種非生物脅迫,并沒(méi)有與非抗品種進(jìn)行比較,不能得出TaCP3對(duì)這四種非生物脅迫的作用大小。以后可以設(shè)置抗性組和非抗組,同時(shí)進(jìn)行脅迫處理,比較TaCP3的表達(dá)量差異,也可參照Z(yǔ)ang等[12]將TaCP3轉(zhuǎn)入擬南芥(Arabidopsisthaliana)進(jìn)行抗旱性功能驗(yàn)證。
