轉基因ABP9玉米株系的耐鹽性分析
2019-06-04 07:04:44張昭楊龐軍玲韓梅冷鵬飛趙軍
生物技術通報 2019年5期
關鍵詞:植物
張昭楊 龐軍玲 韓梅 冷鵬飛 趙軍
(1. 吉林農業大學中藥材學院,長春 130118;2. 中國農業科學院生物技術研究所,北京 100081)
土地鹽漬化嚴重制約農業生產,導致世界范圍內作物產量受損。據統計,全球有50%澆灌地和20%農業用地遭受不同程度鹽害[1]。我國鹽漬土分布廣泛,大約占全球鹽漬土面積的1/10,嚴重影響我國農業的發展[2]。玉米是世界三大糧食作物之一,在中國種植面積居世界第二位,在國民經濟中占有重要地位[3]。玉米是中度鹽敏感植物,耐鹽能力比較低,因此限制了玉米種植面積和產量[4]。隨著生物技術的不斷發展,采用轉基因技術培育抗逆轉基因玉米已經成為提高玉米抗逆性的有效途徑之一[5]。
植物中bZIP類轉錄因子參與多種生物過程和對干旱、鹽漬和極端溫度等逆境脅迫的反應[6-7]。中國農業科學院生物技術研究所課題組前期利用酵母單雜交方法從玉米中克隆了一個可與過氧化氫酶基因Cat1[8]上游順式作用元件ABRE2相互作用的bZIP類轉錄因子基因ABP9[9]。通過對諸多轉ABP9植物研究表明,該基因可以大幅度提高植物對鹽漬等多種非生物脅迫的耐性[10-14]。
ABP9過表達可以提高擬南芥、水稻等植物的抗逆性,但在玉米中的研究還知之甚少。為了改良玉米品種的耐鹽性和進一步研究ABP9過表達提高植物耐鹽性的調控機制,中國農業科學院生物技術研究所課題組創制了鄭58背景由逆境誘導型啟動子Pabp9[15]驅動ABP9表達的轉基因玉米株系。旨在通過研究其在NaCl脅迫下的生理和基因表達變化,分析其耐鹽性和初步探討其耐鹽分子調控機制,為后續研究工作提供依據。
1 材料與方法
1.1 材料
試驗材料為鄭58背景由逆境誘導型啟動子Pabp9驅動ABP9表達的轉基因玉米株系,對照為同一遺傳背景的轉基因陰性株系,種子均系中國農業科學院實驗室保藏。
1.2 方法
1.2.1 轉基因株系分子鑒定 采用盆栽法(蛭石∶營養土=1∶1)于人工氣候室內種植,溫度27℃(±3℃),光周期16 h/8 h,光照強度500-1 000μmol/m2·s1,相對濕度為40%-50%。三葉期取葉片保存于-80℃備用。
