棉花磷脅迫響應基因GhWRKY6克隆及功能分析
2019-06-14 02:27:34李智韓笑張一豪馬峙英楊召恩
棉花學報 2019年3期
關鍵詞:植物
李智 ,韓笑 ,張一豪 ,馬峙英 ,楊召恩 *
(1.中國農業科學院棉花研究所/ 棉花生物學國家重點實驗室,河南 安陽 455000;2.河北農業大學/ 棉花生物學國家重點實驗室河北基地,河北 保定 071001;)
在植物整個生命活動中磷元素是不可或缺的大量元素之一,對植物的生長發育有著極其重要的作用。磷元素除了是細胞結構物質磷脂和核酸的重要成分外,還參與糖、脂、蛋白質代謝以及蛋白磷酸化和去磷酸化等過程。此外,種子的萌發過程也需要含磷化合物作為磷源[1]。
磷酸根 (Pi) 是植物吸收磷的形式,主要以H2PO4-、HPO42-和 PO43-等形式存在,其中 H2PO4-最容易被植物吸收。雖然土壤中磷元素含量豐富,但多數以植物不能吸收的形式存在,導致有效磷的濃度降低,加之其又很容易被固定在土壤中,所以植物很容易處于缺磷環境中[2-3]。為了促進作物產量的提高而大量施用磷肥不僅會導致生產成本的增加還會對環境造成不利影響。因此,研究植物磷營養的調控通路,發掘及利用磷誘導的相關基因對提高作物耐低磷能力具有重要意義。
WRKY 轉錄因子是植物特有的轉錄因子,參與多種生物與非生物脅迫反應[4]。最近王琛等鑒定出陸地棉中18個Ⅱc 族WRKY 轉錄因子在棉花對枯萎病菌的抗性方面發揮重要作用[5],司愛君等研究報道GhWRKY40-1 在棉花干旱脅迫應答中發揮重要作用[6]。關于WRKY 轉錄因子參與磷信號途徑在擬南芥中報道較多。WRKY75 的表達受低磷誘導非常明顯,該基因的RNAi 干涉株系在低磷條件下影響植株對磷的吸收[7]。WRKY45 可與PHT1;1 啟動子內的兩個W-box結合調控PHT1;1 上調表達從而參與擬南芥對低Pi 脅迫的反應[8]。AtWRKY42 被研究證明參與磷信號轉導過程,對維持植物體內Pi 的動態平衡有調控作用[9]。過表達AtWRKY6 導致擬南芥對低磷脅迫敏感,降低了幼苗中磷元素的含量[10]。Gb-WRKY1 是最近報道的響應磷脅迫的棉花WRKY基因,其參與調控Pi 的運輸和活化過程,在擬南芥中過表達GbWRKY1 能夠緩解植物缺Pi 癥狀,但在Pi 缺乏條件下不會促進Pi 的吸收[11]。
為了發掘棉花中應答低磷脅迫的WRKY 轉錄因子,我們克隆到陸地棉GhWRKY6 基因的全長cDNA 序列,其核酸序列與擬南芥AtWRKY6[10]相似性較高是其同源基因,故推測GhWRKY6 在磷轉運調控方面有一定作用。本研究利用轉基因技術獲得擬南芥轉基因株系,通過在低磷脅迫條件下轉基因擬南芥表型的鑒定,初步確定了Gh-WRKY6 的功能,為解析GhWRKY6 基因在棉花低磷脅迫應答中的調控機制提供科學依據。
