網(wǎng)紋蟒非入侵式采樣和線粒體基因組分析
2019-06-04 02:17:24何葉艷高旭龍趙阿勇
浙江農林大學學報 2019年3期
關鍵詞:物種
白 天,何葉艷,高旭龍,趙阿勇,何 珂
(浙江農林大學 動物科技學院,浙江 杭州 311300)
網(wǎng)紋蟒Python reticulatus屬于蛇亞目Serpentes原蛇附目Henophidia蟒科Pythonidae,廣泛分布于東南亞,在印度尼西亞存在海島變體[1],其皮張被認為是皮毛產業(yè)奢侈品的來源,是東南亞重要的經(jīng)濟自然資源。目前,該物種被世界自然保護聯(lián)盟評估為易危等級(vulnerable species,VU)[2],屬于瀕危動植物種國際貿易公約附錄Ⅱ物種。受捕獵影響,該物種在東南亞各國不同地區(qū)的種群遺傳多樣性已遭到破壞,亟待有計劃的種群復蘇和保護工作[3]。蛇類進化歷史久遠,生存環(huán)境復雜多樣,但其形態(tài)特征相對較為保守,加上趨同進化,造成可用于蛇類系統(tǒng)學研究的形態(tài)方面信息較少,因此,依據(jù)形態(tài)學信息很難解決蛇類系統(tǒng)關系。生物分子所具有的信息量大、完全不相同、較為真實記錄生物進化信息等特點,可彌補形態(tài)學信息的不足,越來越多地被應用到蛇類系統(tǒng)發(fā)生的研究中[4]。研究認為:作為一個與能量代謝有關的細胞器,真核生物的線粒體具有相對獨立的遺傳物質即線粒體DNA(mitochondrial DNA,mtDNA),研究線粒體基因組并利用線粒體DNA全序列分析進行系統(tǒng)建樹,能夠克服單個基因僅能帶來的有限信息,更加全面地揭示物種進化與系統(tǒng)發(fā)生。目前關于網(wǎng)紋蟒的分子生物學研究僅集中在幾個線粒體基因相關的系統(tǒng)分析上[3],在線粒體全基因組方面上存在欠缺。本研究擬采用無損傷性取樣,提取網(wǎng)紋蟒線粒體DNA,通過聚合酶鏈式反應(PCR)擴增拼接得到線粒體基因組全序列,以評估非入侵式采樣在蛇類研究中的可行性;以網(wǎng)紋蟒線粒體基因組序列信息和結構特征分析為依據(jù),結合GenBank中近緣物種(蟒科,蚺科Boidae和閃鱗蛇科Xenopeltidae)已發(fā)現(xiàn)的線粒體基因組序列,構建系統(tǒng)發(fā)生樹,探討蛇亞目的系統(tǒng)發(fā)生關系,并結合蛇亞目線粒體基因組不同的排列順序來討論蛇亞目線粒體基因組的重排。
1 材料與方法
1.1 試驗樣品
試驗所用網(wǎng)紋蟒的蛇蛻和新鮮糞便均來源于杭州動物園,于每日喂食時觀察并收集得到。糞便用體積分數(shù)70%乙醇浸泡,蛇蛻用體積分數(shù)95%的乙醇固定,帶回實驗室后保存于-20℃的冰箱中備用。
1.2 基因組DNA的提取
糞便樣品分剝離表面和內層部分2種,蛇蛻樣品剪碎后一部分室溫下研磨,另一部液氮研磨至粉碎。各樣品取3次重復,基因組DNA提取參考文獻[5]方法。經(jīng)NanoDropTM3300檢測計算質量濃度和純度D(260)/D(280)。
1.3 片段擴增
根據(jù)親緣物種的線粒體基因組序列設計擴增引物,采用高保真酶(PhantaSuper Fidelity DNA Polymerase,諾唯贊,南京)進行擴增,PCR產物經(jīng)電泳確定大小符合要求時,交由上海博尚生物公司測序。若PCR產物呈現(xiàn)多條帶,則采用割膠回收試劑盒獲取目的大小片段,用pMD-19T質粒(TaKaRa,大連)連接,選取陽性克隆后送出測序。
