甜高粱FAD7基因的功能分析
2019-06-04 07:04:44宋玉雙隋娜
生物技術通報 2019年5期
關鍵詞:植物
宋玉雙 隋娜
(山東師范大學生命科學學院,濟南 250014)
溫度對植物的生長發育和地理分布起著重要的作用,是極其重要的環境因子之一。低溫會降低作物的產量和品質,影響作物的產量和品質[1]。
在低溫條件下,植物體內的親水蛋白、可溶性糖等滲透調節物質大量增加,膜組成改變[2]。低溫脅迫首先傷害的是植物細胞的細胞膜,低溫脅迫下,細胞膜由液晶相逐漸轉變為凝膠相,膜流動性降低、離子滲漏和膜蛋白失活等現象[3]。生物膜是細胞與外界環境聯系的界面,生物膜的基本組成成分之一是多不飽和脂肪酸(PUFAs),PUFAs通常是指含有兩個或兩個以上雙鍵的長鏈脂肪酸,具有重要的生理功能[4]。植物體內膜脂的PUFAs約占總脂肪酸的60%,為適應低溫脅迫,植物往往會通過改變自身不飽和脂肪酸含量進而來調整膜的流動性,從而維持適宜于蛋白發揮活性的環境[5]。而脂肪酸不飽和水平的變化主要通過調節脂肪酸去飽和酶的活性來實現[6-7]。
在植物中,脂肪酸去飽和酶可分為ω-3型(FAD2、FAD6)和 ω-6型(FAD3、FAD7、FAD8)兩大類,催化在脂肪酸鏈的特定位置形成雙鍵,從而產生特定的不飽和脂肪酸。ω-3脂肪酸去飽和酶是不飽和脂肪酸合成途徑中催化16∶2(7,10)或18∶2(9,12)轉化為 16∶3(7,1 0,13)或 18∶3(6,9,12)的關鍵酶,使脂肪酸形成第三個雙鍵。
近年來,已經有許多植物的ω-3脂肪酸去飽和酶基因被克隆,如馬鈴薯[8]、黃瓜[9]、菊花[10]、紫蘇[11]和播娘蒿[12]等。研究也發現這些脂肪酸去飽和酶基因在改變植物膜脂脂肪酸的組成、葉綠體的發育以及葉片成熟過程中三烯脂肪酸含量的增加、抗冷性的增強、低溫光抑制后光合能力的恢復等方面具有重要作用。轉AtFAD8基因煙草的MDA含量明顯低于野生型植株,而抗氧化酶活性明顯高于野生型植株,表明AtFAD8基因通過增加轉基因煙草的抗氧化能力而提高煙草幼苗的耐冷性[13]。將煙草中ω-3脂肪酸去飽和酶的編碼基因沉默會導致突變植株的三烯脂肪酸比野生型植株明顯減少[14]。于超[15]發現過量表達番茄內質網ω-3脂肪酸去飽和酶基因FAD3能夠增強番茄植株的耐冷性,而抑制該基因表達可提高番茄植株的耐熱性。陳瑋國等[16]發現油葵FAD2-5可能參與了油葵對低溫脅迫應答的調節,并且其表達具有晝夜節律性。曹英萍等[17]發現OsFAD7和OsFAD8的 mRNA 在低溫下上升,并且mRNA的表達具有晝夜節律性。
甜高粱是單子葉禾本科高粱屬C4植物,其莖稈含糖量極高,具“二代甘蔗”的美名[18]。甜高粱不僅生物量極高,還耐鹽堿、耐旱、耐澇、耐高溫、耐嚴寒[19]。甜高粱對土壤的適應能力很強,pH值為5.0-8.5,均能很好生長。適應栽培的區域廣泛,10℃以上積溫2 600-4 500℃的地區(從海南島至黑龍江),均可栽培。高粱籽粒可食用、飼用、釀酒;莖稈可做飼料、制糖等,稈渣還可飼用、制紙等[20]。因此,甜高粱具有較大的生物學優勢以及經濟學優勢。而對甜高粱這些優良的特性進行研究有利于我們對其他作物進行基因改造,獲得耐脅迫性的作物。
本研究將從甜高粱分離并克隆得到的ω-3脂肪酸去飽和酶基因FAD7在野生型擬南芥中過量表達,測定了在低溫處理條件下野生型和過表達植株的Fo、Fv/Fm、電導率、MDA、超氧陰離子自由基和過氧化氫含量等指標,闡明SbFAD7在植物抗冷中的作用,旨為探索植物抗冷的機理奠定基礎。
1 材料與方法
1.1 材料
甜高粱自交系M-81E種子,擬南芥野生型種子,大腸桿菌菌株(DH5α),農桿菌菌株(GV3101)。反轉錄試劑盒購自華越洋,植物總RNA提取試劑盒、質粒小提試劑盒膠回收試劑盒等均購自天根生化科技有限公司。
1.2 方法
1.2.1SbFAD7基因的克隆及生物信息學分析 將沙培的長至三葉一心期的甜高粱取材,提取其RNA進行PCR擴增,然后將得到的PCR反應產物進行瓊脂糖凝膠電泳檢測目的條帶以獲取基因的全長。所用引物為FAD-F、FAD-R,序列見表1。利用NCBI數據庫和生物信息學軟件對SbFAD7的蛋白結構、同源性和進化進行分析。
1.2.2 甜高粱FAD7基因的表達分析 將長至三葉一心時期的甜高粱一部分用0、50、100、150和200mmol/L NaCl溶液處理48 h,一部分用100 mmol/L NaCl溶液處理0、12、24、36、48 h,一部分用4℃處理0、4、8、12、24 h后取材提RNA,用Beacon Designer 7軟件設計RT-PCR引物,引物為FAD-S和FAD-A。選用甜高粱的β-actin基因作為內參,引物為Sbactin-S和Sbactin-A。引物序列如表1。
1.2.3 擬南芥過表達植株中SbFAD7基因的表達量分析 以野生型,過表達植株的RNA為模板,用設計的qPCR特異性引物進行基因的定量分析。利用擬南芥Actin2基因的引物作為內參,引物分別為AtActin2-S和AtActin2-A,序列如表1。