1 材料與方法
1.1 試驗材料
試驗中所用的野生型擬南芥是Col-0 型,wrky6 擬南芥突變體來自中國農業大學陳義芳實驗室。
Polymerase chain reaction (PCR)所用的 TKS高保真聚合酶、載體構建所用的T4 連接酶、限制性內切酶和cDNA 反轉錄試劑盒均購自TaKaRa(中國,大連)公司;氨芐、卡那霉素和利福平等抗生素購自Sigma 公司; 植物總RNA 提取試劑盒和植物總DNA 提取試劑盒購自天根生化科技有限公司(中國,北京);PCR 產物膠回收純化試劑盒和大腸桿菌感受態DH5α 購自全式金公司(中國,北京);引物合成及片段測序由北京金唯智公司完成。
1.2 試驗方法
1.2.1RNA 的提取。采用改良 CTAB 法[12]提取在大田中生長的陸地棉中棉所24 各組織的RNA,每個樣本3個重復,RNA 放置-80 ℃冰箱保存。
1.2.2GhWRKY6 基因的克隆。基于本實驗室未發表的RNA-seq 數據中發掘到了一個在非生物脅迫后表達量顯著上調的基因,該基因的核酸序列與擬南芥AtWRKY6 基因具有很高的相似性,故將其命名為GhWRKY6。根據GhWRKY6 基因編碼序列在primer premier5 軟件中設計目的基因擴增引物GhWRKY6-F和GhWRKY6-R (表1),PCR 擴增得到GhWRKY6 基因片段并回收連接到pMD○R18-T SIMPLE 載體,轉化大腸桿菌感受態DH5α,送PCR 鑒定的陽性菌到北京金唯智公司測序獲得正確序列的GhWRKY6 基因片段。
1.2.3生物信息學分析。使用DNAMAN 軟件進行核苷酸多序列分析和比對,使用MEGA6 軟件構建進化樹,利用CDD (http://www.ncbi,nlm.nihgov/Structure/cdd/wrpsb.cgi) 對蛋白保守結構域進行預測,使用 EXPASY (http://web.expasy.org/cgi/-bin/compute_pi/pi_tool)對蛋白質的相對分子質量(Mr)和理論等電點進行分析,使用SignalP 4.0 Server (http://www.cbs.dtu.dk/services/TMHMM/)進行跨膜結構域預測。
1.2.4GhWRKY6 的組織表達模式分析。為了進一步研究GhWRKY6 的表達模式,以陸地棉中棉所 24 的根、莖、葉、花瓣(開花當天)、胚珠(開花當天)和纖維(開花后5 d、開花后10 d、開花后15 d) 為材料提取 RNA,提取方法如 1.2.1 所述。qRT-RCR 操作過程及計算方法如1.2.7 所述。
