雞腿菇多糖的液態發酵工藝優化及抗氧化活性
2019-06-11 07:41:14陳旭峰王佳麗賈瑞娟
中國釀造 2019年5期
陳旭峰,王佳麗,賈瑞娟,許 女*
(1.山西農業大學 實驗教學中心,山西 太谷 030801;2.山西農業大學 食品科學與工程學院,山西 太谷 030801)
雞腿菇(Coprinus comatus)屬真菌門擔子菌亞門層菌綱傘菌目鬼傘科鬼傘屬,肉質細嫩、鮮美可口,研究表明,其具有降血糖、降血脂、增強免疫和抗腫瘤等多種生理功能,已被確定為符合聯合國糧農組織(food and agriculture organization,FAO)和世界衛生組織(world health organization,WHO)要求的具“天然、營養、保健”三種機能為一體的16種珍稀食用菌之一[1-3]。雞腿菇含有豐富的糖類,據測定,干品雞腿菇中總糖含量為57.65%,還原糖含量為53.54%,多糖含量為4.11%[4],是發揮生理功能的重要成分。關于雞腿菇多糖生理功能的研究主要集中在免疫[5-6]、降血糖[7]、抗病毒[8]和抗氧化[9]方面,多數研究都采用的是子實體提取的粗多糖。
應用深層發酵技術培養食用菌發展迅速,深層培養產生的菌絲體所含營養物質與子實體類似,深層發酵培養菌絲體生產周期短、產量高、成本低,液態深層發酵是開發雞腿菇活性成分的有效手段。金衛根等[10]采用正交試驗優化了雞腿菇菌絲體生長的最佳培養基和培養條件,研究發現,葡萄糖和黃豆為其最適的碳氮源,最佳培養條件為溫度25℃、裝液量75 mL/250 mL、搖瓶轉速150 r/min。李巍巍等[11]對雞腿菇產溶栓酶液體發酵條件進行了優化,最終得到的溶栓酶活力相當于尿激酶活力150 IU/mL,較優化前提高5倍。凡軍民[12]對雞腿菇菌絲體進行了深層發酵,并對其菌絲體多糖進行了提取、分離純化、結構鑒定和免疫活性研究。陸武祥等[13]比較了5種食用菌(猴頭菇、杏鮑菇、香菇、金針菇、雞腿菇)液體發酵菌絲多糖的抗氧化活性,結果表明,大部分菌絲多糖均具有一定的還原力及清除超氧陰離子自由基(O2-·)、羥自由基(·OH)和1,1-二苯基-2-三硝基苯肼(1,1-diphenyl-2-picrylhydrazyl,DPPH)自由基(DPPH·)的能力。
本研究在單因素試驗基礎上,通過響應面方法優化雞腿菇胞外多糖液態發酵工藝,并且探討發酵液上清的胞外多糖及菌絲體胞內多糖對DPPH·、羥基自由基(·OH)、一氧化氮自由基(NO·)、超氧陰離子自由基(O2-·)的清除能力及對由亞鐵離子(Fe2+)誘發的卵黃脂蛋白脂質過氧化抑制能力,評價其抗氧化活性,以期為雞腿菇液態發酵生物活性多糖產業化開發提供基礎研究。
1 材料與方法
1.1 材料與試劑
1.1.1 菌種
雞腿菇(Corprinus comatus)菌株SXAU-Chang:山西農業大學食用菌中心保藏、提供。
1.1.2 主要試劑
DPPH(純度95%):北京索萊寶科技有限公司;氯化硝基四氮唑藍(nitroblue tetrazolium,NBT)(純度95%):上海恒星應用化學研究所;核黃素(分析純):北京奧博星生物技術責任有限公司。
