典型高寒植物有性繁殖對長期增溫的響應
2019-05-31 03:33:26康瀕月徐恒康姜風巖邵新慶
草地學報 2019年2期
康瀕月,徐恒康,姜風巖,邵新慶
(中國農業大學草業科學與技術學院,北京 100193)
全球氣候對陸地生態系統的影響及其反饋是全球變化與陸地生態系統(Global Change and Terrestrial Ecosystem,GCTE)研究的熱點,據大氣環流模型(General Circulation Model,GCM)預測,21世紀全球溫度將升高1.5℃~4.5℃[1]。這種全球氣候變暖趨勢不但會影響植物生理生態特征,也會對植物種群、群落、生態系統乃至整個生物圈產生巨大影響。尤其在高緯度和高海拔的青藏高原,生態環境脆弱[2-3],植物對溫度升高的響應非常敏感[4-5]。
有性繁殖是植物繁衍后代的主要方式之一,有利于植物對當地環境的適應,維持遺傳多樣性[5]。研究表明,有性生殖階段對環境的變化最敏感[6]。隨著全球氣候變化,青藏高原氣候呈現出暖化趨勢,溫度升高可能對當地植物生殖生長過程的授粉受精、生殖產量產生明顯影響[7]。在研究增溫對植物有性生殖的影響中,一些研究者得出了相似的結論:Benlloch-Gonzalez等[8]發現溫度升高4℃會降低果實產量并產生較小的果實,也有研究[9-10]表明增溫顯著降低了短花針茅結實率、種子百粒重和發芽率。但不同研究者通過模擬增溫對不同草地生態系統的影響的研究得到的結論不盡相同。曹素珍認為[11]增溫降低植物的繁殖投入,促進營養生長,Klady等[12]認為變暖促進了大多數物種的繁殖生長,并且北極地區長期變暖可能會促進苔原植物的有性繁殖;也有研究認為增溫對禾草科和雜類草的生長沒有顯著影響[13]。研究植物繁殖特征對增溫的響應,是研究氣候變化背景下高寒草甸群落結構演替和功能變化的基礎。目前鮮有對植物有性繁殖對增溫響應及敏感性的研究,有關這一方面的大部分研究結論大多基于短期和室內控制試驗,難以獲取高寒植物響應氣候變化的綜合信息,更難以得出響應敏感性與趨勢的結論。因此,本研究在青藏高原海北試驗站使用開頂式生長室(Open-Top Chambers,OTCs)進行長期和室外控制模擬增溫試驗[14-16],以高寒典型植物莎草科矮生嵩草(Kobresiahumilis)、青藏苔草(Carexmoorcroftii)和禾本科垂穗披堿草(Elymusnutans)為研究對象,研究增溫處理下這3種植物的結實率、種子產量、種子活力等繁殖特征對增溫的響應,分析3種植物的生殖策略,為預測高寒草甸草原植物群落變化趨勢以及科學評估氣候變化對高寒草甸的影響奠定了數據和理論基礎。
1 材料與方法
1.1 研究區概況