1.2.535S::GhWRKY6 過表達載體的構建。Gh-WRKY6 基因片段和植物表達載體pCambia2300使用限制性內切酶XbaⅠ和AscⅠ完成雙酶切,酶切產物片段使用凝膠回收試劑盒回收,之后使用T4 連接酶連接構建完整的目的基因表達載體pCambia2300-GhWRKY6。使用電轉法[13]將構建好的過表達載體轉入農桿菌GV3101 菌株中以備轉化擬南芥。
1.2.6擬南芥的生長條件和低磷脅迫。使用營養土和蛭石比例為1:1 的混合土壤種植擬南芥植株,放置于人工氣候室(溫度19~20 ℃,濕度70%,光周期為 16 h 光 /8 h 黑暗)生長至收獲種子。
利用低磷 (10 μmol·L-1)MS 培養基模擬擬南芥的低磷脅迫環境,同時設置正常磷濃度的(1.25 mmol·L-1)MS 培養基作為對照。無論是擬南芥種子萌發的統計還是根長的統計都需要預先對種子進行滅菌處理。首先將種子在NaClO(50%)溶液中滅菌5 min,然后用無菌蒸餾水洗滌5~6 次,之后均勻鋪在正常MS 或低磷MS培養基。對于擬南芥種子萌發的統計是每天固定時間點統計種子萌發數目,連續統計1 周。對于擬南芥根部的觀察是先將滅菌的種子在正常MS培養基上生長1 周,之后轉入低磷或正常MS 培養基上繼續生長,所有培養基需要垂直放置。
需要注意的是擬南芥在轉入恒溫培養室之前要預先在4 ℃低溫、黑暗環境處理2 d,以保持種子萌發一致。
1.2.7RT-PCR 和qRT-PCR。采用 TaKaRa 公司的PrimeScript○RRT Reagent Kit with gDNA Eraser反轉錄試劑盒對RNA 完成反轉錄,具體步驟可參考試劑盒說明書。RT-PCR 的程序為:94 ℃5 min;98 ℃ 15 s,60 ℃ 15 s,68 ℃ 30 s,27個循環;68 ℃ 3 min;4 ℃保存。
Real-time PCR 反應體系和反應程序按照SYBR Premix Ex Taq (DRR041A) 熒光定量試劑盒說明書進行,使用的熒光定量PCR 儀器型號是ABI 7900,操作方法可參考分析儀說明書。棉花組織內參基因為GhHistone3,擬南芥組織內參基因為Atactin2,每個反應設置3個重復。獲得結果采用2-△△Ct計算基因的相對表達量。
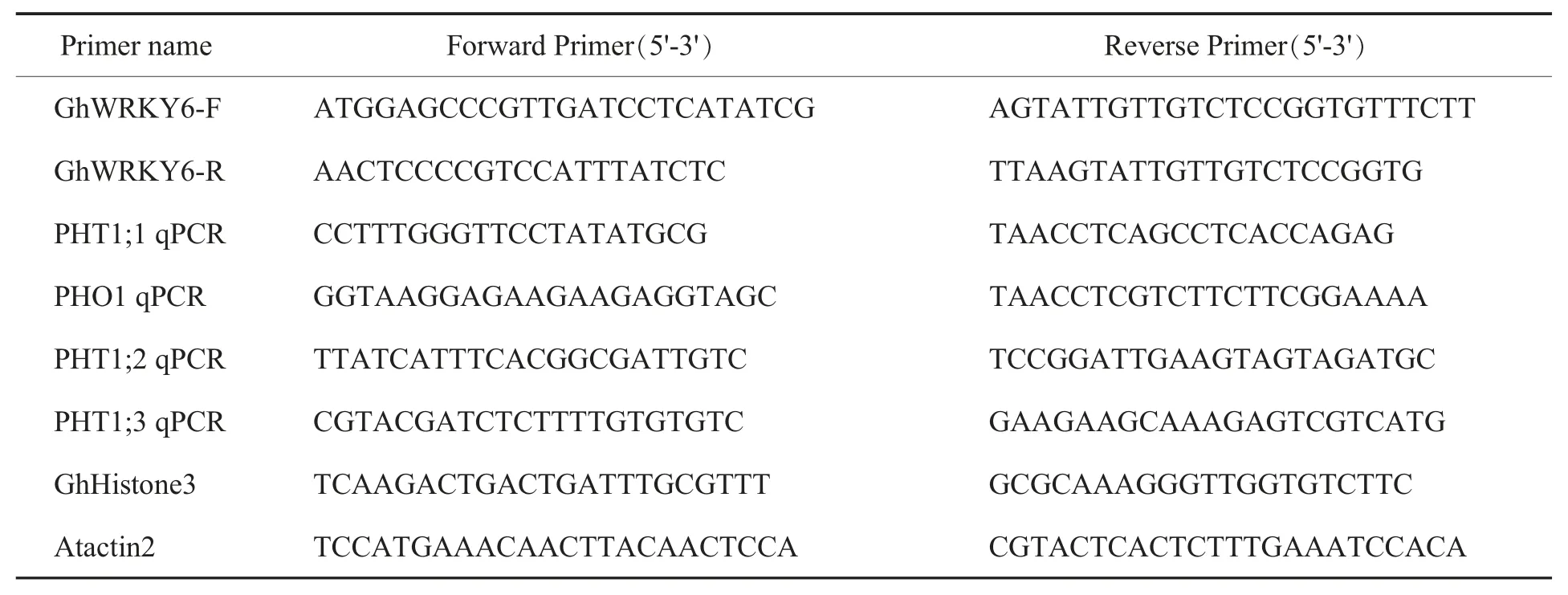
表1 本研究中使用的引物Table 1 Primers used in this study
2 結果與分析
2.1 GhWRKY6基因的克隆及特征分析
對GhWRKY6 基因的開放閱讀框(Open reading frame,ORF)全長序列設計特異擴增引物,以非生物脅迫處理后的陸地棉中棉所24 cDNA 為模板,通過TA 克隆并測序驗證得到正確的Gh-WRKY6 基因序列 (圖1-A)。該基因全長 1 575 bp,編碼525個氨基酸 。GhWRKY6(Gh_A01G0639 ) 定位于A01號染色體的1 1509 602-1 1511 977 bp (base pair)位置,包含有6個外顯子(圖1-B)。利用在線網站 CDD 對蛋白的保守結構域進行預測,結果顯示GhWRKY6 有1個典型的 WRKY 結構域(281~341AA),表明其是 WRKY 家族成員(圖1-C)。