(1)基因組DNA提取及PCR檢測 玉米葉片基因組DNA提取使用新型植物基因組DNA提取試劑盒(天根生化科技有限公司)。
使用特異引物ABP9-Fw:5'-CGAAGAAAAAGAAGCTGGTGGAG-3',Tnos-Rv:5'-TGCGGGACTCTAATCATAAAAACC-3',對轉基因ABP9株系進行PCR檢測。
PCR反應程序為95℃ 5 min;95℃ 30 s,60℃40 s,72℃ 40 s,32個循環;72℃ 10 min。經電泳檢測PCR產物長度為582 bp的即為陽性植株。
(2)Southern blot分析 采用CTAB法大量提取玉米基因組DNA,探針合成及雜交試劑盒均購自羅氏。具體方法參照試劑盒說明及《分子克隆實驗指南·第三版》
(3)植物總RNA提取及qRT-PCR分析 采用RNAprep Pure植物總RNA提取試劑盒(天根生化科技有限公司)提取植物總RNA,參照TransScript One-Step gDNA Removal and cDNA Synthesis SuperMix試劑盒(北京全式金生物技術有限公司)反轉錄成cDNA,用7500熒光定量PCR儀檢測相關基因的表達,反應程序為95℃ 30 s;95℃ 15 s,60℃ 15 s,72℃ 30 s,40個循環;采用 2-ΔΔCt方法分析基因的表達水平。引物如下:
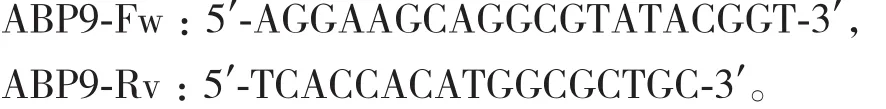
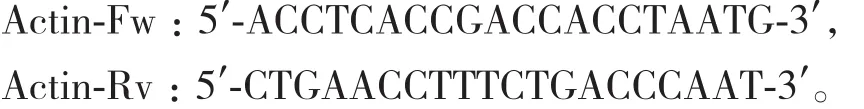
內源參照基因Actin引物為:
1.2.2 生理指標測定 挑選籽粒飽滿程度一致的種子用5%次氯酸鈉溶液消毒20 min,去離子水沖洗干凈,種于蛭石中。生長至一葉期時在溫室中用1/2Hoagland營養液進行水培,光周期、溫度及濕度與1.2相同。兩葉期時逐漸增加營養液濃度至全營養液培養,每天氣泵增氧2 h,每隔2 d更換一次營養液。三葉期開始鹽脅迫處理,為了防止鹽應激,每隔12 h增加50 mmol/L NaCl,直至終濃度150 mmol/L NaCl,對照組全營養液培養。處理7 d,期間觀察表型及測量各項生理指標。
(1)葉綠素含量測定 采用乙醇丙酮法測定葉綠素含量[16]。