表1 PCR和RT-PCR所用的引物序列
1.2.4 轉基因擬南芥中脂肪酸含量的測定 采用毛細管氣相色譜法測定野生型和過表達植株中脂肪酸的含量。將葉片在研缽中研磨后放入105℃的恒溫干燥箱烘至恒重。在10 mL比色管中加入樣品0.5 g,再加入2 mL提取液(苯∶石油醚=1∶1),靜置 30 min 以上。然后加入1 mL 0.4 N 的 KOH/CH3OH溶液,靜置10 min以上。加入超純水定容,待上層液澄清后,將0.25 mL的上清液移入安培瓶中,在N2下吹干,再加入50 μL正己烷,最后取0.5 μL進樣分析。每個處理均做3個重復。
1.2.5 Fo和Fv/Fm的測定 葉片暗適應20-30 min后,以英國Hansatech公司生產的FMS2便攜調制式熒光儀測定Fo、Fm等熒光參數。暗適應下可變熒光可根據公式計算:
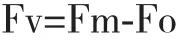
式中,Fo:初始熒光,即PSII反應中心處于完全開放時的熒光產量;Fm:最大熒光產量,即PSII反應中心處于完全關閉時的熒光產量;Fv:暗適應下的可變熒光,反應QA的氧化還原狀態;Fv/Fm:暗適應下PSII的最大光化學效率。
1.2.6 電導率的測定 將長勢差不多的葉片放入具塞試管中,先用去離子水洗3次,然后加15 mL去離子水置于真空泵中抽氣30 min,振蕩器上振蕩2 h,靜置搖勻,測定初電導。然后沸水煮30 min,冷卻至室溫10 min,測定終電導。根據公式計算:
相對電導率(%)=(初電導-空白)×100/(終電導-空白)。
1.2.7 MDA的測定 取擬南芥的葉片,稱重,剪碎,放入研缽中,加入少量石英砂和2 mL 0.1%的三氯乙酸(TCA),充分研磨,將勻漿移入試管中,用1.5mL 0.1%的三氯乙酸分別沖洗研缽兩次,將沖洗液都倒入試管中,再加入2.5 mL 0.5%的硫代巴比妥酸,搖勻,沸水浴10 min后,立即取出放入冷水中,冷卻后,3 000 r/min離心10 min,取上清液量其體積,以0.5%的硫代巴比妥酸溶液為空白對照,用分光光度計分別測量532 nm和600 nm的吸光度。根據公式計算MDA的含量:
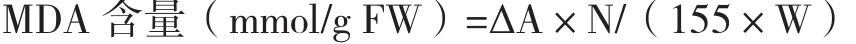
式中:ΔA:A532和A600之差,N:上清液總體積,155為1 mmol反應產物在532 nm處的吸收系數,W:稱取植物材料的鮮重(g)。
1.2.8 超氧陰離子自由基含量的測定 稱取植物材料 0.3 g,加 5 mL 預冷的 50 mmol/L PBS(pH 7.8)在冰上研磨。勻漿液轉入10 mL離心管中,12 000 r/min離心20 min,冰上保存,此時上清液為的待測液。反應體系:1 mL待測液+0.8 mL 50 mmol/L PBS(pH 7.8)+0.2 mL 10 mmol/L 鹽酸羥胺,振蕩混勻,25℃溫浴1 h,再分別向各管中加1 mL 17 mmol/L對氨基苯磺酸,1 mL 7 mmol/L α-萘胺,振蕩混勻,25℃溫浴20 min,以標曲中的1號管作對照,測定530 nm處的吸光度值。以標準曲線計算O2-.的產生速率。
1.2.9 過氧化氫含量的測定 稱取植物材料0.2 g,加5 mL 4℃預冷的丙酮研磨提取。勻漿液轉入5mL離心管中,3 000 r/min離心10 min,上清液即為H2O2待測液。反應體系為:1 mL待測液+0.1 mL 20% TiCl4+0.2 mL濃氨水,待沉淀形成后3 000 r/min離心10 min,棄上清,用適量丙酮(5 mL)洗滌3到5次,最后用5 mL 2 mol/L H2SO4溶解沉淀后轉移至10 mL容量瓶中,用蒸餾水定容,以標曲中的1號管作對照,測定415 nm處的吸光度值。以標準曲線計算H2O2的含量。
2 結果
2.1 基因的克隆及生物信息學分析
SbFAD7基因的編碼區序列長度為1 356 bp,編碼452個氨基酸。從圖1可以看出,玉米的ω-3脂肪酸去飽和酶基因與甜高粱SbFAD7基因親緣關系最近。
2.2 甜高粱SbFAD7基因在不同處理下的表達分析
如圖2所示,SbFAD7基因的表達受到鹽和低溫的誘導,隨著鹽濃度的增加,SbFAD7基因的表達量逐漸升高,在200 mmol/L NaCl處理下,SbFAD7基因的表達量最高(圖2-A);隨著鹽和低溫處理時間的增加,SbFAD7基因的表達量也隨之增加,分別在鹽溶液處理48 h、低溫處理24 h時,SbFAD7基因的表達量達到最高(圖2-B和2-C)。
2.3 擬南芥過表達植株中SbFAD7的表達分析
擬南芥過表達植株的不同株系中,SbFAD7的轉錄水平和野生型相比都顯著提高,表明SbFAD7基因已成功轉入到擬南芥植株中(圖3)。其次,過表達植株OE1、OE13和OE17中SbFAD7的轉錄水平相對于其他株系來說比較高,因此我們選擇OE1、OE13和OE17進行后期生理指標的驗證。

