干旱荒漠區植物生態位對水鹽的響應
2019-05-31 00:51:30郄亞棟滕德雄呂光輝
生態學報 2019年8期
郄亞棟,滕德雄,呂光輝
1 綠洲生態教育部重點實驗室,烏魯木齊 830046 2 新疆大學資源與環境科學學院,烏魯木齊 830046 3 新疆大學干旱生態環境研究所,烏魯木齊 830046
生態位廣泛地指在一個由一系列環境軸所定義的n維空間內種群可以維持存活的能力,能夠反映物種在群落中的功能地位,并且可以定量地研究種內、種間、生境三者之間的相互關系[1- 2],已成為現在生態學研究中的一個重要理論,特別是在種間聯系、群落結構和功能、生物資源供需、物種進化等方面的研究中有重要作用,為闡釋自然生物群落中物種共存與競爭機制提供了夯實的根基[3- 5]。
目前,全球氣候變化的持續正在強烈地影響著生態系統的功能,導致生物生境的改變,并對生物的生存已構成嚴重的危險,極端地情況下可能會引起全球生物多樣性的喪失[6- 7]。然而,不同的物種或生態系統對生境壓力的脆弱性不同,具有不同的生態位特征[3,8- 10]。以往研究表明,兩個物種在競爭相同環境資源的同時,如果其他所有的生態因素不變,那么兩者可能是不能共存的[8,11]。種群對資源利用能力的大小體現了自身在群落生態空間內的地位,也就是生態位[1,12]。一些學者認為,普遍的物種或生態系統都具有較廣泛的環境適應范圍,其生態位寬度較大,分布范圍狹窄的物種或生態系統,其生態位寬度較小[13- 14]。然而,一些研究發現,物種在群落生態空間的地位是可以通過自身的生態策略扭轉的,認為競爭能力較弱的物種可能會在多維的生態空間下選擇其中的一小部分進行繁殖和生存,以此逃避競爭的排斥作用,隨著時間的推移,物種反而會在受限制的生態空間內占據主導地位,對該資源梯度下其他物種的生態空間產生作用,甚至可能會將其他物種排除在外[12,15]。賀強等[16]在黃河三角洲濕地的水鹽梯度上發現,物種出現生態位分化現象,且同資源緯度下的物種間生態位重疊較大;García-Baquero等[17]、劉揚等[18]和彭舜磊等[19]對植物生態位分析均發現,土壤水分是引起生態位分化的驅動因子;Iturrate等[20]發現土壤水分和鹽分共同引起了植物生態位分化,并認為是決定凍原物種共存和群落組成的重要因子;張東梅等[21]在花馬池鹽湖地區發現,鹽分梯度上生態位重疊值與Ochiai指數相關程度均高于水分梯度。上述對植物生態位的研究均是集中在半干旱、高寒或濕潤地區,對干旱荒漠區植物生態位的研究較少,并且在干旱荒漠區由于物種稀少、群落單一和系統脆弱對環境變化響應相對敏感[22-24],土壤水分和鹽分更是其植物生存和群落組成的驅動因子和限制因子。然而,整合水鹽因子在自然梯度下對植物生態位研究更是缺乏系統性。
艾比湖流域,生物資源較豐富,景觀類型多樣,兼具荒漠生態系統和濕地沼澤生態系統的雙重特征,是研究干旱荒漠區植物生態位的適合對象[24]。本文在研究干旱荒漠區土壤水分、鹽分和植物生態位特征的基礎上,分析了不同水鹽梯度下植物的群落組成和植物生態位響應土壤水鹽的變化趨勢,以回答以下科學問題:(1)沿水鹽梯度植物群落的特征;(2)水鹽梯度下群落中植物的生態位特征;(3)干旱荒漠區植物生態位對土壤水鹽響應,以期為艾比湖流域植物生態位研究提供基礎科學數據,為干旱荒漠區植物的利用與保護提供理論支撐。