(2)葉綠素熒光參數的測定 挑選長勢一致的幼苗,選取相同葉位功能葉,固定葉夾,避光20 min,使用便攜式植物熒光檢測儀(Handy PEA,Hansatech Instrument Ltd,UK)測PSII的最大光化學效率(The maximal efficiency of PSII photochemistry,Fv/Fm),測定光源為波長650 nm、光強3 000 μmol/m2·s1的紅光,熒光信號的記錄時長為2 s。計算公式為Fv/Fm=(Fm-Fo)/Fm,具體生理意義參照王立豐等的研究結果[17]。
(3)葉片相對含水量測定 三葉期水培苗直接用150 mmol/L NaCl處理12 h,恢復生長3 h后,剪取第二片全展葉1-2 cm稱取鮮重(Fresh Weight,FW),將葉片放入去離子水中浸泡至恒重(Total Weigh,TW)。再將葉片放入烘箱105℃殺青10 min,80℃烘干至恒重(Dry Weight,DW)。葉片相對含水量(%)=(FW-DW)/(TW-DW)×100%。
(4)游離脯氨酸及可溶性糖含量測定 脯氨酸含量測定參照脯氨酸(PRO)測試盒(南京建成生物工程研究所),可溶性糖測定參照植物可溶性糖含量檢測試劑盒提供的方法(北京索萊寶生物科技有限公司)。
(5)丙二醛(Malondialdehyde,MDA)含量測定 測定方法參照植物MDA試劑盒提供的方法(南京建成生物工程研究所)。
(6)相對電導率測定 三葉期水培苗用150 mmol/L NaCl處理3 d時,將幼苗葉片用超純水中沖洗3次,使用打孔器剪取葉片,置于裝有10 mL超純水的離心管中,抽真空10 min,讓葉片全部沉入管底,室溫靜置1 h,搖勻后測初始電導值(E1)。沸水浴10 min以殺死植物組織,流水冷卻至室溫,搖勻后次測定終電導值(E2)。相對電導率計算公式為相對電導率(RE)=E1 /E2。
(7)抗氧化酶活性測定 超氧化物歧化酶(Superoxide dismutase,SOD)、過氧化物酶(Peroxidase,POD)及過氧化氫酶(Catalase,CAT)活性測定參照試劑盒提供的方法(南京建成生物工程研究所)。粗酶液制備方法為稱取0.5 g葉片,加2 mL磷酸鹽緩沖液(0.1 mol/L,pH=7.4),冰水浴條件下加入石英砂研磨,離心取上清。