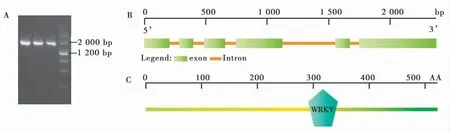
圖1 GhWRKY6 基因的克隆及特征分析Fig.1 Cloning and characterization of GhWRKY6 gene
2.2 GhWRKY6及其編碼蛋白的生物信息學分析
在 EXPASY 網 站上的 ProtParam 對Gh-WRKY6 基因編碼蛋白進行理化性質分析,結果表明,該蛋白的理論相對分子質量為57.629 kD,理論等電點為6.95,分子式為C2444H3919N761O804S25,脂肪酸系數為66.15,平均親水性為-0.772,不穩定系數為41.0,根據Guruprasad 等方法的計算結果表明,該蛋白為不穩定蛋白。利用SignalP4.0 Server 人工神經網絡算法對GhWRKY6 基因編碼蛋白進行信號肽預測,結果表明該蛋白的N 端至第70 位氨基酸之間剪切位點分值(C-Value)和綜合剪切分值(Y-Value)無明顯峰值,說明該蛋白的N 端不存在剪切位點,不包含信號肽,推測其為非分泌蛋白。利用TMHMM2.0 SERVER對GhWRKY6 基因編碼蛋白進行跨膜結構預測,在1~525個氨基酸之間沒有峰值,表明不存在跨膜蛋白,因此推測其不具有跨膜結構,是細胞膜外蛋白。
將GhWRKY6在NCBI中行進BLASTP比對,在氨基酸水平上發現與已報道的哥倫比亞錦葵 (Herrania umbratical,XP_021287397.1)、可可(Theobroma cacao,XP_007020766.2)、擬南芥(Arabidopsis thaliana,NP_564792.1)、煙草(Nicotiana attenuata,XP_0129249656.1)、葡萄 (Vitis vinifera,XP_002269696.2)和毛果楊(Populus trichocarppe,XP_002321134.3) 的相似性分別為73%、72%、50%、52%、51%和51%。序列比對發現GhWRKY6 基因編碼的氨基酸序列與以上物種存在高度保守區域(圖2-A)。用MEGA6 軟件繪制系統進化樹,結果表明GhWRKY6 蛋白與可可、哥倫比亞錦葵和擬南芥中的基因存在同源性(圖2-B)。
2.3 GhWRKY6的組織表達模式
提取陸地棉中棉所24 的根、莖、葉、花瓣(開花當天)、胚珠(開花當天)和纖維(開花后5 d、開花后10 d、開花后15 d)組織的RNA 進行組織特異性表達模式分析。結果發現該基因在根、莖和葉中均有表達,其中莖中表達量最高,是根中表達量的約5.5 倍;但是在花瓣、胚珠和纖維中表達量極低,尤其是在胚珠中幾乎不表達(圖3)。由此可知GhWRKY6 在棉花中有很明顯的組織特異性表達。