1 材料和方法
1.1 研究區概況
艾比湖濕地國家級自然保護區位于阿拉山口東南方向,地理坐標為44°30′—45°09′N,82°36′—83°50′E,屬于濕地-荒漠森林復合生態系統類型的自然保護區。保護區總面積2670.85 km2,海拔189 m。該地區屬于溫帶大陸性干旱氣候,夏季酷熱,冬季嚴寒,干旱少雨。1月份平均氣溫-16℃,極端最低溫-36℃;7月份平均氣溫27℃,極端最高溫44℃,年平均降雨量100 mm左右,年平均蒸發量1600 mm以上,全年大風日數約170d,最大風速55 m/s。區內土壤類型為灰漠土、灰棕漠土和風沙土,隱域性土壤為鹽(鹽漬化)土、草甸土和沼澤土。區內土壤鹽漬化程度較高,其主要形成以湖區為中心向四周呈環狀遞減格局,或受河流和氣候影響呈垂直于河的帶狀減少分布,類型有原生鹽堿化和次生鹽堿化[25]。研究區主要植物種類有胡楊(Populuseuphratica)、梭梭(Haloxylonammodendron)、白刺(Nitrariatangutorum)、琵琶柴(Reaumuriasoongorica)、鹽爪爪(Kalidiumfoliatum)、駱駝刺(Alhagisparsifolia)、羅布麻(Apocynumvenetum)、花花柴(Kareliniacaspica)和蘆葦(Phragmitesaustralis)等。
1.2 研究方法
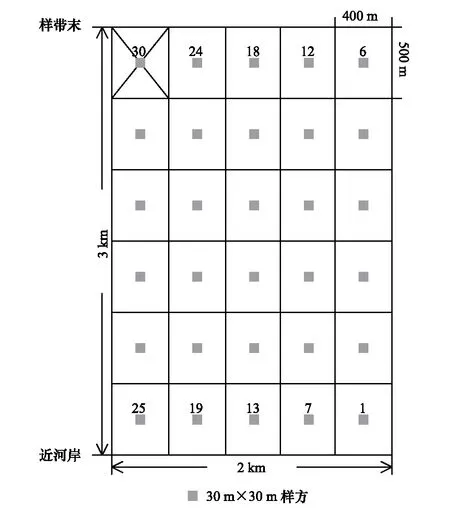
圖1 樣方分布圖Fig.1 The location of the plots
1.2.1野外調查和樣品采集
于2014年7—8月份(植物生長旺季)在艾比湖濕地保護區內東大橋管護站西北方向進行野外調查。垂直于阿其克蘇河,布置2 km×3 km樣地,將其等分成30個400 m×500 m小樣地(圖 1)。于每個小樣地中心部位設置一個30 m×30 m樣方(圖 1),進行群落調查和樣品采集。在調查樣方過程中,記錄每個樣方中心點的地理坐標、海拔,以及樣方內出現的所有物種的種類,對于不能夠識別的物種,采集標本并編號,帶回實驗室鑒別;對于喬木,記錄其多度、高度、冠幅、胸徑;對于灌木和草本,記錄高度、叢(株)數、叢幅、蓋度。樣方清查后,分別在30個樣方內按照冠下土、裸地土挖取0—50 cm的土壤剖面,并將兩種類型的土壤用四分法混合均勻作為該樣方的土壤樣品,取適量裝入鋁盒中,并稱其重量,用于測定含水量的土壤鮮重數據;再取適量樣品土壤裝入密封袋內,將其帶回實驗室用于鹽分含量的測定。
1.2.2樣品分析
用烘干稱重法測定土壤的含水量,用殘渣烘干法(土水比1∶5)測定土壤的含鹽量。
1.2.3水鹽協同聚類
依據水分和鹽分含量對樣方進行系統聚類[26],分析水鹽協同作用對群落生態位的影響。由聚類分析樹的結果,將樣方劃分成不同群落類型,以不同群落下的樣方數作為“資源狀態”,計算不同物種在不同群落中的重要值,進而得出它們在不同水鹽梯度下的生態位寬度以及生態位重疊,并分析它們在不同水鹽梯度下群落中的功能地位以及相互之間利用資源的狀態。
1.2.4物種重要值的計算