1.2.3 轉錄組測序 用150 mol/L NaCl處理三葉期玉米幼苗,取處理0 h、15 min、30 min、1 h、2 h、3 h、6 h、9 h和12 h的玉米葉片,液氮速凍后-80℃保存。用RNAprep Pure植物總RNA提取試劑盒(天根生化科技有限公司)提取RNA,由諾禾致源測序公司完成測序。利用Tophat2軟件分析測序后的序列信息,并且只保留比對到基因組上單一位點的序列進行后續分析。利用GenomicFeatures和GenomicAlignments軟件包統計比對到參考基因上的序列數目。然后用DESeq2軟件包進行樣本間的歸一化,并且計算基因的表達量(FPKM值)。樣本之間的差異基因也是利用DESeq2軟件包完成。
基因的GO富集分析利用在線分析軟件agriGO完成。每一個條目中允許的最少基因數為5,Fisher精確檢驗用來檢測輸入的基因列表與背景基因(AGPv3,Ensembl release 30)之間的差異。采用Benjamini-Yekutieli方法校正P值,最終取FDR <0.05作為最終顯著性檢驗的標準。對篩選的部分差異表達基因進行qRT-PCR驗證,方法同1.2.3,引物見表1。

表1 引物設計
2 結果
2.1 轉基因玉米植株分子鑒定
利用特異性引物進行PCR檢測,植株均能擴增出特異性條帶(圖1-A)。根據T-DNA片段所在區域,選用HindⅢ和EcoRI酶切后,進行southern blot分析,結果(圖1-C)顯示為單個拷貝插入。
利用qRT-PCR檢測ABP9在轉基因及非轉基因植株中的表達水平,轉基因植株中ABP9表達量高于陰性對照(圖1-B)。結果表明,ABP9以單拷貝形式整合到轉基因玉米基因組中,并得到較高水平表達。
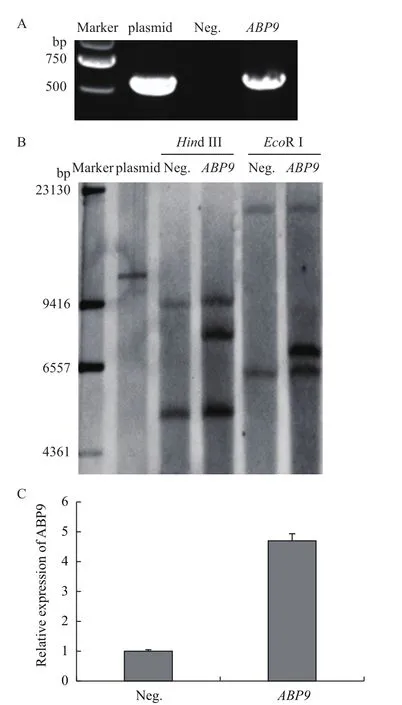
圖1 轉基因植株分子鑒定
2.2 表型觀察及葉綠素含量測定
鹽處理至第3 d時,陰性植株第一片真葉失綠枯萎,而轉基因植株仍能保持較鮮綠狀態(圖2-A)。分別測定處理3 d的第一片真葉(圖2-B)和處理7 d第二片全展葉(圖2-C)的葉綠素含量,結果顯示,與對照組相比,處理組葉綠素含量均有下降,但轉基因植株葉綠素含量均極顯著高于陰性植株(P<0.01)。