1.1.3 培養基
斜面種子培養基:土豆200 g,葡萄糖30 g,蛋白胨4 g,KH2PO42 g,MgSO4·7H2O 1 g,維生素B1(vitamin B1,VB1)0.01 g,水1 000 mL,pH自然。
液體發酵基礎培養基:葡萄糖40 g,蛋白胨4 g,KH2PO42 g,MgSO4·7H2O 1 g,VB10.01 g,水1 000 mL,pH自然。
1.2 儀器與設備
BS-2F恒溫振蕩培養箱:上海常思工貿有限公司;HZQF160全溫振蕩培養箱:哈爾濱市東聯電子有限公司;Vanti J-E型柜式冷凍離心機:美國Beckman Coulter公司;JA2003電子天平:上海精密科學儀器有限公司;DZKW-C型水浴鍋:河北省黃驊儀表廠;UV-9100型紫外可見分光光度計:北京瑞利分析儀器有限公司。
1.3 試驗方法
1.3.1 菌種活化
將保藏的雞腿菇菌種轉接于斜面培養基上,25℃培養至滿管,進行菌種活化,選擇菌絲生長旺盛的菌種備用。將斜面菌種切割成約0.5 cm2大小的菌絲塊,接種于裝有100 mL液體發酵基礎培養基的250 mL錐形瓶中,每瓶接半支斜面,25℃、160 r/min振蕩培養4 d。
1.3.2 搖瓶培養
將活化好的液體種子培養基,按10%(V/V)的接種量接種于液體發酵基礎培養基中,25℃、160 r/min振蕩培養6 d,測定菌絲體干質量及發酵液中的多糖含量。
1.3.3 雞腿菇菌絲體胞外和胞內多糖的制備
胞外多糖的制備:雞腿菇發酵液在8 000 r/min離心20min,將得到的上清液用旋轉蒸發儀濃縮至原體積的1/3,加入與原上清液等體積的體積分數為95%乙醇,于4℃條件下醇沉24 h,8 000 r/min離心15 min,取沉淀物于55℃烘箱中烘干至恒質量。
胞內多糖的制備:干菌絲體按照料水比1∶2(g∶mL)加水勻漿,在70℃條件下熱水抽提4 h,離心取上清液,濃縮至原體積的1/3,加入與原上清液等體積的體積分數為95%乙醇,于4℃條件下醇沉24 h,8 000 r/min離心15 min,取沉淀物于55℃烘干至恒質量。
1.3.4 分析檢測
采用干重法測定菌絲體生物量,將發酵液8 000 r/min離心20 min,菌絲體水洗兩次,60℃烘干至恒質量稱質量;采用苯酚-硫酸法測定雞腿菇胞外和胞內多糖含量。
1.3.5 雞腿菇多糖液態發酵工藝優化
(1)單因素試驗
在液體發酵基礎培養基的基礎上,分別替換其中的碳氮源,考查添加量均為4%的不同碳源(玉米粉、紅薯淀粉、可溶性淀粉、蔗糖、麥芽糖、葡萄糖),添加量均為0.4%的不同氮源(黃豆餅粉、花生餅粉、酵母膏、蛋白胨、硫酸銨、硝酸銨)對菌絲體生物量及發酵液中胞外多糖含量的影響。
(2)響應面試驗
選取玉米粉(X1)、蔗糖(X2)、黃豆餅粉(X3)、酵母膏(X4)作為主要的影響因素,以菌絲體生物量(Y1)和胞外多糖含量(Y2)為響應值,采用Box-Behnken設計,對雞腿菇液態發酵培養基進行優化,并進行響應面分析(response surface analysis,RSD),篩選最佳的發酵培養基條件。Box-Behnken試驗設計因素與水平見表1。