根據荒漠植物群落中植物的高度、個體數量、頻度和蓋度等數量指標,分別統計和計算喬木、灌木和草本物種的重要值:重要值喬木=(相對多度+相對頻度+相對蓋度+相對高度)/4,重要值灌木=(相對多度+相對蓋度+相對頻度)/3,重要值草本=(相對蓋度+相對頻度)/2[27]。其中,相對多度=(某種植物的多度/全部植物的多度之和)×100%,相對頻度=(某種植物的頻度/全部植物的頻度之和)×100%,相對蓋度=(某種植物的蓋度/全部植物的蓋度之和)×100%,相對高度=(某種植物的平均高度/全部植物的平均高度之和)×100%。
1.2.5種群生態位的研究方法
(1)生態位寬度采用Levins[28]提出后經Colwell[29]加權修改的計算方法:
式中,Bi為物種i的生態位寬度,Pij=nij/Ni,nij為物種i在資源j上的重要值,Ni表示種i在所有資源等級上的重要值之和;Pij代表物種i在資源j上的重要值占該物種在所有資源等級上的重要值總和的比例,r為樣方數。
(2)生態位重疊采用Pianka生態位重疊指數[30]:
式中,Oij為物種i和j的生態位重疊指數,Pij和Pkj分別為第i和第k物種在第j個資源上的重要值,r為樣方數。
1.2.6群落生態位寬度的計算方法
根據物種的重要值,對其生態位寬度測度值進行加權,把加權后的總和定義為群落的生態位寬度。
1.2.7變系數模型
利用變系數模型(Varying Coefficient Models)[31]進行群落生態位寬度對水鹽響應特征的分析。模型結構如下:
Bi=α(Ui)Xi+b(Ui)+εi,i=1,2,…,n
Bj=β(Xj)Uj+b(Xj)+μj,j=1,2,…,n
式中,Bi為群落生態位寬度;Xi為土壤鹽分,Ui為土壤水分,α(Ui)代表生態位鹽效應指數,b(Ui)為該模型的截距項,i為樣方序號;Bj為群落生態位寬度;Xj為土壤鹽分,Uj為土壤水分,β(Xj)代表生態位水效應指數,b(Xj)是該模型的截距項,j為樣方序號。
1.2.8數據分析
水鹽聚類分析在SPSS 20.0中進行;植物生態位對土壤水鹽的響應特征在R語言(Version 3.4.3,R Core Team)中采用變系數模型分析,并用ggplot2程序包進行繪圖;利用Microsoft Office Excel 2013對數據進行前期整理和計算;運用Origin 8.0對物種生態位測度值作圖。
2 結果和分析
2.1 植物群落調查結果及水鹽聚類
樣地中共有多年生木本植物9科16屬16種,皆為被子植物,其中占優勢科的屬和種主要有豆科(3屬3種)和藜科(4屬4種)。為防止偶然結果,剔除偶見種3種,剩余8科13種,其中喬木1種:胡楊,小喬木2種:梭梭和檉柳(Tamarixramosissima),灌木2種:白刺和鈴鐺刺(Halimodendronhalodendron);半灌木5種:琵琶柴、駱駝刺、羅布麻、鹽爪爪和鹽節木(Halocnemumstrobilaceum);草本3種:甘草(Glycyrrhizauralensis)、蘆葦和花花柴。因20和30號樣方采集數據不完全而被剔除,剩余28個樣方數據,基于這28個樣方土壤的水分和鹽分進行系統聚類,將聚類結果與實際的生態意義相結合,可劃分為3個水鹽梯度群叢(表1),命名結果及環境特征如下:
群叢I,胡楊-梭梭-羅布麻-蘆葦(Ass.Populuseuphratica-Haloxylonammodendron-Apocynumvenetum-Phragmitesaustralis),包含樣地1、2和7,主要分布于阿其克蘇河近岸邊及地下水較淺的地區。該植被系統的主要群落為胡楊群落,鈴鐺刺、琵琶柴和白刺為主要伴生種,未出現種有鹽爪爪、鹽節木和花花柴。該群叢土壤含水量、含鹽量最高。
群叢II,鹽爪爪||胡楊-梭梭-琵琶柴(Ass.Kalidiumfoliatum‖Populuseuphratica-Haloxylonammodendron-Reaumuriasoongorica),包括3—6,8—17,19,21—23和25—28共22個樣方,主要分布于阿其克蘇河的河灘邊緣地帶,13種植物均有出現,為艾比湖流域的主要荒漠植物群落,多沙土與鹽漬灌叢土。植物群落以沙生植物和鹽生植物為主,主要伴生種有檉柳、鹽節木、花花柴和蘆葦。該群叢土壤含水量、含鹽量較高。
群叢III,駱駝刺||胡楊-梭梭-琵琶柴-花花柴(Ass.Alhagisparsifolia‖Populuseuphratica-Haloxylonammodendron-Reaumuriasoongorica-Kareliniacaspica),包含樣地18,24和29,主要分布在距河岸帶較遠的荒沙土上,植物組成種類簡單,植被蓋度較低,檉柳和蘆葦為主要伴生種,未出現種有鈴鐺刺和甘草。該群叢土壤含水量、含鹽量最低。
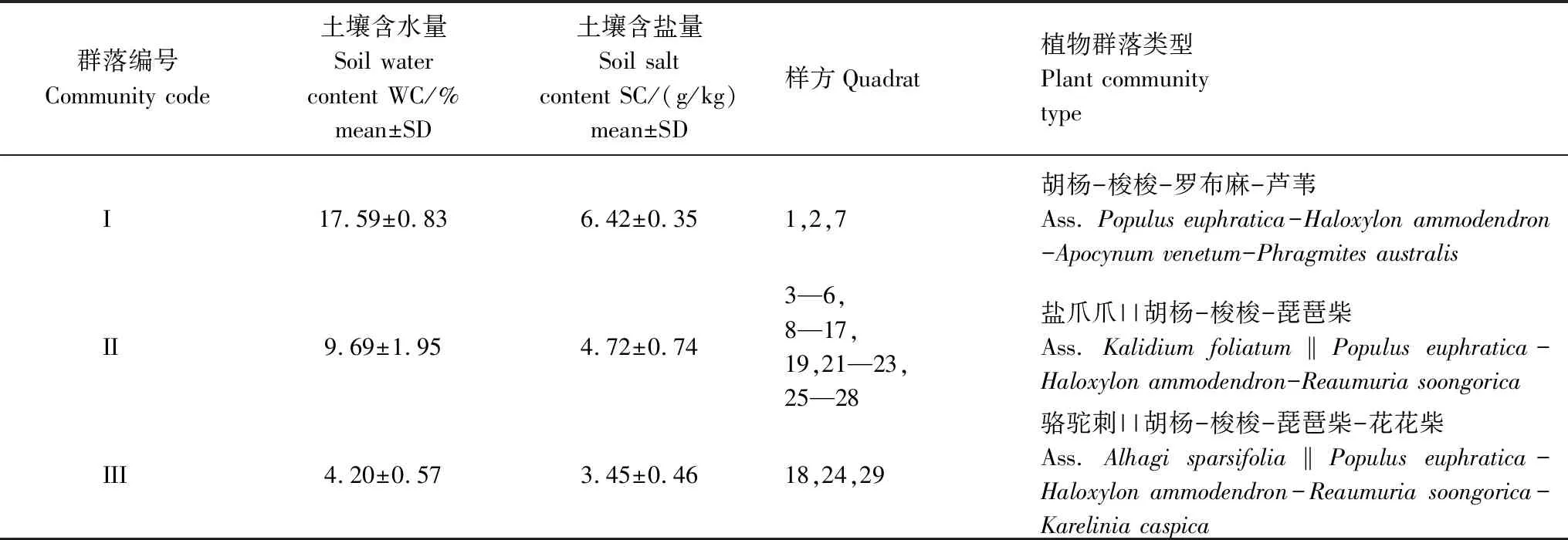
表1 研究區植物群落類型及環境特征(平均值)
I:高水鹽土壤環境的群落I,Community I of the high saltwater soil environments;II:中水鹽土壤環境的群落II,Community II of the moderate saltwater soil environments;III:低水鹽土壤環境的群落III,Community III of the low saltwater soil environments
2.2 不同水鹽梯度群落下種群的生態位寬度和生態位重疊
對不同水鹽梯度下的種群生態位寬度計算結果表明,I類型下,胡楊、琵琶柴和梭梭具有較高的生態位寬度值,分別為0.994、0.994和0.977,但檉柳、駱駝刺和甘草的生態位寬度值較低,均為0.333;II類型下,白刺和琵琶柴具有較高的生態位寬度,分別為0.899和0.879,而鈴鐺刺和甘草明顯低于其他物種,分別為0.045和0.066;III類型下,梭梭、花花柴和琵琶柴具有較高的生態位寬度,分別為0.992、0.967和0.922,而鹽節木的生態位寬度較低,為0.333(表2)。總的來說,不同水鹽梯度下同一物種的生態位寬度存在差異,不同物種在同一水鹽梯度下的生態位寬度也有所差異,其中琵琶柴在各水鹽梯度均具有較高的生態位寬度值,而甘草和鹽節木在各水鹽梯度的生態位寬度值均較小。