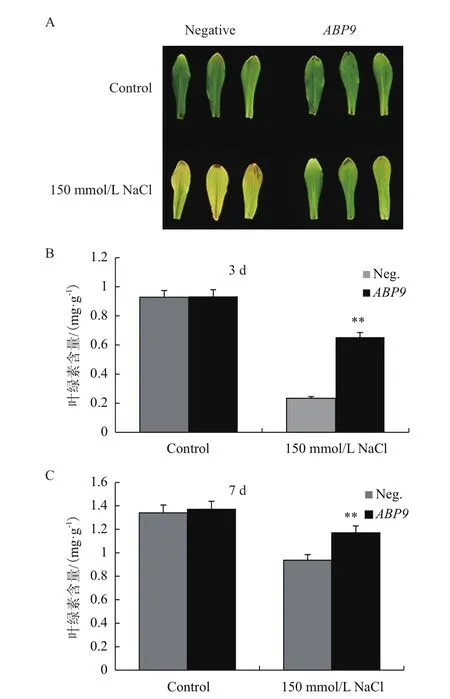
圖2 NaCl脅迫對玉米幼苗葉綠素含量的影響
2.3 NaCl脅迫對玉米幼苗Fv/Fm的影響
葉綠素熒光參數是植物耐鹽性分析中一個重要指標,測定NaCl脅迫下玉米幼苗PSII最大光能轉化效率(Fv/Fm)的變化。隨著脅迫時間延長,玉米幼苗Fv/Fm逐漸下降。當處理至第5天和第7天時,轉基因植株Fv/Fm顯著高于陰性植株(圖3)。
2.4 NaCl脅迫對玉米幼苗葉片含水量的影響
在150 mmol/L NaCl脅迫下,轉基因與陰性幼苗葉片逐漸失水萎蔫,但兩者間無顯著差異。恢復生長3 h后葉片含水量逐漸上升,但轉基因植株葉片含水量極顯著高于陰性植株(P<0.01)(圖4)。
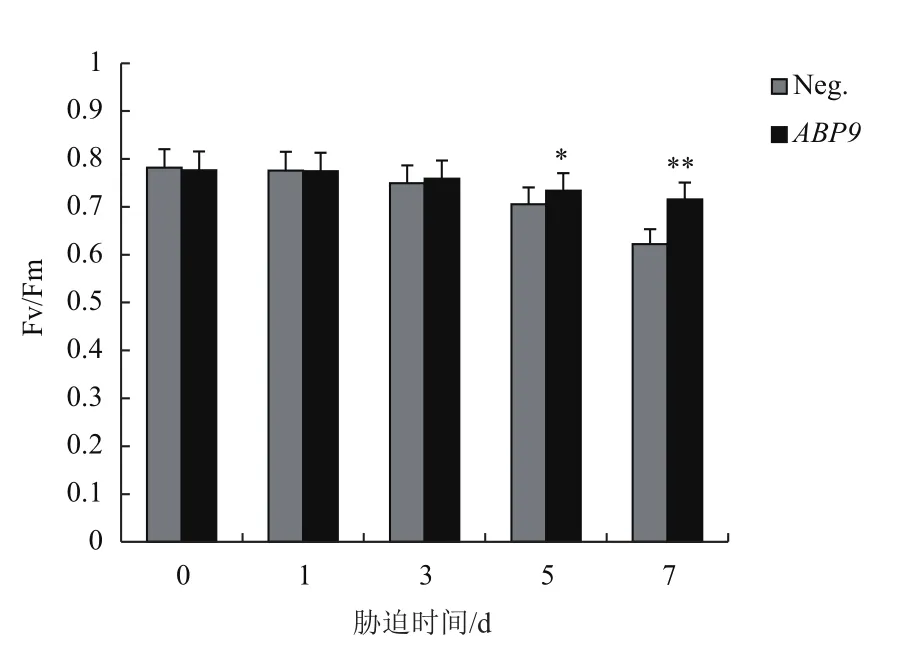
圖3 NaCl脅迫對玉米幼苗Fv/Fm的影響
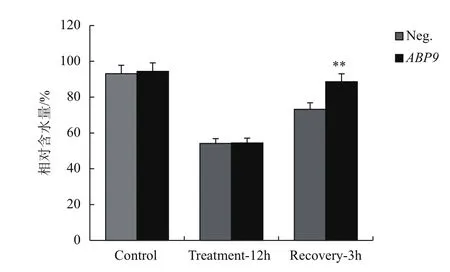
圖4 NaCl脅迫對玉米幼苗葉片相對含水量的影響
2.5 NaCl脅迫對玉米幼苗滲透調節物質含量的影響
鹽脅迫條件下,植物常通過有機物質的積累來維持較高的細胞質滲透壓。通過測定脯氨酸和可溶性糖含量的變化,發現兩種滲透調節物質隨處理時間增長,呈先上升后下降趨勢,但轉基因植株的脯氨酸(圖5-A)和可溶性糖(圖5-B)含量顯著高于陰性植株。
2.6 NaCl脅迫對玉米幼苗丙二醛(MDA)含量和電導率的影響
鹽脅迫導致植物細胞受到傷害發生膜脂過氧化作用,細胞膜透性增加,MDA含量和電導率上升。如圖6所示,隨脅迫時間增加,丙二醛含量呈先上升后下降趨勢,但陰性植株丙二醛含量要高于轉基因植株(圖6-A),在多個時間點達到顯著水平。處理至3 d時,陰性植株葉片相對電導率顯著高于轉基因植株(圖6-B)。
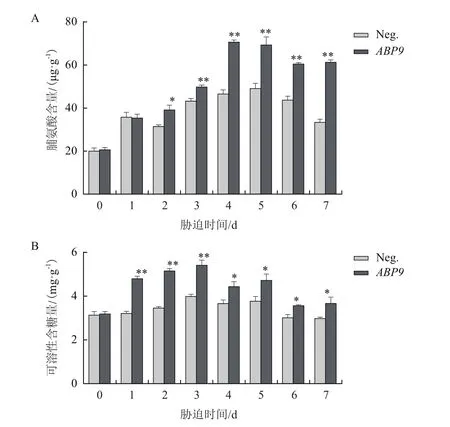
圖5 NaCl脅迫對玉米幼苗脯氨酸(A)和可溶性糖(B)含量的影響
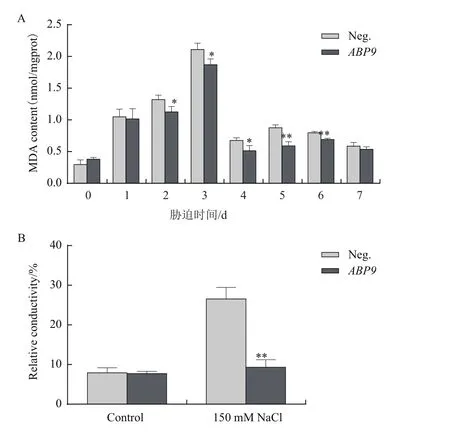
圖6 NaCl脅迫對玉米幼苗丙二醛含量(A)和電導率(B)的影響
2.7 NaCl脅迫對玉米幼苗抗氧化酶活性的影響