1.4 數(shù)據(jù)分析
(1)網(wǎng)紋蟒線粒體基因組各基因、tRNA長度和基因起始密碼子分析。用Dnastar軟件包拼接測序序列, 用 tRNA scan(http://lowelab.ucsc.edu/tRNAscan-SE/)確定 tRNA 的位置和類型, 采用 OGDRAW(https://chlorobox.mpimp-golm.mpg.de/OGDraw.html)繪制網(wǎng)紋蟒線粒體結構及基因結構圖。在分析過程中,網(wǎng)紋蟒記為 RP,以蟒科的緬甸蟒Python bivittaus(PB,NC_021479),印度蟒Python molurus(PM,NC_015812)和球蟒Python regius(PR, NC_007399)作為對比。 (2)根據(jù)相關基因、 控制區(qū)(control region)和線粒體基因組全序列建樹分析蟒科、蚺科和閃鱗蛇科的進化關系。用Mega 5.0軟件[6]比對序列及構建系統(tǒng)進化樹,蟒科物種為緬甸蟒、印度蟒和球蟒,蚺科物種為紅尾蚺Boa constrivtor(BC,NC_007398)和玫瑰蚺Charina trivirgat a(CT, GQ200595), 閃鱗蛇科物種為閃鱗蛇Xenpeltis unicolo r(XU,NC_007402),以原蛇亞目的圓環(huán)蛇Anilius scytale(AS,GQ200593)作為外類群。(3)蛇類具有多種線粒體結構排布次序,采用OGDRAW軟件繪制各蛇類線粒體基因結構圖可清楚地分析線粒體基因在進化過程中的變動。對分屬不同科屬的11個物種構建系統(tǒng)進化樹,分別為緬甸蟒、球蟒、紅尾蚺、閃鱗蛇和存在1個控制區(qū)和tRNA-WQANCY基因簇的細盲蛇科Leptotyphlopidae物種西南細盲蛇Rena humilis(RH,NC_005961),含有2個控制區(qū)和存在tRNA-P復制的蝰科Viperidae物種沖繩烙鐵頭Ovophis okinavensis(OO,NC_007397)和美洲蝮Agkistrodon piscivorus(AP,EF669477),含有2個控制區(qū)和存在tRNA-P復制的游蛇亞科Colubridae物種北美玉米蛇Pantherophis slowinskii(PS,NC_009769)和半棱鱗鏈蛇Dinodon semicarinatus(DS,NC_001945),rRNA 長度存在增加和含有 tRNA-L(Q)M和 tRNA-W(ANCY)結構的疣鱗蛇科Acrochordidae物種疣鱗蛇Acrochordus granulatus(AG,NC_007400)和管蛇科Cylindrophiidae物種紅尾管蛇Cylindrophis ruffus(CR, NC_007401)。
2 結果與分析
2.1 樣品基因組檢測
對提取到的DNA的檢測發(fā)現(xiàn),蛇蛻樣品DNA質量濃度最高,糞便表層次之,糞便內層最低;糞便內層樣品D(260)/D(280)最高, 說明樣品 DNA 純度較差, 其他樣品的D(260)/D(280)均為 1.7~1.8, 符合后續(xù)實驗要求(表1)。PCR擴增試驗顯示:室溫或液氮研磨,蛇蛻樣品中提取的基因組DNA均能擴增出長度為1 000~1 500 bp的片段;而來源于糞便表層或內層樣品的基因組擴增得到700~1 000 bp片段和1 000~1 500 bp片段的效果均不佳。