圖1 甜高粱FAD7基因的生物信息學分析

圖2 甜高粱FAD7基因在非生物脅迫下的表達量分析
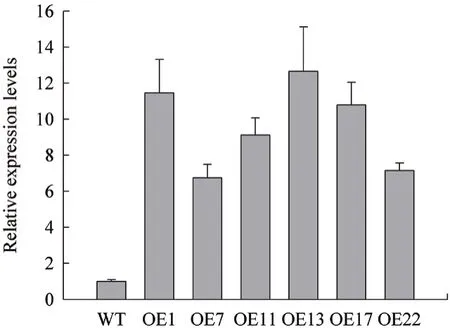
圖3 過表達擬南芥植株中SbFAD7的表達量分析
2.4 擬南芥過表達植株葉片的脂肪酸含量分析
如表2所示,相對于野生型植株,過表達植株的18∶2含量明顯降低,18∶1含量稍微降低,而18∶3含量明顯升高。從圖4也可看出,低溫處理后過表達植株的抗冷性明顯比野生型植株的抗冷性強。說明SbFAD7基因過量表達后,促進了18∶2向18∶3的轉化,提高了擬南芥植株的抗冷性。
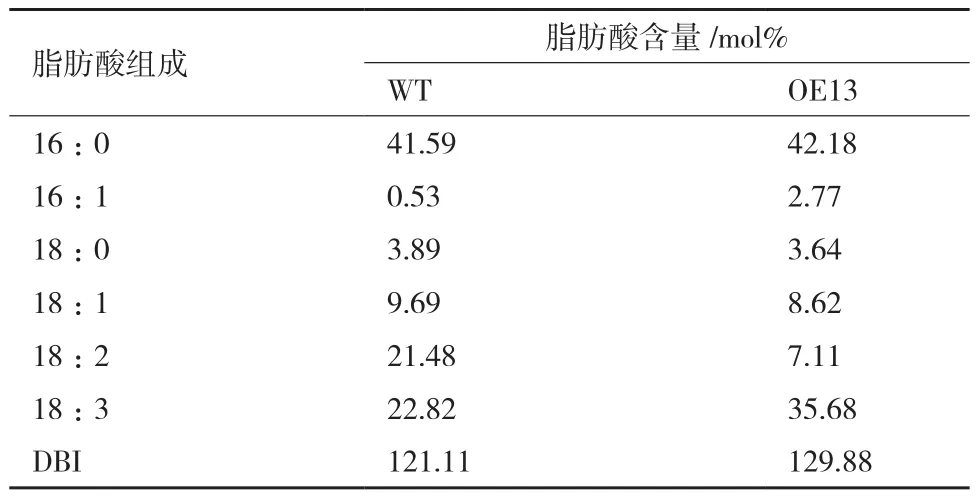
表2 擬南芥植株中脂肪酸含量
2.5 低溫處理對野生型和過表達植株Fo和Fv/Fm的影響
從圖5可看出,低溫脅迫下,野生型和過表達擬南芥的Fo均有所升高,而最大光化學效率降低,但相對于野生型來說,過表達植株OE1,OE13,OE17的Fo增加的少,而Fv/Fm的下降幅度較野生型要低,表明SbFAD7在擬南芥中的過量表達,能夠提高擬南芥的光化學效率,減輕低溫光抑制,提高擬南芥抵抗低溫的能力。