逆境可使植物體內產生大量活性氧,而SOD、POD和CAT是清除除活性氧的重要抗氧化酶,其活性上升是提高玉米抗逆能力的重要因素。如圖7所示,受到鹽脅迫后,轉基因與陰性植株葉片中的SOD、POD和CAT均呈先上升后下降的趨勢,但轉基因植株中抗氧化酶活性顯著高于陰性植株。
2.8 NaCl脅迫下玉米幼苗RNA-seq分析
對轉基因和陰性玉米的三葉期幼苗鹽脅迫處理0 h、15 min、30 min、1 h、2 h、3 h、6 h、9 h 和 12 h的葉片進行RNA-seq分析。首先比較各個時間點中轉基因和陰性玉米的基因表達差異,將各時間點的所有差異基因合并并去除重復基因,共得到2 139個差異表達基因,這些基因是受ABP9調控的基因。
同時,利用轉基因陰性植株的RNA-seq數據挖掘其中響應鹽脅迫的基因,將鹽處理各個時間點的轉錄組數據分別與0 h的數據進行比較,合并所有差異表達的基因之后共得到2 453個基因。將兩組差異基因作交集后得到1 075個基因,這些基因為受ABP9調控的且響應鹽脅迫的基因。將這些基因進行GO功能富集分析,發現這些基因顯著富集在響應非生物脅迫及活性氧清除等生物學過程中。
選取其中一些響應鹽脅迫上調表達且在轉基因和對照植株中表達差異倍數≥2的基因(表2)進行qTR-PCR驗證(圖8),結果顯示,與轉錄組測序結果基本一致,這些基因均是鹽誘導基因,且其脅迫過程中的表達在轉基因植株中相比陰性對照明顯提高。
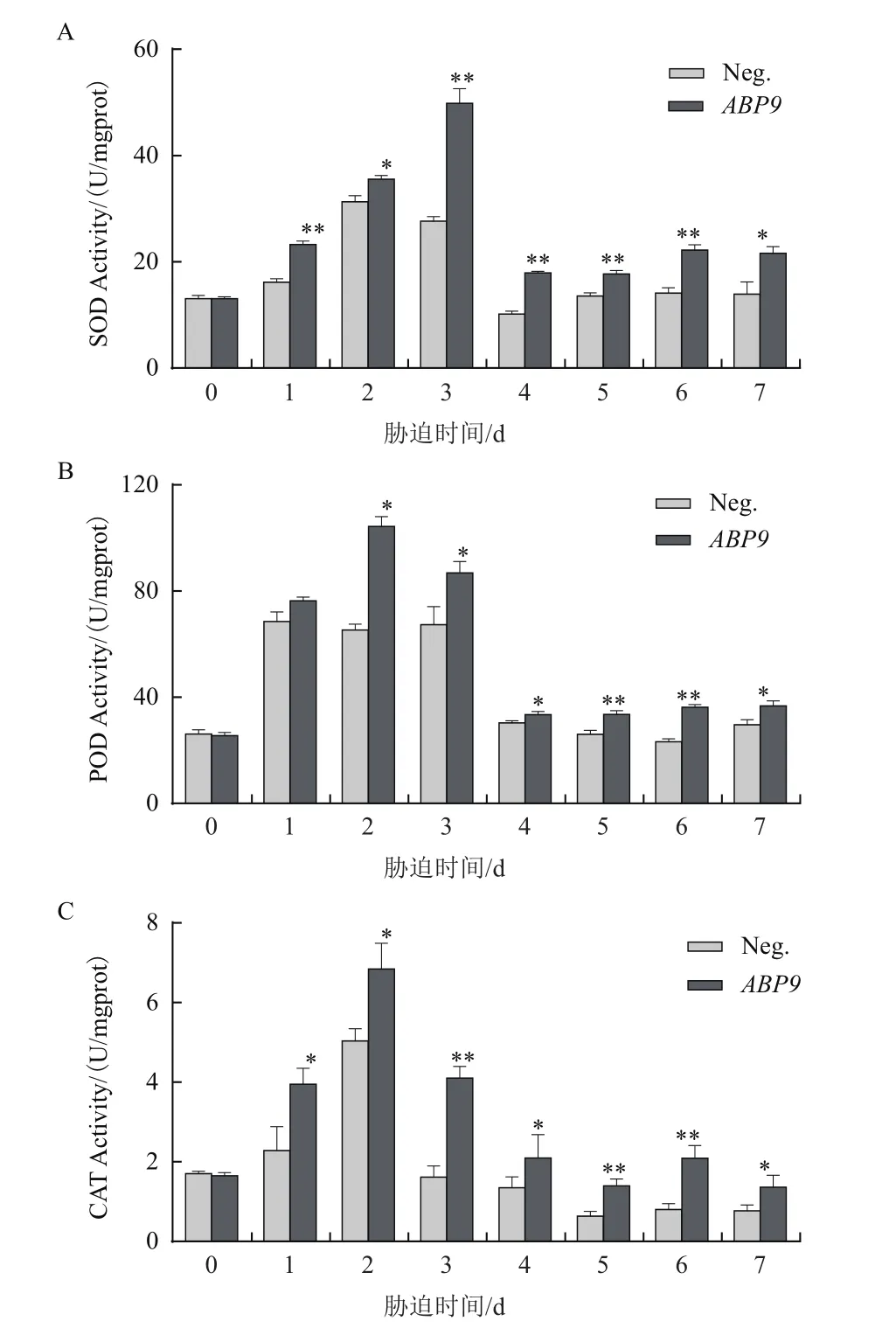
圖7 NaCl脅迫對玉米幼苗SOD(A)、POD(B)和CAT(C)活性的影響
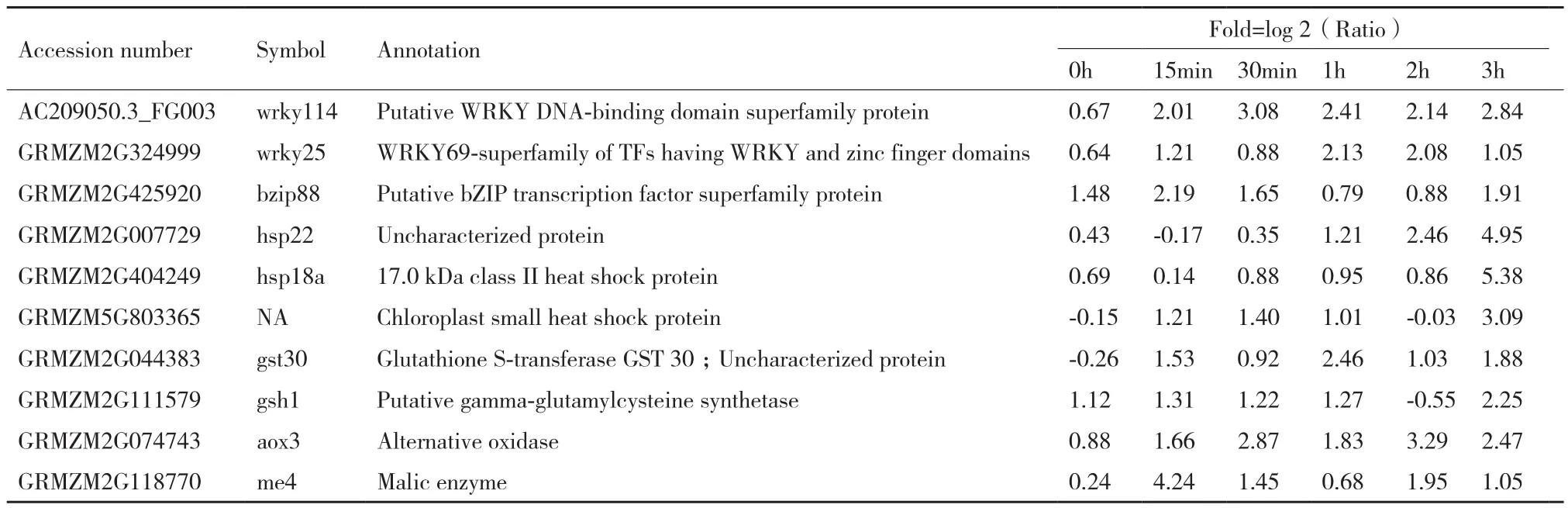
表2 部分差異表達基因列表
3 討論
近年來,隨著轉基因技術的不斷發展,應用該技術對農作物改良已成為培育具有優良性狀新品種的有效途徑[18]。已有諸多報道成功獲得轉基因植株,并在提高其耐鹽性等方面有顯著成效[19-20]。在前期工作的基礎上,創制了鄭58背景的轉ABP9玉米株系,通過PCR檢測和Southern blot分析證明ABP9已成功整合到玉米基因組中,且為單拷貝形式插入。qRT-PCR分析結果顯示ABP9在轉基因株系中表達量較高。