本試驗在海北高原現代生態畜牧業科技試驗示范園進行,該園區位于青海省海北藏族自治州海晏縣西海鎮,地處青藏高原東北部青海湖東岸(36°55' N,100°57' E,海拔3029 m)。該地區屬于高原大陸性季風氣候,僅有冷暖兩季之分。年均溫1.5℃左右,年日照時數2980 h左右,相對濕度為45%~60%,年降雨量約400 mm,降雨主要集中在7-9月。土壤類型為高山草甸土,土層較薄,缺磷,少氮。0~20 cm土壤有機質含量為27.2 g·kg-1,全氮含量2.5 g·kg-1,有效磷含量為0.05 g·kg-1。該區域植被類型為高寒針茅化草甸草,主要優勢種為紫花針茅(Stipapurpurea)、草地早熟禾(PoapratensisL.)、冷地早熟禾(Poacrymophil)、垂穗披堿草、線葉嵩草(Kobresiacapillifolia)、矮嵩草、扁蓿豆(Medicagoruthenica)和二裂委陵菜(Potentillabifurca)等。
1.2 試驗設計
本試驗研究對象為自然狀態下的高寒草原化草甸生態系統,自2012年起采用開頂箱(OTCs)式增溫裝置(圖1)對試驗樣地進行長期增溫。根據國際凍原計劃(International Tundra Experiment,ITEX)標準建立模擬增溫的野外試驗平臺,試驗樣地地勢平坦,植物分布均勻,用圍欄封閉。試驗處理分增溫(Warming,W)和對照(Control,CK),各8組重復,增溫采用OTCs增溫裝置進行增溫2℃,并設計自動控溫系統和抽氣式風扇,當OTCs內外溫差值超過2℃時,風扇自動啟動,保證增溫不超過設定值,維持溫度穩定。OTCs永久性安裝于樣地里,規格為:底部直徑2.2 m,頂部直徑1.5 m,高度0.7 m。在7-8月對試驗對象:矮生嵩草、青藏苔草和垂穗披堿草進行調查,并在各自結實期進行取樣。

圖1 開頂式生長室示意圖Fig.1 The sketch of the open top chamber
1.3 試驗方法
1.3.1土壤溫度測定方法 使用AR5數據采集器采集每個樣地地上20 cm處的空氣溫度、濕度和地下5 cm處的土壤溫度、濕度(15 min一次)。
1.3.2調查植物種群數量特征測定方法 在草地植物旺盛生長期(7-8月),采用方精云等[19]介紹的樣方法對垂穗披堿草、青藏苔草和矮蒿草的種群數量特征進行調查,主要調查指標為相對密度和相對蓋度。3種植物種群特征情況見表1。

表1 增溫下3種植物種群數量特征Table.1 Population characteristics of three plant species under warming
注:CK:空白對照;W:增溫
Note:CK:Control;W:Warming
1.3.3有性繁殖指標測定方法 采用周興民等[17]介紹的方法,測定草甸植被主要種群繁殖對策參數。具體做法為:在每個樣地上布設1 m×1 m樣方,2018年8月在3種代表草本植物:矮生嵩草、青藏苔草、垂穗披堿草的各自果期分別對其繁殖枝和營養枝進行計數并采用全收獲法[18]收獲,將根和根狀莖上的泥土洗凈、晾干,并將植物構件分為繁殖器官(花、花軸或花莖)和非繁殖器官(莖、葉、根等)和果實三部分,同時記錄繁殖枝上的小穗及種子數目。于80 ℃恒溫下將植物烘干至恒重(約需要48 h)[19]。用電子天平精確稱重各部分質量(精確在0.0001 g),計算有性繁殖效力。有性繁殖效力用穗(包括花和種子)占總生物量(以干重計)的比例來計算。