表1 發酵培養基配方優化Box-Behnken試驗因素與水平Table 1 Factors and levels of Box-Behnken design for fermentation medium formula optimization
(3)發酵條件優化
采用優化后的培養基,25℃條件下發酵6d,考察不同裝液量(50mL/250mL、70mL/250mL、90mL/250mL、120mL/250 mL)和轉速(120 r/min、160 r/min、180 r/min、200 r/min)對菌絲體干質量及發酵液中的胞外多糖含量影響。
1.3.6 抗氧化活性測定
對雞腿菇液態發酵產生的胞外多糖和胞內多糖進行DPPH·、·OH、O2-·、NO·清除能力及還原力和脂質過氧化抑制能力測定[14-15]。
2 結果與分析
2.1 雞腿菇多糖發酵培養基的優化
2.1.1 不同碳源對雞腿菇胞外多糖和菌絲體生成量的影響
碳源是菌體生長和多糖合成的物質基礎,適宜的碳源能使胞外多糖的產量得到提高,其濃度的大小也對胞外多糖產量產生影響,過高或過低都不利于胞外多糖的產生,甚至會抑制胞外多糖的合成。不同碳源對雞腿菇的生物量及胞外多糖含量的影響結果見圖1。由圖1可知,雞腿菇對碳源的利用相當廣泛,對單糖、寡糖、多糖都可以利用,其中以玉米粉和葡萄糖作為碳源時,生物量和胞外多糖含量較高,生物量分別為1.8 g/100 mL、2.0 g/100 mL,胞外多糖含量分別為1.5 g/L、1.7 g/L,考慮到葡萄糖成本高,所以綜合培養基的價格、來源、成本等因素,選擇玉米粉和蔗糖作為最優碳源。
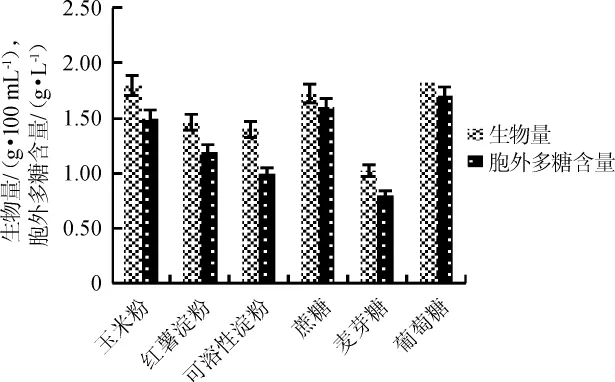
圖1 不同碳源對雞腿菇生物量及胞外多糖含量的影響Fig.1 Effect of different carbon sources on the biomass and extracellular polysaccharide of Coprinus comatus
2.1.2 不同氮源對雞腿菇多糖和菌絲體生成量的影響
氮源是食用菌細胞合成蛋白質、核酸必不可少的主要原料,在深層發酵過程中起著非常重要的作用。不同氮源對雞腿菇的生物量及胞外多糖含量的影響結果見圖2。由圖2可知,雞腿菇對有機氮的利用率高于無機氮,酵母膏、黃豆餅粉和花生餅粉作為氮源時,雞腿菇的生物量和胞外多糖含量較高,這與金萍[16]報道的結果相似,即酵母膏作為氮源時,菌絲體的生物量和多糖合成量最高,分別為2.2 g/100 mL和2.1 g/L。一方面這些氮源中含有豐富的蛋白質及多種生長因子,菌絲生長較快,胞外多糖產量也較高,另一方面可能是由于菌體自身分泌的蛋白酶較適合于這類蛋白。結合成本考慮,選擇黃豆餅粉和酵母膏作為雞腿菇生長的最優氮源。