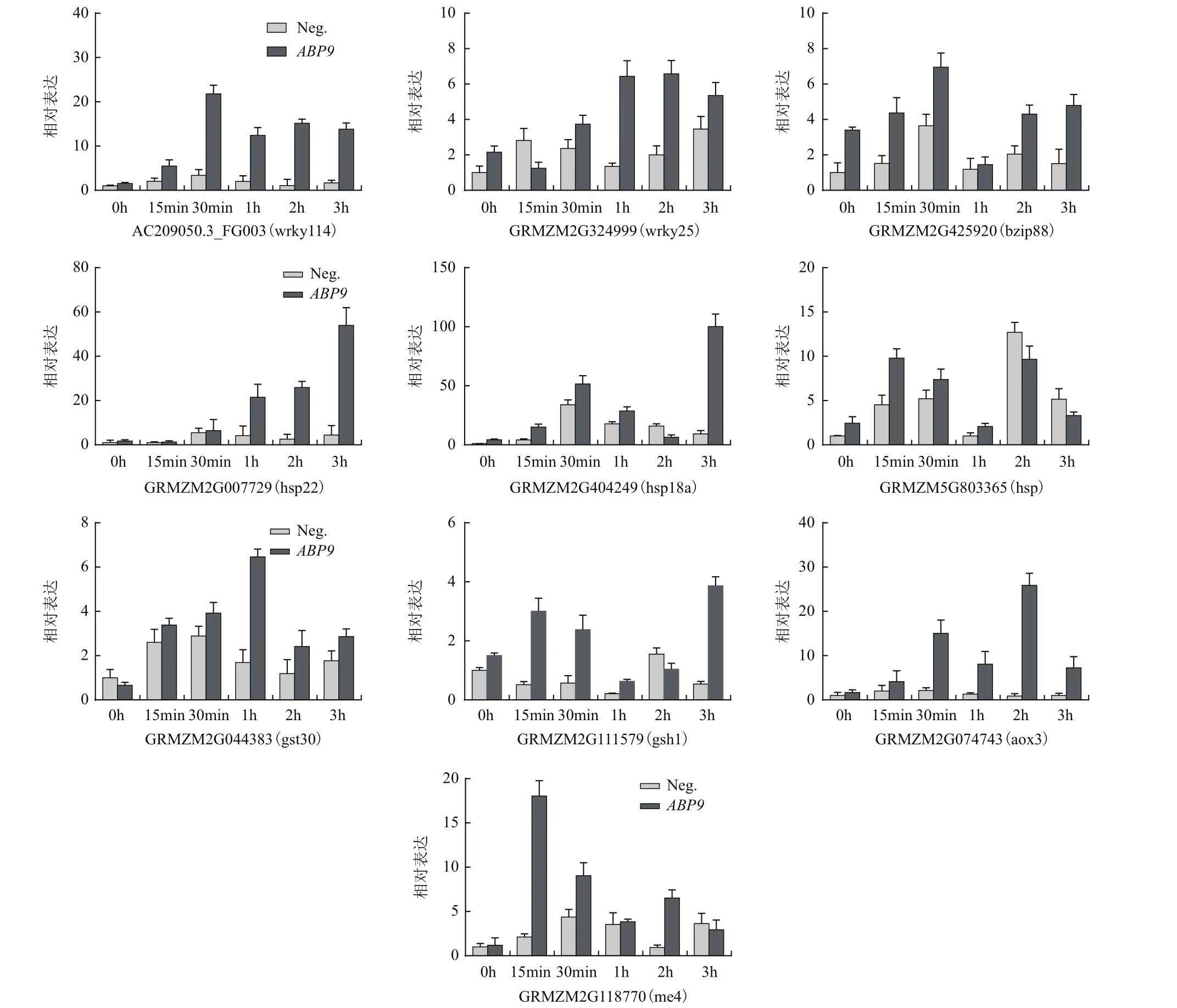
圖8 鹽脅迫下轉基因植株中差異表達基因qRT-PCR鑒定
當植物遭受鹽脅迫時,可在生理及分子水平快速響應,通過一系列的生理生化反應適應和抵御鹽害。葉綠素是對鹽脅迫敏感的光合色素,鹽脅迫會加速植物葉綠素降解,降低葉綠體對光的吸收,從而導致光合速率下降[21]。本研究結果顯示,鹽脅迫后轉基因和陰性對照玉米葉片的葉綠素含量下降,說明長時間高濃度的鹽脅迫導致葉綠素大量分解。但轉基因植株葉綠素含量顯著(P<0.01)高于陰性植株,說明前者具有較強的抵御葉綠素降解的能力,進而保持了較高的光合能力。通過葉綠素熒光參數的變化可以了解植物光系統對光能的吸收、耗散等一系列反應,能夠反映植物受逆境影響的情況[22]。研究表明,鹽脅迫會破壞PSII、抑制光反應活性,進而導致植物光合能力降低[23]。本研究Fv/Fm測定結果顯示,隨鹽脅迫時間增加,Fv/Fm逐漸下降,但在脅迫第5天、第7天時轉基因植株顯著(P<0.05)高于陰性植株,說明雖然轉基因和陰性對照玉米植株光反應活性均受到抑制,但轉基因植株的PSII活性中心受損程度較小,且具有較高光反應活性,從而保持了較高的光能捕獲能力與轉化效率。進一步說明鹽脅迫下,轉基因玉米植株的光合能力要高于陰性植株。
鹽脅迫對植物的危害來源之一就是滲透脅迫。高鹽導致環境滲透勢降低,抑制植物水分吸收或致使植物失水,造成植物生理干旱。本研究中,鹽脅迫后玉米葉片相對含水量下降,轉基因植株與陰性株之間無顯著差異,但恢復生長后,轉基因植株能夠快速吸收水分,葉片相對含水量高于陰性植株(圖4),說明其具有較高的恢復能力,推測這可能與滲透調節物質積累有關。研究表明,鹽脅迫后植物細胞常通過積累一些有機小分子物質(如脯氨酸、可溶性糖等)來維持較高的細胞質滲透壓[24]。本研究中玉米葉片中脯氨酸和可溶性糖含量受到脅迫后上升,且轉基因植株始終高于陰性。由此可見,遭受鹽脅迫后轉基因植株可以積累更多的有機調節物質,從而提高自身對高鹽脅迫的耐受性。
鹽脅迫導致植物體內產生大量活性氧,造成膜脂過氧化,破壞細胞膜完整性[25]。MDA是膜脂過氧化作用的產物之一,而相對電導率能夠代表細胞膜透性大小,二者可以反映植物細胞膜的損傷程度[26]。本研究的結果顯示,受到鹽脅迫后,玉米葉片中MDA含量上升,在第3天時達到最高,但轉基因植株低于陰性;測定3 d時葉片相對電導率發現轉基因植株膜透性也低于陰性株,說明在鹽脅迫條件下轉基因植株細胞膜損傷程度較小。