1.3.4種子生產能力測定方法 在采樣過程中基于繁殖枝是否萌發將所有樣本分為繁殖性和非繁殖性[20]。對于每株繁殖性植物,對其繁殖枝數進行統計,然后使用枝數來估計每株植物(i)每年(y)產生的種子數量(Seed,S),關系如下:
Si,y=Ti,y×f×s
注:T:繁殖枝數;f:每根繁殖枝上果實數;s:每個果實產生的種子數量
1.3.5種子發芽率測定方法 參考鄧自發等[21]介紹的方法采集種子,在各自結實期結束時,在完全隨機設計的每個樣地內隨機采集3種調查植物繁殖枝25枝,逐穗收集種子,對有成熟種子的穗和每穗種子數進行計數。同時為避免OTCs氣室的邊緣效應,在其中心取樣。對樣品種子依據完全隨機設計試驗的樣地順序進行編號并按照標準發芽試驗步驟進行室內發芽,采用紙上法在恒溫條件下發芽,從每個樣地內3種調查植物種子中分別取50粒、4組重復,置于培養皿中潤濕的濾紙上(2層),加皿蓋在25℃光照處理12 h,暗處理12 h)恒溫培養箱中萌發。由于苔草種子的休眠特
性,提前將青藏苔草種子進行低濕層積14 d[22]。萌發期間,保持適宜水分,統計發芽勢、發芽率,以胚根突破種皮1 mm或以上計為發芽[23]。14 d結束實驗。統計試驗末期發芽種子占供試種子的百分數。
發芽率(%)=種子總發芽數/供試種子數×100%
發芽勢(%)=規定時間內種子發芽數/供試種子數×100%
1.4 數據分析
采用Excel 2016進行數據整理、計算和作圖,數值均以平均值±標準誤表示。利用IBM SPSS Statistics 23軟件進行單因素方差分析(One-way ANOVA),用LSD法在顯著性水平0.05和0.01下進行差異性檢驗,不同大寫字母表示在增溫處理下差異極顯著(P<0.01),不同小寫字母表示增溫處理下差異顯著(P<0.05)。用R語言3.4.4對3種植物繁殖枝數量、小穗長度、繁殖效力、種子產量、發芽勢和發芽率的數據進行heatmap分析并繪制圖像。
2 結果分析
2.1 繁殖枝數量、小穗長度和繁殖效力的變化
由圖2可知,增溫處理下,3種植物繁殖枝數量都有減少的趨勢,小穗長度有增加的趨勢。其中,矮嵩草繁殖枝數量在增溫下極顯著降低(P<0.01),平均每株矮嵩草減少1~2枝繁殖枝,其小穗長度的變化趨勢與繁殖枝數量變化趨勢基本相反(圖2),有增長趨勢但是與對照之間差異不顯著。從圖2可見,增溫對青藏苔草、垂穗披堿草有效穗長度有顯著影響(P<0.05),其中垂穗披堿草有效穗長度在增溫處理下平均每株增加2~3 cm,青藏苔草有效穗長度在增溫處理下平均每株增加0.5~1 cm。說明在增溫影響下,3種調查植物繁殖枝數量與其有效穗長度變化方向相反,且變化幅度不一致。
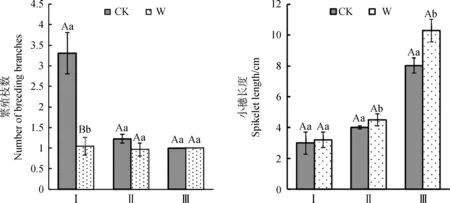