表1 所采用的樣品類型提取DNA和擴增情況Table 1 Genomic DNA extracted from different types of samples and the success rate of PCR
2.2 蟒科蛇類線粒體基因組分析
對PCR擴增產物進行測序和序列拼接,得到長度為17 641 bp的網(wǎng)紋蟒線粒體基因組完整序列(GenBank:MH410033)(圖1),序列含2個rRNA基因,13個編碼區(qū),22個tRNA基因和2個控制區(qū)(表2)。其中重鏈編碼28個基因,輕鏈編碼9個基因。包含的2個rRNA基因(12S rRNA和16S rRNA)位置和其余蛇類相同。除了NADH6(ND6)之外,其余12個蛋白編碼基因均位于重鏈上;所有蛋白基因都沒有內含子,但和相鄰的基因存在少許重疊或間隔。發(fā)現(xiàn)的22個tRNA基因存在蛇類特有的LQM和WANCY基因簇。2個控制區(qū)位于非編碼區(qū),長度分別為1 271和1 196 bp,序列相似度為96.68%。
對蟒科物種相關基因的長度、起始密碼子和終止密碼子的分析發(fā)現(xiàn),4個物種之間存在較高的同源性。網(wǎng)紋蟒的NADH2(ND2)終止密碼子和ATP8起始密碼子與其他物種不同,是物種所特有的;NADH6(ND6)基因變異最大,起始密碼子僅2個物種一致,終止密碼子緬甸蟒不同于其余3個物種。COX2基因和NADH1(ND1)較為保守,前者4個物種均以TA為終止密碼子,后者則使用單堿基T作為不完全終止密碼子。

表2 蟒科蛇類的線粒體蛋白質基因長度、起始密碼子、終止密碼子和rRNATable 2 Comparion of length,start codon and stop codon in Pythonidae
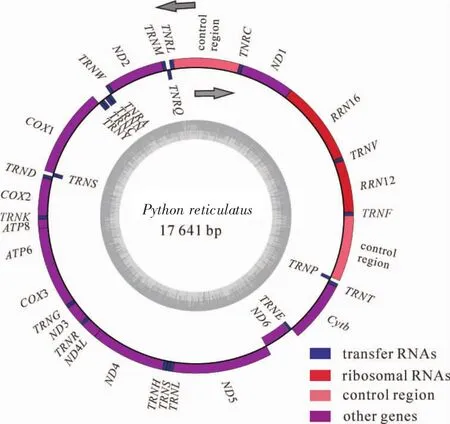
圖1 網(wǎng)紋蟒線粒體基因組結構示意圖圖1 Schematic diagram of mitochondrial genome structure in Python reticulatus

圖2 蟒科、閃鱗蛇科和蚺科的系統(tǒng)發(fā)育樹Figure 2 Phylogenetic relationship between Pythonidae,Xenopeltidae and Boidae
2.3 蟒科和蚺科物種進化分析
利用相關基因、控制區(qū)和線粒體基因組全序列構建系統(tǒng)樹。分析發(fā)現(xiàn):用蛋白基因構建的系統(tǒng)樹拓撲結構較為相近;以Cytb基因為例(圖2A),蟒科物種聚集的支持率為97%(PR,PM,PB和RP枝),就親緣關系而言,閃鱗蛇科(XU)比蚺科(BC和CT)更接近蟒科(圖2A)。用線粒體基因組全序列構建的系統(tǒng)樹則表明:蟒科(PR,PM,PB和RP枝)和蚺科(BC和CT枝)各自聚為一枝,支持率都為100%;同樣,閃鱗蛇科比蚺科更接近蟒科(支持率99%)(圖2B)。由于數(shù)據(jù)庫中玫瑰蚺(CT)的2個控制區(qū)長度僅為288和553 bp,和其他數(shù)據(jù)存在明顯差異,因此刪除該序列后對其他7個物種進行基于控制區(qū)的系統(tǒng)樹構建。分析發(fā)現(xiàn):各物種的2個獨立的控制區(qū)均先聚類,物種間的控制區(qū)聚類關系表明蟒科的支持率為100%,而閃鱗蛇、蚺科和圓環(huán)蛇則為另外一分支(圖2C)。以上結果表明:來源于同個物種的控制區(qū)相似度更高;相較于蚺科,閃鱗蛇科和蟒科的進化關系更接近。