2.4 轉基因擬南芥的獲得
野生型擬南芥通過蘸花法[14]進行轉化后收獲種子,使用含有卡那霉素的MS 培養基篩選得到抗性苗。使用GhWRKY6 基因特異引物(表1)對篩選出的抗性苗進行PCR 擴增,以去除假陽性苗。結果有34 株擬南芥抗性苗可以擴增出與陽性對照(質粒)一致的特異性條帶(圖4-B)。
為了得到單拷貝的純合高代轉基因擬南芥,我們需要對陽性苗進行逐步篩選。經PCR 檢測獲得最終確定的陽性苗為T1代陽性苗,收獲種子并用含有卡那霉素的MS 培養基篩選,選擇后代中抗性性狀與非抗性性狀比值符合3:1 的株系,即T2代陽性苗。繼續收獲T2代陽性苗的種子再經過含有卡那霉素的MS 培養基篩選,選擇后代全部為抗性性狀的株系,即T3代純合陽性苗(圖4-A)。后續試驗選擇基因表達量較高的兩個純合轉基因株系OE3 和OE6 進行。
2.5 轉基因擬南芥表現出對低磷脅迫的超敏感表型
我們通過GhWRKY6 在擬南芥中的過表達來進一步明確GhWRKY6 在低磷脅迫中的功能。將野生型擬南芥、GhWRKY6 轉基因擬南芥和wrky6 突變體擬南芥種子在正常磷濃度的MS 培養基和低磷濃度(LP)的MS 培養基中生長1 周后發現,所有擬南芥種子在正常的MS 培養基上萌發和萌發后生長均無明顯差異,植物長勢正常;但在低磷MS 培養基中兩個轉基因株系OE3和OE6 表現出比野生型更明顯的受脅迫,種子萌發速率下降,子葉黃化,而此條件下的wrky6 突變體植株則表現出比野生型較強的抗性,植株的長勢稍強于野生型擬南芥(圖5-A)。
低磷脅迫會影響植物地上和地下部分的生長。我們將正常MS 培養基上生長1 周的野生型擬南芥、GhWRKY6 轉基因擬南芥和wrky6 突變體擬南芥轉移到低磷 (LP)MS 培養基上繼續生長,同時設置在正常磷含量的MS 培養基上生長的擬南芥為對照,結果觀察到當Pi 充足時,所有株系的擬南芥根部和葉片生長無明顯差異;但是當Pi 缺乏時,轉基因擬南芥主根生長受到抑制,側根數目減少。因此,在低磷脅迫下,GhWRKY6轉基因擬南芥與野生型擬南芥相比,無論是地上部分還是地下部分,其生長發育均受到抑制,抗逆性減弱,而wrky6 突變體的種子萌發和生長均強于野生型。
2.6 GhWRKY6影響磷信號通路相關基因的表達