圖4 冷處理后野生型植株和過表達植株的生長情況
2.6 低溫處理對野生型和過表達植株電導率和MDA的影響
如圖6所示,低溫處理24 h條件下,野生型和過表達擬南芥的電導率和MDA含量都增加,但過表達植株的增加幅度明顯比對照低,這說明SbFAD7的過量表達能夠減輕低溫脅迫下膜脂過氧化水平,從而降低植物細胞遭受逆境傷害的程度。
2.7 低溫處理對野生型和過表達植株H2O2含量和O2-.含量的影響
由圖7中可以看出,在低溫處理24 h條件下,野生型和過表達擬南芥葉片的H2O2含量和O2-.產生速率均有所增加,而且過表達植株增加的量都比野生型增加的少,說明SbFAD7的過量表達能夠降低細胞中活性氧的產生。

圖5 冷處理后對擬南芥葉綠素熒光的影響

圖6 冷處理后擬南芥葉片中電導率和丙二醛含量的變化
3 討論
低溫脅迫是對植物尤其是農作物的生長發育威脅較大的一類非生物脅迫因子[21],因此研究農作物的耐冷基因一方面可以揭示農作物的耐冷機理,另一方面還可以提高農作物的耐冷性。
本研究發現SbFAD7基因的表達量與鹽濃度、鹽處理時間以及低溫處理時間成正相關,隨著鹽濃度的增大,基因的表達量也隨之增大,并在200 mmol/L下達到最大。分別隨著鹽和低溫處理時間的增加,SbFAD7基因的表達量也隨之增加。表明SbFAD7基因是組成型表達,其表達受高鹽和低溫誘導。
ω-3脂肪酸去飽和酶是催化18∶2轉化為18∶3的關鍵酶,研究表明,低溫處理下膜脂的三烯脂肪酸含量增加是對低溫脅迫的適應[22]。本試驗發現SbFAD7基因過表達之后促進了18∶2向18∶3的轉化。

圖7 冷處理對擬南芥中H2O2和O2-.的影響
在葉綠素熒光參數中,Fv/Fm是暗適應下PSII最大光化學效率,代表PSII原初光能轉化效率,是光抑制的重要指標[23]。本研究發現低溫處理24 h條件下,SbFAD7基因過表達植株的Fv/Fm較野生型要高,表明在低溫脅迫條件下,SbFAD7基因過表達能夠增強膜的穩定性,減輕PSII的光抑制。
膜脂脂肪酸含量的不飽和程度會影響膜的流動性[24],通過測定電導率來研究低溫處理對膜的傷害程度。低溫處理下,SbFAD7基因過表達植株的電導率比野生型的電導率低。由此說明,相對于野生型,轉基因植株的膜系統的傷害程度比較輕。
植物的防御系統在低溫等逆境條件下遭到破壞后,活性氧(H2O2和O2-.)會大量積累,引發膜脂過氧化作用,進而引發一系列的生理代謝紊亂[25]。MDA作為膜脂過氧化的產物,其含量代表膜脂過氧化的水平,反映植物細胞遭受逆境傷害的程度[26]。本研究發現在低溫處理24 h下,轉基因植株的MDA、H2O2含量以及O2-.產生速率均比野生型的低,這就說明SbFAD7基因能夠通過減少植物體內的活性氧的積累,降低膜脂過氧化的程度,進而保護膜結構,提高植物細胞的抗低溫能力。
本研究通過對甜高粱SbFAD7基因的研究發現該基因受低溫脅迫誘導,將該基因過表達擬南芥之后會促進亞油酸向亞麻酸的轉化,提高不飽和脂肪的含量,減輕低溫光抑制,減少了活性氧的積累,從而減輕膜脂過氧化程度,保持低溫脅迫下細胞膜的完整性,提高轉基因植株的抗低溫能力。
4 結論
本研究發現甜高粱ω-3脂肪酸去飽和酶FAD7基因能夠通過促進18∶2向18∶3的轉化,減輕PSII的光抑制,減少植物體內活性氧的積累,降低膜脂過氧化的程度,增強膜的穩定性來提高植物的抗冷性。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