對不同植物群落類型下種群生態位重疊的計算結果表明,在群落類型I中,胡楊、梭梭和琵琶柴與其他物種的生態位重疊均較高,檉柳和駱駝刺的生態位重疊值最高,為1,但它們兩者與其他物種的生態位重疊值都不高,其中分別與鈴鐺刺和甘草的重疊值最低,均為0(表3)。在群落類型II中,白刺和琵琶柴與其他物種的生態位重疊值較高,其中白刺和琵琶柴的重疊值最高,為0.917,鈴鐺刺和甘草與其他物種的生態位重疊均較低,其中鈴鐺刺與檉柳、鹽爪爪、鹽節木、花花柴和甘草以及檉柳與甘草的重疊均最低,為0(表4)。群落類型III下,胡楊、梭梭、琵琶柴和花花柴與其他物種的生態位重疊值均較高,但檉柳和白刺的生態位重疊值最高,為0.998,鹽節木和蘆葦的重疊值最低,為0(表5)。

表2 不同水鹽梯度下13種植物的生態位寬度值
符號“—”代表此水鹽梯度下不存在該物種
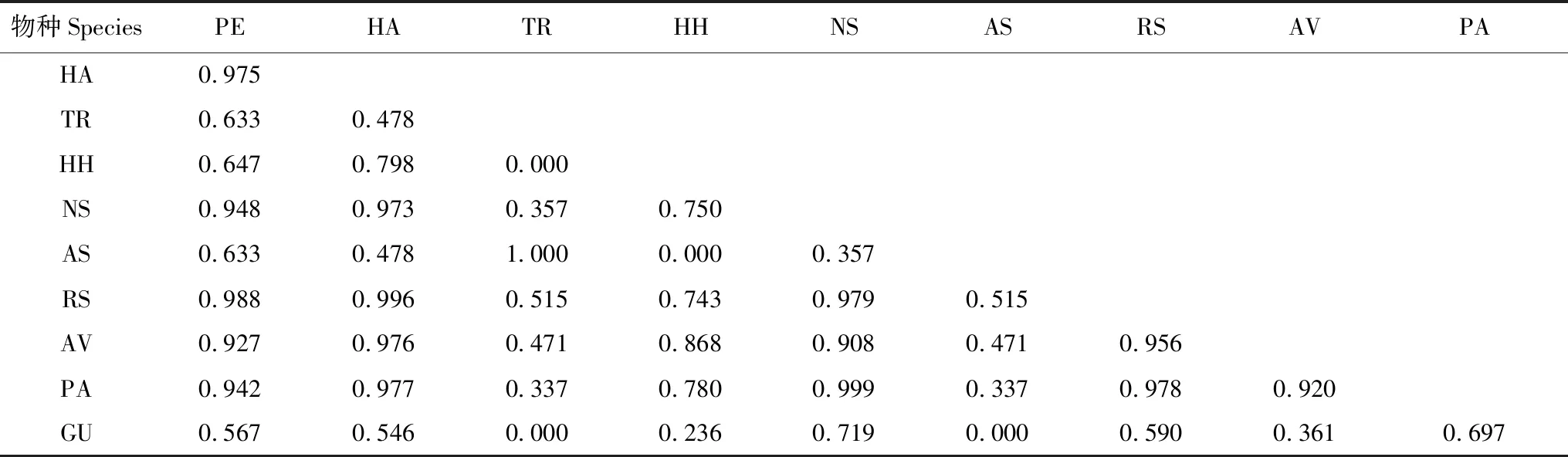
表3 群落類型I的植物生態位重疊值
字母的含義見表2
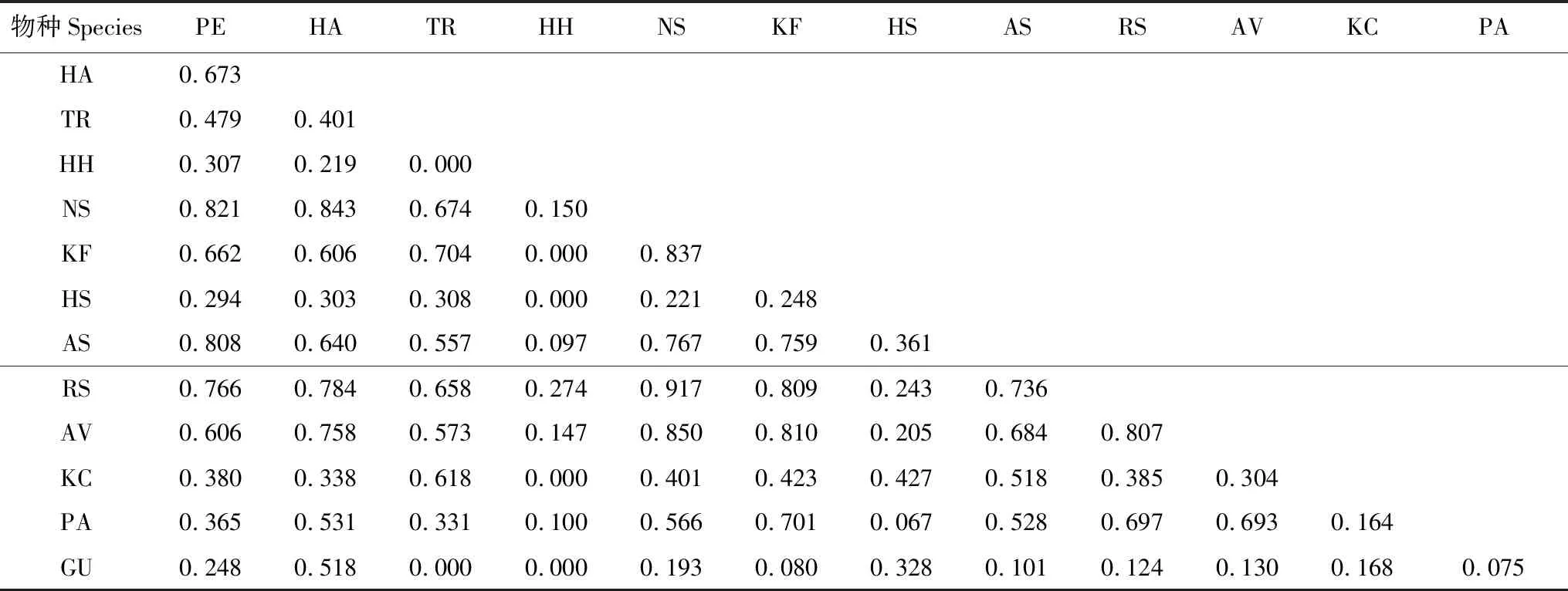
表4 群落類型II的植物生態位重疊值
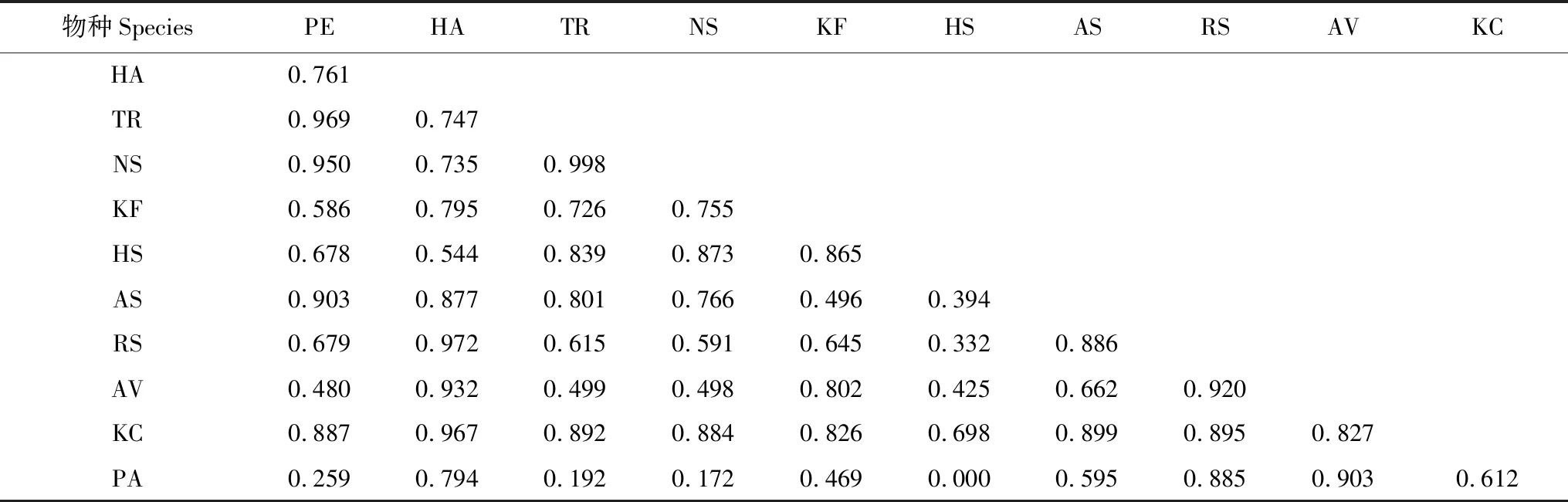
表5 群落類型III的植物生態位重疊值
綜合生態位寬度和生態位重疊值的分析表明,植物群落類型I、II和III下生態位寬度大于0.8的物種分別占所有物種的60.00%,15.38%,45.45%,而低于0.2的分別占有0、23.08%、0;植物群落類型I、II和III中生態位重疊值超過0.8的種對分別占所有種的37.78%,11.54%,43.64%,而低于0.2的分別占有8.89%,24.36%,5.45%(圖2),說明群落類型I和III種群的生態位寬度較大,并且具有較高的生態位重疊,而群落II種群的生態位寬度較低,其生態位重疊值也較低。在群落水平上考慮種群的生態位寬度與生態位重疊之間的關系可見(圖3),兩者之間在一定程度上具有顯著的相關關系(R2=0.45,P<0.01),生態位寬度較大的種群,其生態位重疊值也較大,而生態位寬度較小的種群,生態位重疊值也較小。
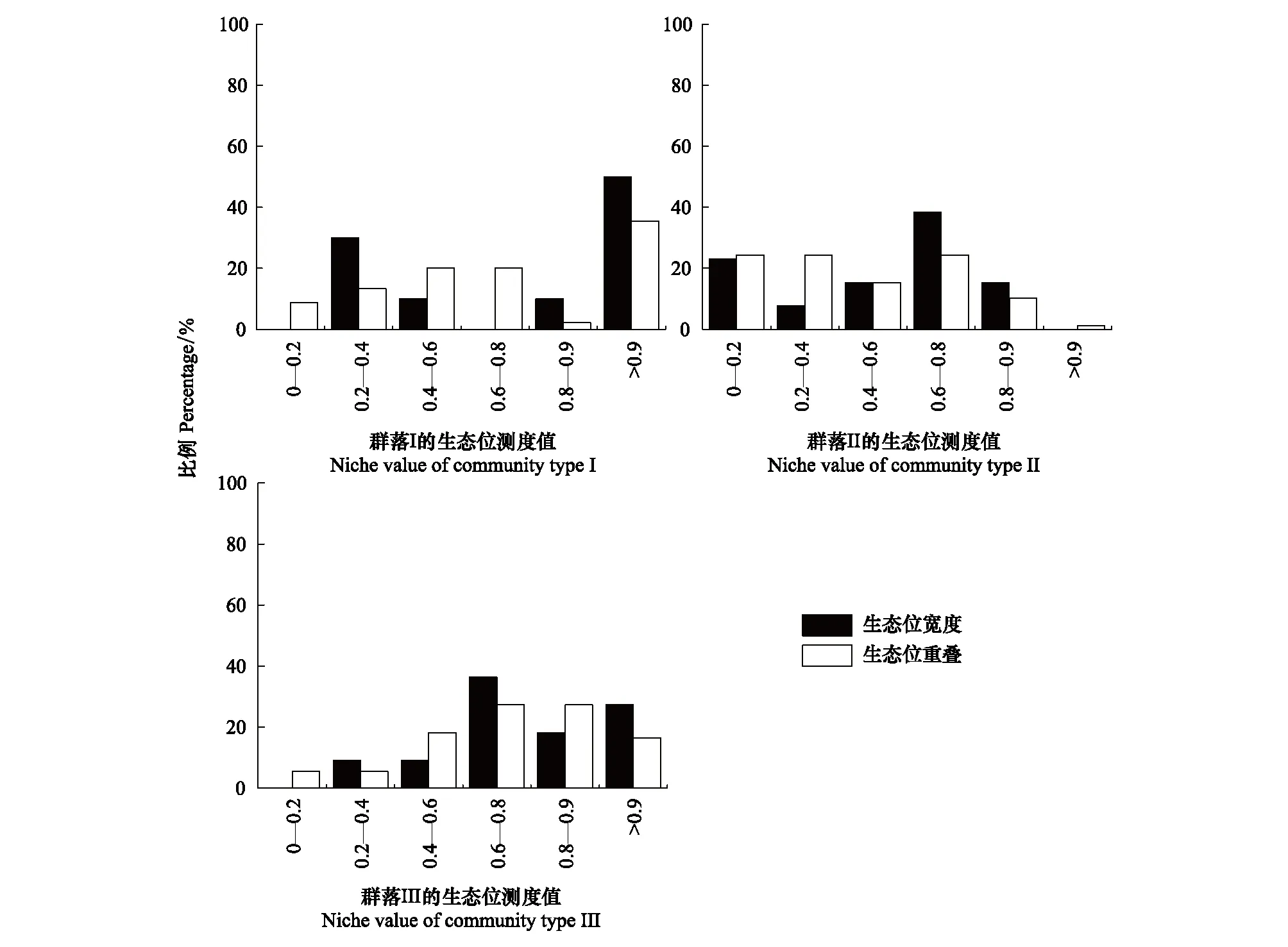
圖2 研究區不同群落下物種生態位測度值的分布格局Fig.2 Distribution pattern of species niche values in different communities in the study areaI:高水鹽土壤環境的群落I,Community I of the high saltwater soil environments;II:中水鹽土壤環境的群落II,Community II of the moderate saltwater soil environments;III:低水鹽土壤環境的群落III,Community III of the low saltwater soil environments
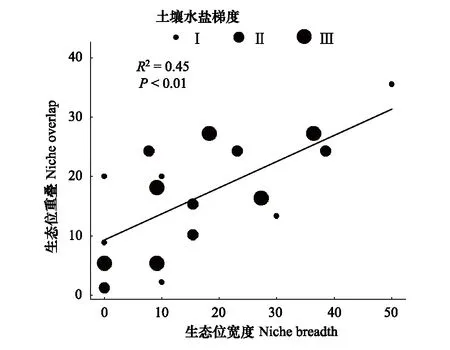
圖3 群落水平上物種生態位測度值的相關關系 Fig.3 Correlation of species niche values at community level
2.3 群落生態位對水鹽的響應特征