2.4 蛇亞目物種線粒體結構排步分析
和其他脊椎動物線粒體基因組全序列的比較發(fā)現(xiàn),蛇亞目物種長度大小變化較大,主要原因為控制區(qū)的長度和數(shù)量不同,基因組上存在一些重復序列區(qū)域。對不同物種線粒體結構序列進行分析并構建系統(tǒng)樹(圖3),結果發(fā)現(xiàn)存在3種不同的基因結構,結構一主要存在于蟒科(RP和PR),閃鱗蛇科(XU),蚺科(BC)和疣鱗蛇科(AG 和 CR)中,含有 2 個控制區(qū)及 tRNA-L(Q)M和 tRNA-W(ANCY)結構; 結構二存在于蝮亞科(OO和AP)和游蛇科(DS和PS),和結構一的差異在于在tRNA-I和控制區(qū)1中多了1個tRNA-P;相比于結構二,結構三的tRNA-L(Q)M基因簇中,tRNA-Q產生了基因輕重鏈的轉化,形成了tRNA-LQM;而結構四(代表為盲蛇總目RH)則為僅存在1個控制區(qū),tRNA-L產生了移位,挪動至ND1前,而tRNA-L(Q)M和 tRNA-W(ANCY)通過移位分別轉變?yōu)?tRNA-IM和 tRNA-W(QANCY)。

圖3 不同基因結構的線粒體基因組系統(tǒng)進化樹及基因結構示意圖Figure 3 Phylogenetic tree of mitochondrion based on different gene structures and the rank of variable area
3 討論
3.1 非入侵式采樣在蛇類研究中的應用
在野生瀕危動物中采集血液和臟器等常規(guī)樣品非常困難,要建立瀕危動物的物種基因庫,亟待尋找新的采樣方法。研究認為:通過提取糞便DNA或其他生物痕跡DNA等非入侵式采樣方法能夠為遺傳學和生態(tài)學研究提供有效信息,在檢測種群大小、物種飲食結構和大量物種的激素信息上具有現(xiàn)實意義[7]。蛇類的蛻皮行為為非入侵式采樣提供了實例。KHEDKAR等[8]提取92份蛇蛻樣品基因組,通過DNA條形碼技術鑒定出了23種蛇類,但部分樣品不能有效進行擴增。石林春等[9]在利用蛇蛻鑒定中藥材中的蛇類物種來源時發(fā)現(xiàn),采用線粒體細胞色素C氧化酶I亞基(COI)作為引物,擴增片段不超過1 000 bp。網(wǎng)紋蟒是目前已知最長的蟒蛇,在東南亞國家捕殺嚴重,已被列為易危等級。本研究以糞便樣品和新鮮蛇蛻為材料提取基因組DNA,發(fā)現(xiàn)相較糞便樣品,蛇蛻樣品擴增得到的片段長度更長(1 000~1 500 bp),液氮研磨能有效提高DNA的提取品質,滿足后續(xù)實驗要求。糞便樣品由于提取的DNA品質和質量濃度不佳,在長片段擴增中效率低下,因此不建議使用。
3.2 蟒科和蚺科物種進化
本研究第1次完整地解析了網(wǎng)紋蟒的線粒體全基因組結構,填補了網(wǎng)紋蟒線粒體基因組全序列的空白。分析發(fā)現(xiàn):網(wǎng)紋蟒的基因結構、tRNA順序和控制區(qū)位置等均與蟒科物種一致,但也發(fā)現(xiàn)存在物種特異的特征,如NADH2以AGA作為終止密碼子,ATP8以GTG作為起始密碼子,NADH6以ATA作為起始密碼子,這3點均不同于其他蟒科已報道的物種(表2)。