為了進一步研究GhWRKY6 在低磷脅迫信號通路中的作用,我們對WT、GhWRKY6 轉基因擬南芥和wrky6 突變體在正常和低磷處理條件下Pi 轉運通路中一些重要Marker 基因的表達量進行檢測。有研究表明,磷酸鹽轉運體(Phosphate transporter,PHT) 家族基因和 SPX-EXS 亞家族PHO1 基因是維持植物體內Pi 穩態的關鍵基因[15]。因此我們重點關注了這兩類基因在野生型、wrky6 突變體和轉基因擬南芥中表達量的變化。結果表明: 無論是在Pi 充足還是缺乏的條件下,PHT1;1和PHO1 基因的表達量在wrky6 突變體中均明顯上調。在轉基因株系中,當Pi 充足時,PHO1 基因的表達量下降,PHT1;1 基因的表達量上升; 當Pi 缺乏時,PHO1 基因的表達量上升,PHT1;1 基因的表達被抑制。根據以上結果我們推測GhWRKY6 在磷元素的轉運調控中起到作用,且對PHT1;1 和PHO1 基因的調控依賴于磷含量的變化。
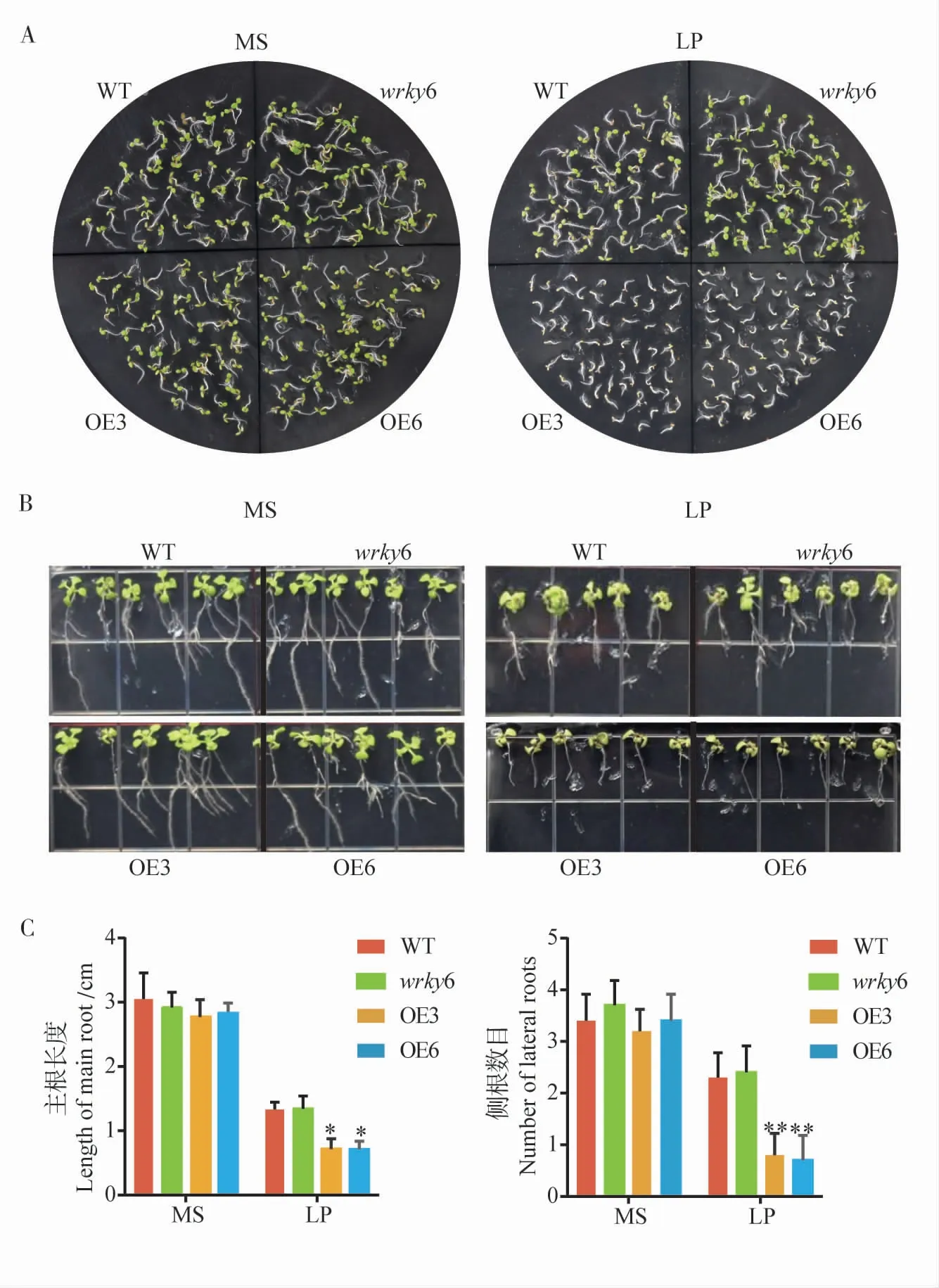
圖5 低磷脅迫對擬南芥的影響Fig.5 The effect of low phosphorus stress on Arabidopsis
當 Pi 含量充足時,PHT 家族中的PHT1;2和PHT1;3 的表達量在所有擬南芥株系中差異不明顯,但是在低磷(LP)處理條件下PHT1;3 表達量升高,PHT1;2 表達量變化不明顯,這可能是低Pi 處理誘導了PHT1;3 的表達。通過對低磷脅迫信號通路中Marker 基因表達量的變化,我們推測GhWRKY6 可能是通過調控磷轉運相關基因的表達影響植物體內Pi 的穩態,從而響應低磷脅迫。
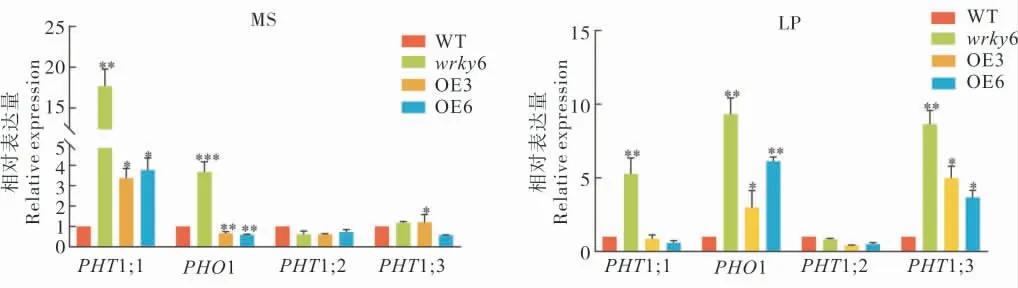
圖6 qRT-PCR 分析基因的表達量Fig.6 Analysis of the gene expression by qRT-PCR
3 討論
WRKY 轉錄因子是植物中最重要的轉錄因子家族之一。WRKY 轉錄因子家族最顯著的特征是家族成員至少包含一個由60個氨基酸殘基構成的WRKY 結構域。根據WRKY 結構域的數量及鋅指結構的特征,WRKY 轉錄因子家族可以被分為3 大組:Ⅰ組通常含有2個WRKY 結構域,位于C 端的WRKY 結構域具有DNA 結合活性,N 端的WRKY 域不能單獨與DNA 結合,該組鋅指結構是C2H2; Ⅱ組和Ⅲ組都含有一個WRKY結構域,但不同的是Ⅱ組鋅指結構是C2H2,而Ⅲ組鋅指結構是C2HC[25]。Dou 等利用基因組測序鑒定出了102個陸地棉WRKY 基因,并發現非生物脅迫下表達量明顯升高的大多是Ⅱ組和Ⅲ組的WRKY 基因[26]。本研究從陸地棉中克隆到一個WRKY 家族轉錄因子GhWRKY6,其屬于WRKY 轉錄因子家族的Ⅱb 亞組。GhWRKY6 與擬南芥AtWRKY6 的蛋白序列相似性最高為50%,故兩者可能存在相似功能。雖然有文獻報道了AtWRKY6 能夠影響種子萌發[16],增強擬南芥對病原體的抗性,加速擬南芥的早衰[17],參與磷的轉運調控[10],但對于陸地棉中GhWRKY6 的功能還尚未明確。