根據變系數模型分析群落生態位對水鹽響應的結果如圖4所示,其中圖4,分別為模型(1)的參數項[α(Ui)]和截距項[b(Ui)]的擬合結果,分析表明,當土壤水分小于6.06%時,生態位鹽效應指數[α(Ui)]小于0,說明鹽分對群落生態位是呈現負效應的,反之,土壤水分大于6.06%時,生態位鹽效應指數大于0,說明鹽分對群落生態位是呈現正效應的,而當土壤水分等于6.06%時,生態位鹽效應指數等于0(圖4);如果不考慮鹽分對生態位的效應時,那么群落生態位隨著土壤水分的減少呈現降低的趨勢(圖4)。圖4,分別為模型(4)的參數項[β(Xj)]和截距項[b(Xj)]的擬合結果,分析表明,當土壤鹽分小于4.87 g/kg時,生態位水效應指數[β(Xj)]小于0,說明水分對群落生態位是呈現負效應的,反之,土壤鹽分大于4.87 g/kg時,生態位水效應指數大于0,說明水分對群落生態位是呈現正效應的,而當土壤鹽分等于4.87 g/kg時,生態位水效應指數等于0(圖4);在不考慮水分對生態位效應時,群落生態位隨著土壤鹽分的減少呈現降低的趨勢(圖4)。總的來說,在高水高鹽的土壤水鹽分配模式下,群落的生態位受水鹽的協同作用,主要表現為升高趨勢;在低水低鹽的土壤水鹽分配模式下,群落的生態位受水鹽的協同影響,主要表現為降低趨勢。

圖4 群落生態位對水鹽的響應Fig.4 Response of community niche to water and salt
3 討論
3.1 群叢分類及其決定因素
植物群落是植物在生存環境的長期變化中通過自身的調節機制和適應策略,而聚集在相同地段上的各物種種群的復合體形成的,是維持生態系統物質循環和能量流動的基礎與動力[32]。根據艾比湖地區土壤水鹽特征,將28個植被樣地劃分為3個主要植物群叢,其物種類型的組成(I:10種-II:13種-III:11種)沿水鹽環境因子梯度呈現倒“V”型,這與Kuglerová等[33]的研究結果不一致,Kuglerová等認為,河岸帶的土壤水分條件最好,其植物組成應該最為豐富;Zhu等[34]和Sara等[35]也有相似的結論,認為地下水埋深是制約群落類型、植物分布格局的主要因子,物種的豐富度與地下水位的埋深呈高度正相關。對于本研究區群落的環境特征,因艾比湖研究區處于準噶爾盆地西南緣洼地和水鹽的匯聚中心,I型群叢距離阿其克蘇河河岸較近,土壤獲得河流的補給水分相對較多,地下水位高,而在干旱的環境下,鹽分隨著水分的蒸散運移至土壤表層,淺層土壤鹽分集聚,形成高水高鹽的水鹽空間分布模式,致使植物群落面臨鹽堿脅迫,這對深根系植物胡楊和梭梭的生長有益,對不耐鹽堿的淺根系植物不利[36],因此,河岸邊呈現出以胡楊占絕對優勢的胡楊群落,植物群落的物種豐富度較低;隨著與河流距離的增大,土壤可獲得的補給水量減少,地下水位下降,土壤表層鹽分隨之降低,根系較潛的鹽爪爪、鹽節木和花花柴等鹽生小灌木和草本植物出現,其物種豐富度達到最大,在沿河梯度約1—2.5 km的區域內,土壤適宜的水分條件與較低的鹽漬環境,維持著較高的物種多樣性;進一步加大與河道的距離,地下水位持續下降,導致土壤含水量明顯減少,不足以維持較高的物種豐富度[32-35]。
3.2 不同水鹽梯度群落下種群的生態位寬度和生態位重疊