圖2 增溫對高寒植物繁殖莖數量、小穗長度的影響Fig.2 Effects of warming on the numbers of stems and spikelets of alpine plants注:Ⅰ:矮嵩草;Ⅱ:青藏苔草;Ⅲ:垂穗披堿草。下同Note:Ⅰ:Kobresia humilis;Ⅱ:Carex moorcroftii;Ⅲ:Elymus nutans. The same as below

圖3 增溫對有性繁殖效力的影響Fig.3 Effect of warming on sexual reproduction efficiency
由圖3所示,總體上,矮嵩草有性繁殖效力在增溫處理下平均每株減少2%~3%,其變化趨勢與繁殖枝數變化趨勢基本一致,青藏苔草與垂穗披堿草有性繁殖效力呈下降趨勢并與其小穗長度變化趨勢基本一致;其中矮嵩草增溫下有性繁殖效力極顯著降低(P<0.01),而青藏苔草、垂穗披堿草有性繁殖效力無顯著變化(P>0.05)。以上結果說明在增溫處理下,矮嵩草的繁殖枝數、有性繁殖效力受影響程度較大,與增溫呈負響應;青藏苔草、垂穗披堿草的有效穗長度受影響程度較大,且與增溫呈正響應。
2.2 種子生產能力的變化
從種子生產能力的角度來看,總體上,增溫會引起植物種子生產能力降低(圖4)。其中,增溫引起矮嵩草植物種子生產能力極顯著降低(P<0.01),平均每株矮嵩草所產種子在增溫下減少5~6粒;增溫引起垂穗披堿草種子生產能力顯著降低(P<0.05),平均每株垂穗披堿草所產種子減少13~16粒;青藏苔草種子產量沒有顯著差異。
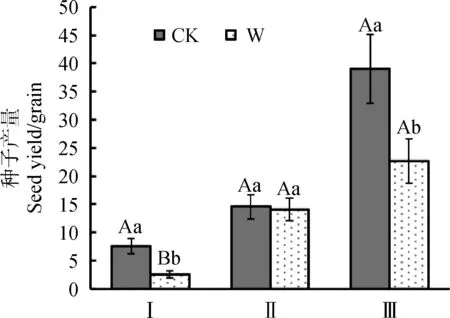
圖4 增溫對植物種子產量的影響Fig.4 Effect of warming on seed yield
2.3 發芽勢與發芽率的變化
在增溫處理下,3種植物種子室內發芽勢都降低(圖5)。其中矮嵩草、垂穗披堿草室內發芽勢在增溫下極顯著降低(P<0.01)。矮嵩草種子發芽勢從63.5%降低至22.0%,垂穗披堿草種子發芽勢從25.1%降低至6.8%,青藏苔草室內發芽勢在增溫下顯著降低(P<0.05),從6.9%降低至3.6%。3種植物種子室內發芽率在增溫下都無顯著變化。該結果說明增溫處理會降低調查植物種子發芽勢,使發芽速度降低,但對其發芽率無明顯影響。
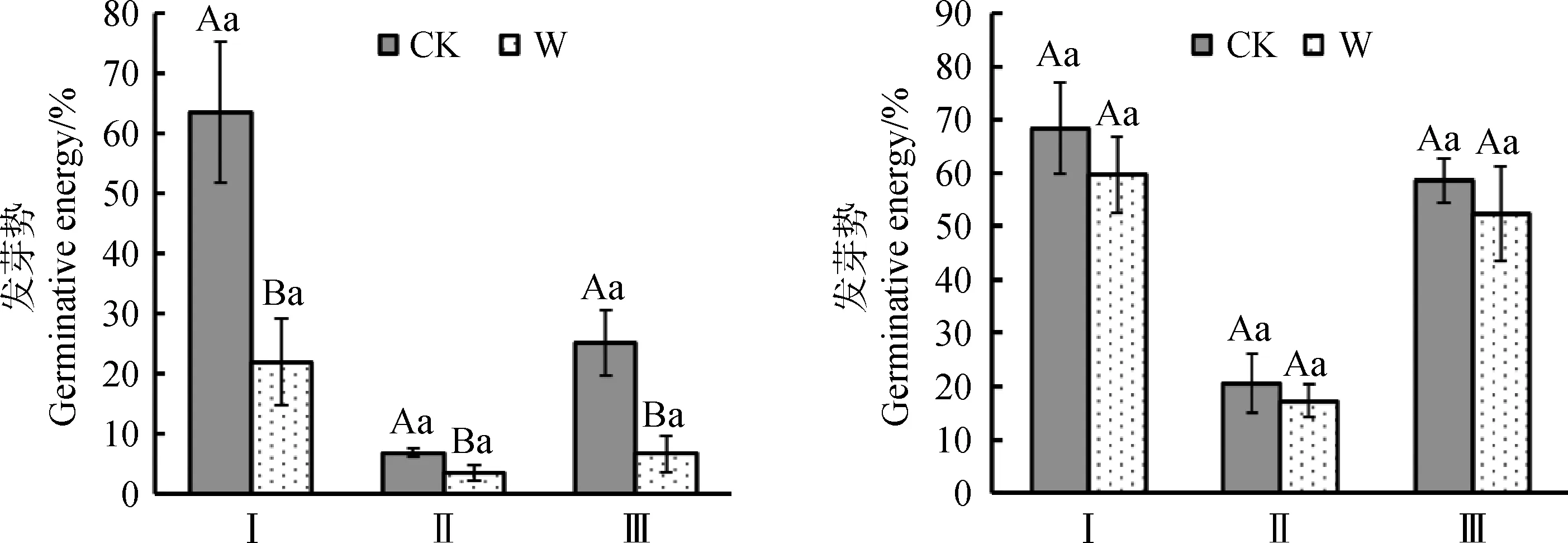
圖5 增溫對發芽勢與發芽率的影響Fig.5 Effect of warming on germination energy and germination rate
2.4 3種植物有性繁殖對增溫的響應敏感性分析
根據3種植物有性繁殖指標對增溫的響應進行Heatmap作圖分析。敏感程度用一定顏色梯度表示(圖6中左側為顏色梯度代表數值),顏色越深代表在增溫下該植物的指標越敏感,表示該植物的指標相較其他兩種植物對增溫的響應程度越大,用可視化的圖形綜合反應2.1~2.3的結果。