逆境脅迫下,植物細胞的存活與活性氧清除能力密切相關,而抗氧化酶的存在是清除活性氧的重要機制。Vijayalakshmi等[27]和 Farhangi-Abriz等[28]研究表明,鹽脅迫下植物通過提高抗氧化酶活性,能夠保護植物細胞免受氧化傷害,提高了植物的耐鹽能力。本研究測定了SOD、POD和CAT活性,在遭受鹽脅迫后抗氧化酶活性上升,且轉基因植株高于陰性植株,說明前者具有更高的活性氧清除和降低膜脂過氧化的能力。此外,有研究表明,脯氨酸等一些滲透調節物質也和植物抗氧化有關,不僅能夠維持細胞質滲透壓,還起到保護抗氧化酶、增強植物蛋白穩定性的作用[29-30],因而本研究中轉基因植株中脯氨酸含量的提高也使轉基因植株具有更高的抗氧化能力。
本研究通過RNA-seq和qRT-PCR分析發現,與陰性植株相比,轉基因植株中包括轉錄因子、熱激蛋白,抗氧化酶等大量鹽誘導基因的表達顯著增強。例如,WRKY轉錄因子,大量研究表明,其參與植物多種生理生化過程,包括應對多種非生物脅迫、參與葉片衰老及次生代謝等[31-34]。研究中發現,在受到鹽脅迫后,轉基因株系中WRKY基因AC209050.3_FG003的表達能在短時間內迅速升高,而在陰性材料中表達量變化幅度較小。熱激蛋白作為分子伴侶,能夠防止蛋白質降解,保護細胞免受傷害,相關研究表明其在植物抗逆方面發揮著重要作用[35-37]。結果顯示,熱激蛋白基因GRMZM2G007729在轉基因株系中高表達,且上升幅度較大。此外,還有一些與抗氧化相關基因,如GRMZM2G044383、GRMZM2G074743等,在轉基因植株中的表達顯著高于陰性植株。研究表明,這些基因在活性氧清除方面起重要作用[38-39]。因此,上述抗逆基因的增強表達很可能是導致轉基因玉米植株耐鹽性提高的關鍵。
4 結論
ABP9的增強表達提高了大量抗逆相關基因的表達,導致鹽脅迫下其抗氧化能力、滲透調解能力、膜完整性和光合能力的提高。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