關于蟒科和蚺科物種的分子系統(tǒng)相關研究,主要采用部分線粒體基因結合核基因進行;由于線粒體全基因不能通過簡單擴增得到,在分析物種進化過程中尚未廣泛使用[4,10]。但線粒體全基因組包含的信息更多,在解釋進化關系時更具有說服力[11]。本研究結合多個線粒體基因(Cytb,12S和16S)和多個核基因的序列,分析蟒科和蚺科物種的進化關系,認為蚺科在這3個物種中先分化出來,而蟒科和閃鱗蛇科始終聚成姐妹群關系。單基因和線粒體基因組的系統(tǒng)進化分析同樣得到上述研究結果 (圖2A,圖2B),進一步確定了蟒科、閃鱗蛇科和蚺科三者之間的進化關系[10]。
3.3 蛇亞目線粒體基因組的重排
線粒體基因排列次序在進化過程中較為穩(wěn)定,但在兩棲類、魚類和一些有袋類等類群中發(fā)現(xiàn)存在基因重復、基因重排和基因缺失等現(xiàn)象[11-12]。蛇類線粒體基因組在演化過程中出現(xiàn)了某些基因排列順序的變化,如OL缺失、tRNA-LQM基因簇和tRNA-WANCY基因簇中的基因重排、控制區(qū)重復、tRNA-L移位和tRNA-P的功能缺失等[11],進化程度高級的新蛇附目Caenophidia中重排更加多樣[13]。由此認為:具有共同基因重排順序的物種很有可能具有共同的祖先,因為不同物種間共享的基因排列順序,不可能是通過趨同進化而獨立產生的。
蟒科在分類上屬于蛇亞目原蛇附目。結合基因結構和順序分析,蟒科目前已報道的3個物種和本研究所報道的網(wǎng)紋蟒線粒體結構均體現(xiàn)出真蛇類物種的線粒體基因結構類型,存在tRNA-LQM基因簇和tRNA-WANCY基因簇,并且沒有發(fā)生tRNA-P的假基因化現(xiàn)象,存在2個控制區(qū),且結構基本統(tǒng)一。tRNA-LQM基因簇由早期真蛇類的tRNALeu發(fā)生移位形成。本研究發(fā)現(xiàn):除細盲蛇科物種(RH)外,所列舉的物種均含有該基因;而tRNA-WQANCY基因簇由tRNAGln移位生成,僅存在于細盲蛇科(RH);真蛇類tRNAPro假基因的出現(xiàn),則表現(xiàn)為控制區(qū)2附近的tRNAPro假基因化(OO和AP)和控制區(qū)1附近的tRNAPro缺失(RH)。雙控制區(qū)在其他爬行動物[14]、 兩棲類[13]和鳥類[15]中也有存在, 重復控制區(qū)認為可能是功能上(使得可以從多個位點啟動復制和轉錄,提高效率)和進化選擇上的優(yōu)勢,多次復制和插入形成并維持雙控制區(qū)[12]。
對控制區(qū)的聚類分析發(fā)現(xiàn):物種中獨立存在的2個控制區(qū)在系統(tǒng)發(fā)生過程中并非按排列的位置成簇聚集,而是在各物種間聚成一枝(圖3C),這和之前其他蛇類中描述的結果保持一致[12]。控制區(qū)重復事件認為發(fā)生在7 000萬a前[12]:在原始類群蠕蛇附目Scolecophidia和細盲蛇科中僅存在1個控制區(qū),隨著蛇類演化,真蛇類線粒體中出現(xiàn)了控制區(qū)重復的事件,而在此基礎上基因重排演化出tRNA的移位和轉化。但聚類結果中2個控制區(qū)并沒有按照2個分布進行聚類,主要原因則是因為在物種進化過程中發(fā)生了協(xié)同進化,導致同個物種的2個控制區(qū)形成了較高的同源性,相似度達100%[12]。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創(chuàng)造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