由圖6可知,增溫下,矮嵩草繁殖枝數、有性繁殖力、發芽勢和發芽率指標響應敏感,且幅度較大,青藏苔草的發芽勢與發芽率響應敏感;垂穗披堿草與矮嵩草和青藏苔草相比,其小穗長度與種子產量響應敏感,其余不明顯。綜上,在增溫影響下各指標對增溫效應響應敏感性最大的是矮嵩草,響應敏感性較大的為青藏苔草,響應敏感性最弱的為垂穗披堿草。

圖6 植物有性繁殖對溫度響應的敏感性Fig.6 Sensitivity of the response of plant sexual reproduction to temperature注:NBB:繁殖枝數;SRE:有性繁殖力;GE:發芽勢;GR:發芽率;SL:小穗長度;SY:種子產量Note:NBB:number of breeding branches;SRE:sexual reproduction efficiency;GE:germination energy;GR:germination rate;SL:spikelet length;SY:seed yield
3 討論
3.1 增溫對3種高寒植物有性繁殖的影響分析
環境因子對植物的有性繁殖特征影響強烈,因此在不同環境因子作用下,草地植物會有很大的形態特征改變甚至會影響植被組成[24]。本研究中利用OTCs氣室模擬長期增溫探究對3種調查植物有性繁殖的影響,結果表明,增溫會使其生殖枝減少而使小穗長度增加,這與王志保等[25]的研究結果一致。由于研究對象的物侯期同樣受溫度影響[26],長期觀測與增溫試驗發現:隨著持續增溫,植物返青期或初花期顯著提前,秋季枯黃期顯著延遲[26],物侯期的變動影響收取種子時間的準確性,若按照原有果實期來進行種子的取樣,可能會造成對種子產量、發芽率、發芽勢的低估,從而影響試驗結果的正確性。但Jiang等[28]在青藏高原研究發現,與其他物候期相比,果期物候對增溫和降溫的響應保持相對穩定性,這可能是植物維持種子成熟以及傳播成功的機制,基于此結論可以確保野外調查時間的準確性。本研究發現,盡管青藏苔草和垂穗披堿草的小穗長度在增溫下增加,但種子產量降低,這說明盡管小花數量變多但是結實率反而降低,因此種子產量降低。從結實率低的原因來考慮,由于禾本科和莎草科植物大部分都是風媒花,需要借助風力進行傳粉完成受精,且OTCs的“花房效應”限制了水平方向的空氣流動,在一定程度上降低了垂直方向的對流過程,室內空氣湍流作用減弱[29],極有可能影響了低矮植物矮嵩草、青藏苔草的傳粉受精,但垂穗披堿草長勢高,可能不受OTCs氣室遮擋的影響,能保證傳粉自由進行,但依然表現出增溫后種子產量降低的效應,這說明增溫的確會使植物結實率降低,從而導致其種子產量減少。