生態位寬度可以描述種群在群落中所利用資源總和的能力,其大小取決于環境因子的分配模式、物種對環境的生存力和種間的競爭強度。生態位寬度值大的物種,對環境的適應能力越強,越能充分地利用各種資源,且在群落中往往處于優勢地位[2- 4]。在植物群落I類型下,土壤表層屬于高水高鹽的水鹽空間分布模式,利于胡楊、琵琶柴和梭梭等深根系植物的生長,具有較高的生態位寬度值,但不利于檉柳、駱駝刺和甘草等不耐鹽堿的淺根系植物生長,其生態位寬度值較低;植物群落II類型下,土壤水鹽有所降低,一些淺根系耐鹽堿植物開始生長,比如鹽爪爪和鹽節木,但其生態位寬度并非是最大,而白刺和琵琶柴作為研究區內鹽生灌叢的建群種,分布廣泛,具有一定的優勢地位,生態位寬度較大,鈴鐺刺和甘草的生態位寬度均比較小,這說明兩者分布范圍比較窄,對生境的要求具有選擇性;III類型植物群落下,土壤水鹽含量明顯減少,梭梭、花花柴和琵琶柴等旱生植物占取該群落的主要地位,其生態位寬度均較大,而鹽生植物鹽節木的生態位寬度較低,這說明該群落土壤的水鹽環境不適合鹽節木的生存。以上表明,不同的土壤水鹽環境決定了物種的生態位寬度,反映了種群資源利用狀況對水鹽梯度的響應,表明各物種在群落中的地位,進一步明確了不同水鹽梯度下植物群落的結構組成[2, 8- 9]。另一方面,本研究區種群的生態位寬度除了由不同水鹽梯度下植物群落的結構組成決定以外,物種不同的生物學和生態學特性也是一個很重要的因素[12-14],如琵琶柴在3種水鹽梯度下均具有較高的生態位寬度,說明其在該地區具有特殊的適應機制,葉片屬于肉質短小的圓柱形,能夠有效的儲存水分,以及阻止水分散失,同時琵琶柴具有基生葉稠密、根系發達和簇生等特點[37],可以充分利用環境資源,在各水鹽梯度下占取群落中的優勢地位;而甘草以無性繁殖為主,依靠其根莖斷裂脫離母體而形成,決定了對生境的要求具有一定的苛刻性,繁殖與生長受到限制,分布范圍較窄,在3種水鹽梯度下的生態位寬度均較低[38]。
生態位重疊反映兩個或兩個以上生態位相似的物種對同一空間資源利用和競爭的力度[3,5]。對環境資源利用能力較強的物種,與其他物種往往擁有較高的生態位重疊,但是生態位相似的物種間,資源豐富時可能只是占據相近的生態空間,在生態系統學中的功能和特性不一定相同,對同一空間資源的需求類別存在一定差異,不一定會導致種間競爭[2, 7- 8]。本研究發現,在物種水平上,生態位寬度大的物種間的生態位重疊均較大,而生態位寬度較小的物種間的生態位重疊不一定小,如I類型群落的檉柳和駱駝刺,生態位重疊值為1,這在Sandeep等[39]的研究中也有相同的結果,可能存在著兩種原因,一方面物種對生境條件和資源類型的重疊部分較大,在對資源共享的同時也存在競爭關系;另一方面物種在生態系統中的地位需要彼此共同促進而決定其生態學作用和意義。而艾比湖流域屬溫帶荒漠干旱區,資源相對貧缺,具有優勢的物種并不會主動割舍資源生態空間,這樣在長期適應干旱鹽漬化生境過程中,大部分物種對資源匱乏的需求趨于特化而發生趨同適應,生態位重疊普通較高,導致物種間資源競爭激烈,限制了物種共存并驅使群落演替,而群落結構和功能的自身穩定性,反過來又作用于各物種,使其各自扮演著群落中不可或缺的生物生態角色[5- 6]。另外,綜合生態位寬度和生態位重疊值的分析可見,群落水平上種群的生態位寬度與生態位重疊具有一定的相關關系,群落類型I和III物種的生態位寬度較大,生態位重疊較高,說明在這2種土壤水鹽的環境下,種群間存在較強的競爭關系。群落類型II中物種的數量最多,但是個別物種出現的頻率較低,整體上種群的生態位寬度較小,生態位重疊普通較低,物種間的競爭關系相對較弱。這與物種水平上的結果不太一致,可能是因為在資源匱乏的干旱區出現了生境相似的特化種或者伴生種,這些物種生態位寬度值較低,但由于生長或生存的資源緯度相似,物種間生態位重疊值較大,說明生態位寬度和生態位重疊值由物種向群落尺度轉換時,兩者之間的關系存在冗余,尤其是在生態位重疊值較低時,生態位寬度的冗余效應較大。