圖2 不同氮源對雞腿菇生物量及胞外多糖含量的影響Fig.2 Effect of different nitrogen sources on the biomass and extracellular polysaccharide of Coprinus comatus
2.1.3 雞腿菇最佳碳氮源的響應面優化[20]
在單因素試驗基礎上,選取玉米粉(X1)、蔗糖(X2)、黃豆餅粉(X3)、酵母膏(X4)作為主要的影響因素,以菌絲體生物量(Y1)和胞外多糖含量(Y2)為響應值,采用Box-Behnken試驗設計,對雞腿菇多糖碳氮源進行優化,響應面試驗結果見表2,方差分析結果見表3,由RSA軟件分析可以得到響應面見圖3。

表2 發酵培養基配方優化Box-Behnken試驗設計及結果Table 2 Design and results of Box-Behnken experiments for fermentation medium formula optimization

表3 回歸模型方差分析Table 3 Variance analysis of regression model
由表3可知,模型在α=0.01水平上回歸顯著;失擬項反映的是實驗數據與模型是否相符的情況,失擬項P值=0.941 508>0.1,不顯著,因此,模型選擇正確。同時,一次項和平方項影響顯著(P<0.05),而交互項的影響不顯著(P>0.05)。復相關系數R2=0.9326,說明模型可以解釋93.26%試驗所得菌絲體生物量的變化,表明方程擬合較好,即回歸方程給雞腿菇培養基優化提供了一個合適模型。變異系數(coefficientof variation,CV)5.15%<10%,表明模型準確度高,變異可能性低。最終經軟件分析可得最優水平值為玉米粉3%、蔗糖3%、黃豆餅粉0.2%、酵母膏0.2%。在此最佳培養基條件下,菌絲體生物量為2.7 g/100 mL,胞外多糖產量為2.7 g/L。

圖3 玉米粉、蔗糖、黃豆餅粉及酵母膏添加量交互作用對菌絲體生物量影響的響應面及等高線Fig.3 Response surface plots and contour line of effects of interaction between corn flour,sucrose,soybean powder and yeast extract addition on mycelium biomass
2.2 雞腿菇多糖發酵條件優化
雞腿菇作為一種好氣性真菌,通氣量大,生長好,進而影響其代謝物胞外多糖的產生。本試驗采用優化后的培養基,考察不同的搖瓶裝液量和轉速對菌株發酵的影響,試驗結果見圖4和圖5。
由圖4可知,裝液量為50~90mL/250mL時,雞腿菇的生物量和胞外多糖產量逐漸增加;裝液量為90mL/250mL時,雞腿菇的生物量和胞外多糖產量最高,分別為2.9 g/100 mL和2.8g/L;裝液量為90~120mL/250mL時,雞腿菇的生物量和胞外多糖產量逐漸降低。因此,最適裝液量為90mL/250mL。
由圖5可知,轉速為120~160 r/min時,雞腿菇的生物量和胞外多糖產量逐漸增加;轉速為160 r/min時,雞腿菇的生物量和胞外多糖產量最高,分別為3.0g/100mL和2.8g/L;轉速為160~200 r/min時,雞腿菇的生物量和胞外多糖產量逐漸降低。因此,最適轉速為160 r/min。

圖4 裝液量對雞腿菇生物量及胞外多糖含量的影響Fig.4 Effect of liquid loading volume on the biomass and extracellular polysaccharide of Coprinus comatus

圖5 轉速對雞腿菇生物量及胞外多糖含量的影響Fig.5 Effect of rotation speed on the biomass and extracellular polysaccharide of Coprinus comatus
2.3 雞腿菇多糖發酵曲線測定
采用上述優化的發酵工藝,10%接種量,12層紗布包扎瓶口,25℃條件下培養10d。每天測定菌絲體生物量和發酵液中的胞外多糖含量,繪制雞腿菇多糖發酵曲線見圖6。