因此我們通過構建過表達Gh-WRKY6 的轉基因擬南芥初步明確了GhWRKY6在植物響應低磷脅迫信號途徑中有重要功能。植物受到低磷脅迫時,主根的生長受到抑制,側根數量和長度會增加[18];地上部分生物量降低,葉片由于花青素積累顏色由綠變紫[19]。GhWRKY6 轉基因擬南芥在低磷脅迫條件下比野生型擬南芥表現出更為明顯的受脅迫,而wrky6 突變體在長勢上強于野生型。因此,GhWRKY6 可能作為一個負調控因子參與到植物響應低磷脅迫信號通路中。相對于干旱、高鹽、冷害和生物脅迫,WRKY 基因參與磷脅迫途徑的報道相對較少,尤其是在棉花中。海島棉GbWRKY1 基因是最近幾年報道的參與Pi 轉運調控的棉花WRKY 基因[20],而有關陸地棉WRKY 基因的類似報道目前并未出現,因此本研究結果為棉花WRKY 基因的功能研究提供了新的依據。
植物從外界吸收磷元素主要依賴于磷酸轉運蛋白,通過磷酸轉運蛋白將外界無機形式的磷轉為能夠被植物吸收和利用的有效磷[21]。磷酸轉運蛋白可被分為PHT 和PHO 兩大類,PHO 家族基因主要作用于植物體內Pi 的運輸[22],PHT 家族基因主要作用于植物從外界對磷的獲取[23]。前人的研究表明擬南芥AtWRKY6 在Pi 充足條件下結合PHO1 啟動子的W-box 抑制PHO1 基因表達[10]。AtWRKY42 與AtWRKY6 在調控 Pi轉運信號途徑中存在部分功能冗余,兩者都可以抑制PHO1 的表達,但是PHT1;1 的表達受AtWRKY42 的激活,從而提高植物獲取Pi 的能力[9]。PHT1;1和PHO1 基因的表達在wrky6 突變體中上調,而在轉基因株系中,GhWRKY6 對PHT1;1 和PHO1 基因的調控依賴于磷含量的變化。因此,GhWRKY6 在棉花中是否存在相似的調控機制還需要進一步試驗證明。
4 結論
本文以陸地棉cDNA 為模板克隆到一個WRKY 轉錄因子,cDNA 序列全長 1 575 bp,編碼 525個氨基酸,定位于 A01 號染色體的11 509 602~115 119 77 bp 位置,包含有 6個外顯子,有 1個典型的 WRKY 結構域 (281~341 AA),命名為GhWRKY6。組織表達模式分析表明該基因在棉花的根、莖、葉組織中表達量較高,在其他組織不表達或表達量極低,顯示出較明顯的組織特異性表達模式。過表達GhWRKY6 轉基因擬南芥與野生型擬南芥相比在低磷脅迫下主根生長受到抑制,側根減少,生物量減少,表現為磷脅迫敏感表型。磷轉運相關基因的qRT-PCR分析表明,GhWRKY6 參與磷轉運蛋白基因的調控。上述結果表明GhWRKY6 作為負調控因子參與到棉花低磷脅迫響應信號途徑中。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