3.3 群落生態位對水鹽的響應特征
群落生態位寬度可以表示植物群落整體水平對環境資源利用狀況尺度的大小,能夠反映群落中物種的分布狀況,以及間接地衡量群落結構的穩定性,在一定程度上,數值越高表示群落中各物種分布越均勻,該群落的結構越穩定[6]。不同的生境可能會引起群落生態位的變化[7],以往研究表明,自然環境壓力會逐漸淘汰掉一些物種,群落的物種豐富度會降低,并且泛化種的數量也會因為生境壓力逐漸減少,導致群落的結構組成單一化,那么群落整體的資源利用效率和環境適應能力就會減弱,從而限制群落的生態位幅[6-8]。與之相反,也有學者認為,自然環境壓力雖然會對植物進行優劣篩選,但是植物普通的會采用本身的生態學特性、共生或特化等策略保持個體的數量并均勻地分布,提高自身在群落中的生態空間地位[12],群落整體的資源利用能力和環境適應能力會增強,促進群落生態位寬度的增幅[40]。但在該過程中,另有觀點認為,在干旱區荒漠基質植物群落中,資源匱乏導致物種間的競爭關系加強,整體群落的生態幅并不會有太大的改變,極端情況下可能會發生抑制作用[4,41]。本研究發現,高水高鹽的土壤水鹽分配格局對群落的生態位寬度有一定的促進效應,而低水低鹽的土壤水鹽分配模式對群落的生態位寬度具有一定的限制作用,土壤的水鹽環境梯度與群落生態幅呈現一種非線性相關特征,這一結論與上述觀點相悖。這可能是因為,近河岸荒漠森林生態系統的資源充足,植物分布均勻,并且能夠有效地提高群落中各物種的數量,加強群落的穩定性,群落對資源的利用能力和效率也會有效增加,從而群落生態幅增大[24,40,42],相反,遠離河岸的低水低鹽土壤環境導致植物分布不均勻,并且群落中各物種的個體數量無法持續維持,群落結構組成單一,穩定性較差,對資源利用的能力和效率整體降低,從而群落生態幅減小[8,35]。另一方面,研究區鹽漬化以及干旱缺水的天然缺陷,使水分補給量成為植物生長和群落演替的關鍵因素,而水分的補給來源和效率在維持植物數量和群落穩定方面可能起著重要作用,近河岸土壤具有高水高鹽的分配特征,擁有長期持續性的河水的補給占比較大,容易被植物群落吸收利用,同時水分補給效率也較高,而遠離河岸低水低鹽的土壤環境,深層地下水的補給方式增加了植物地下的根投入比,其水分補給效率較低[35,39-41],生境間水源可獲取的多樣性,影響著植物地下和地上的形態結構,進一步改變了群落的生態結構,這個可能是決定研究區植物生態位寬度的重要因素,將成為今后研究的重點。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
保健醫苑(2021年7期)2021-08-13 08:48:02
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