植物的有性繁殖階段容易受到氣候變暖的影響[30],且氣候變暖可以改變植物的有性繁殖物候,從而大大降低密度依賴和某些植物物種的有性繁殖能力[31],這能很好地解釋本研究中矮嵩草在增溫處理下各項指標都降低的現象,表明有性繁殖能力被抑制。但也有一些研究表明增溫促進植物有性繁殖[29],原因可能是由于增溫時間的不同或繁殖對策不同。本研究中,增溫后3種植物種子室內發芽勢降低,原因可能是在形成種子的過程中,由于增溫的影響,加快了土壤水分蒸發速率,植物受到干旱脅迫,在不良環境下易形成休眠種子以渡過不良環境,有利于其種質的延續[32],說明發芽勢的降低可能是植物維持種子活力的一種繁殖對策。趙玉紅等[33]認為發芽率無明顯變化說明種子發芽能力未受損,這就很好解釋本研究中增溫處理未使種子發芽率明顯降低的現象,揭示增溫下植物維持種子發芽能力的對策。
綜上所述,溫度升高一方面將影響土壤含水量及結構,另一方面將影響植物對養分與水分的吸收以及有性繁殖[34],因此推測短期增溫促進植物的有性繁殖,但長期增溫抑制植物有性繁殖,其原因可能是土壤類型發生改變[29],進而影響高寒草甸植被群落組成和結構,影響群落變化的方向,根據“生草土形成理論”可預測最后會發生植被的演替[32]。
3.2 3種植物有性繁殖對增溫的響應敏感性分析
最新數據顯示:溫度升高使植物有性繁殖過程對氣候變化的敏感性增強[30]。不同植物對于溫度的敏感程度不同[35]。趙建中等[36]認為不同功能群植物種對地表溫度升高的響應是不一致的,地表溫度持續升高對禾本科功能群植物種的生長有利,而對莎草科、雜類草功能群物種均不利。這與龐曉瑜[37]研究結論一致,即增溫促進禾本科生長,抑制莎草科生長,這能很好解釋本研究中矮嵩草的有性繁殖對溫度的變化最為敏感,且基本上為負響應,而青藏苔草有性繁殖對溫度的響應程度較高,垂穗披堿草有性繁殖對增溫的響應程度最低的現象(圖6),且對莎草科物種來說不同植物響應程度也是不一致的。從群落中種群的空間分布來考慮,垂穗披堿草處于群落上層,對底層莎草有遮蔽作用,極有可能造成不同功能群之間有性繁殖特征的差異,但是同屬莎草科且長勢低的矮嵩草與青藏苔草的有性繁殖對溫度響應程度也各有不同,因此可以排除垂穗披堿草的遮蔽作用帶來的影響,把原因歸結為各植物間繁殖對策的差異,使得有性繁殖對溫度的響應各有不同,這也就能解釋為何增溫后青藏苔草與垂穗披堿草的有性繁殖效力無顯著變化(圖3)。垂穗披堿草的有性繁殖增溫下并未明顯表示出正響應,因此,若要探究垂穗披堿草是否會取締矮嵩草成為優勢種還需結合無性繁殖進行系統分析。
4 結論
在增溫處理下,不同植物表現出不同的繁殖策略,矮嵩草的繁殖枝數量、有性繁殖效力極顯著降低(P<0.01)。青藏苔草表現為有效穗長度顯著增加和種子產量顯著降低的趨勢,垂穗披堿草小穗長度極顯著增加,種子產量顯著降低;增溫使3種植物的發芽勢有一定下降的趨勢,對發芽率影響不明顯。在增溫下,矮嵩草有性繁殖受到抑制,且最為敏感。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
媽媽寶寶(2017年3期)2017-02-21 01:22:28
爆笑show(2016年7期)2017-02-09 09:36:13
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