圖6 雞腿菇菌絲干質量及胞外多糖含量搖瓶發酵曲線Fig.6 Fermentation curve of mycelium dry mass and extracellular polysaccharide of Coprinus comatus in shaking flask
由圖6可知,菌絲體生物量在第7天達到最大值,為3.2 g/100 mL,之后由于發酵液中營養物質減少,菌絲生長速度變慢,部分菌絲開始自溶,所以整體表現為菌絲干質量下降,發酵液顏色也逐漸變深。在發酵第6天,雞腿菇產生的胞外多糖含量達到最大值,為2.9 g/L,之后胞外多糖的含量開始下降,最終趨于平緩。分析可能是因為隨著發酵的進行,營養物質逐漸變得不充足,菌體分泌出的胞外多糖又被自身重新利用,所以胞外多糖含量在后期開始下降。
2.4 雞腿菇胞外和胞內多糖抗氧化活性測定
2.4.1 總還原力測定
一般情況下,物質的還原能力越強,其抗氧化活性也越高。雞腿菇胞內及胞外多糖的還原力測定結果圖7。由圖7可知,雞腿菇的胞外多糖和胞內多糖溶液均具有一定的還原能力,但胞內多糖的還原能力高于胞外多糖,在質量濃度為8mg/mL時,胞外及胞內多糖的還原力分別達到0.52和0.78,但仍顯著低于VC的還原力。
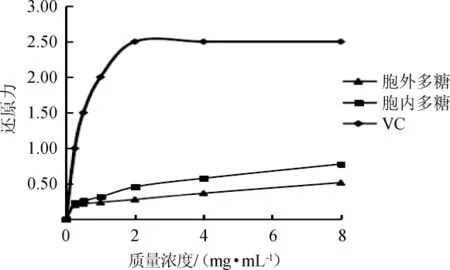
圖7 雞腿菇胞外和胞內粗多糖的還原力Fig.7 Reducing power of extracellular and intracellular crude polysaccharides from Coprinus comatus
2.4.2 DPPH·清除能力測定
由圖8可知,雞腿菇胞外和胞內多糖清除DPPH·的能力隨濃度的增加而增強,具有明顯的濃度依賴性。在低濃度下就有一定的還原力,當質量濃度為0.25 mg/mL,胞外及胞內多糖對DPPH·的清除率分別達到15.9%和20.1%;當質量濃度升高至2.00 mg/mL時,胞外多糖對DPPH·的清除率達到47.4%;胞內多糖對DPPH·的清除能力則達到55.1%,但二者均低于VC的DPPH·清除能力。

圖8 雞腿菇胞外和胞內粗多糖對DPPH自由基清除能力測定Fig.8 Scavenging capacity of extracellular and intracellular crude polysaccharides from Coprinus comatus on DPPH·
2.4.3 ·OH清除能力測定
羥基自由基是活性氧類物質中化學活性最強的物質。在細胞內,羥基自由基很容易通過特定位置穿過細胞膜,然后和氨基酸、蛋白質和脫氧核糖核酸(deoxyribonucleic acid,DNA)等生物分子發生反應,造成組織破壞和細胞死亡。因此,去除羥基自由基可能是生物體預防多種疾病最有效的措施之一[17-18]。雞腿菇胞外、胞內粗多糖對羥基自由基的捕獲能力結果見圖9。
由圖9可知,雞腿菇多糖對·OH清除作用與濃度有著較強的依賴關系。當胞外多糖質量濃度為8 mg/mL時,清除率達44.1%;胞內多糖對·OH清除的清除率達52.2%,遠遠高于吳艷兵[8]報道的同等濃度下經脫蛋白處理的雞腿菇子實體多糖的·OH清除率(25%);然而卻低于王金璽等[20]的報道(80%)。

圖9 雞腿菇胞外和胞內粗多糖對·OH清除能力測定Fig.9 Scavenging capacity of extracellular and intracellular crude polysaccharides from Coprinus comatus on·OH
2.4.4 O2-·清除能力測定
由圖10可知,雞腿菇胞外和胞內多糖具有一定的O2-·清除能力,在8 mg/mL的濃度時,二者的清除率分別為27.2%和38.2%,與王金璽等[20]報道的同等濃度下雞腿菇粗多糖的結果相當(26%),但二者都遠遠低于VC的O2-·清除能力。雞腿菇粗多糖表現出較弱的清除O2-·的能力,其原因可能是與其空間構象相關,使其羥基被包裹在結構內部,無法與結構外圍的O2-·發生作用[20]。

圖10 雞腿菇胞外和胞內粗多糖對O2-·清除能力測定Fig.10 Scavenging capacity of extracellular and intracellular crude polysaccharides from Coprinus comatus on O2-·
2.4.5 NO·的抑制能力測定
一氧化氮自由基(NO·)已被廣泛認同在各種生理和病理過程中發揮調節作用。但是,過多產生的一氧化氮自由基(NO·)能夠引發各種不利于健康的細胞反應,并且造成包括炎癥、膿腫、中風、動脈硬化在內的多種疾病。一氧化氮自由基(NO·)捕獲劑可以中止由于過多硝基自由基引發的鏈式反應。雞腿菇胞外、胞內多糖和VC對NO·的抑制作用結果見圖11。

圖11 雞腿菇胞外和胞內粗多糖對NO·的抑制能力Fig.11 Inhibiting capacity of extracellular and intracellular crude polysaccharides from Coprinus comatus on NO·
由圖11可知,在試驗濃度范圍內,兩種多糖組分的抑制作用雖然比維生素C要弱,但也具有明顯的效果。胞外多糖、胞內多糖在8mg/mL時,NO·清除率分別達48.1%、65.1%。
2.4.6 抑制脂質過氧化能力測定
生物體內的脂質過氧化主要發生在生物膜上,由于構成生物膜骨架的主要成分磷脂中含有多不飽和脂肪酸,最容易發生過氧化反應,而脂質過氧化和細胞損傷的發生,可引起人體衰老和心臟病、動脈粥樣硬化、癌癥、炎癥、糖尿病等嚴重疾病。引起機體脂質過氧化的因素很多,其中二價鐵離子是生物體內最主要的脂質過氧化促進劑[21],故本研究以二價鐵離子為氧化促進劑,以卵黃脂質體體系為模型,研究雞腿菇多糖成分對脂質過氧化的抑制作用。卵黃中磷脂C-2位上所含的極低密度脂蛋白(very low density lipoprotein,VLDL)和低密度脂蛋白(low densitylipoprotein,LDL)及過不飽和脂肪酸(polyunsaturated fattyacids,PUFA)在鐵離子的催化下,經振蕩,能誘發過氧化,產生烷氧基(LO·)和烷過氧基(LOO·)物質,再引發鏈式反應。當有抗氧化劑存在時,可清除過氧化產物,從而阻斷鏈式反應的發生,使得該卵黃脂蛋白脂質過氧化體系的過氧化產物生成受阻,根據吸光度數值的變化可測定抗氧化劑的抑制率[21]。

圖12 雞腿菇胞外和胞內粗多糖對卵黃脂蛋白過氧化的抑制能力Fig.12 Inhibiting capacity of extracellular and intracellular crude polysaccharides from Coprinus comatus on lipovitellinin peroxidation
雞腿菇胞外和胞內多糖對Fe2+誘發的卵黃脂蛋白脂質過氧化的抑制結果見圖12。由圖12可知,胞外和胞內對Fe2+誘發的脂質過氧化的抑制能力與濃度均呈正相關,兩者在試驗濃度最高濃度時的抑制率分別為25.2%和37.1%,抑制率作用均不明顯,遠低于VC抑制作用。
3 結論
通過響應面方法優化雞腿菇胞外多糖液態發酵工藝,并且對發酵液上清的胞外多糖及菌絲體胞內多糖的抗氧化活性進行系統研究,結果表明,最優發酵工藝為玉米粉3%、蔗糖3%,黃豆餅粉0.2%,酵母膏0.2%,裝液量90mL/250mL,轉速160 r/min,25℃條件下發酵6 d。在此最優工藝下,雞腿菇菌絲體生物量可達到3.2 g/100 mL,胞外多糖產量可達到2.9 g/L。抗氧化試驗結果表明,雞腿菇多糖對DPPH自由基的清除率很高,胞內多糖的清除能力強于發酵上清胞外多糖,并且對·OH和NO·也有著較好的清除效果,另外,雞腿菇胞內和胞外多糖還對O2-·及Fe2+誘發的卵黃脂蛋白脂質過氧化具有一定的清除和抑制能力。雞腿菇液態發酵多糖具有良好的的抗氧化活性,顯示了其作為功能食品添加劑的應用潛力。
