AmoA、AmoE和AmoF蛋白在嗜水氣單胞菌鐵載體合成中的作用研究
2019-05-29 06:58:28馮宇晴張倩倩章晉勇張蘭梨馬麗娜馮東岳吳振兵郝靜文王姝懿李愛華
水生生物學報 2019年3期
馮宇晴 張倩倩 章晉勇 張蘭梨 馬麗娜 馮東岳 吳振兵 郝靜文 王姝懿 李愛華
(1. 中國科學院水生生物研究所淡水生態與生物技術國家重點實驗室, 武漢 430072; 2. 中國科學院大學, 北京 100049;3. 吉林大學藥學院, 長春 130000; 4. 全國水產技術推廣總站, 北京 100125)
嗜水氣單胞菌(Aeromonas hydrophila)隸屬于氣單胞菌科氣單胞菌屬, 革蘭氏陰性短桿菌, 廣泛分布于水體環境中[1,2], 是一種典型的人、畜、魚共患病病原菌[3], 該致病菌對淡水養殖魚類危害很大, 給水產養殖業造成了嚴重的經濟損失[4]。嗜水氣單胞菌致病機制十分復雜, 涉及到分泌系統、胞外產物、黏附因子、群體感應系統及鐵螯合系統等多種毒力因子[5]。
鐵作為重要的微量元素, 在生物體的光合作用、呼吸作用、三羧酸循環、氧轉運、基因調控、DNA生物合成等生物過程中發揮重要的作用[6]。然而, 環境中可直接利用的鐵濃度很低, 在有氧和中性pH條件下, 鐵高度不溶, 通常以三價鐵氧化物水合物的形式存在[7], 在動物體內, 鐵與轉鐵蛋白、乳鐵蛋白、鐵蛋白和血紅素蛋白等蛋白緊密結合[8]。因此, 細菌進化出多種復雜的機制從環境和宿主體內攝取鐵, 其中一條主要的途徑是分泌出一種低分子量且結構多樣的鐵螯合劑——鐵載體[9]。鐵載體能夠有效的從三價鐵復合物和鐵結合蛋白中攝取鐵, 然后通過細菌外膜鐵載體受體、周質結合蛋白、ABC轉運系統將三價鐵轉運至胞內, 以維持細菌的正常生長[10]。有研究表明, 嗜水氣單胞菌可同時利用2種機制攝取鐵, 一種是依賴鐵載體的機制, 另一種則是從含有血紅素的蛋白中攝取鐵[11]。
1982年, 國外研究者Andrus等首次報道嗜水氣單胞菌能夠產生兒茶酚類鐵載體, 隨著研究的推進,十幾年后鐵載體的四種結構得到確證, 每種結構中都包含2,3-二羥基苯甲酸和賴氨酸, 苯丙氨酸、色氨酸和甘氨酸的存在與否決定了鐵載體的4種不同結構[12]。嗜水氣單胞菌ATCC 7966全基因組測序完成, 通過基因組比對方法推測出鐵載體合成基因簇AHA2473-AHA2479, 共包含7個基因[13]。Barghouthi等[14]通過轉座子插入失活的方法證實了amoC基因(原文獻中名為amoA, 現命名為amoC)參與鐵載體的合成, 該基因轉錄翻譯為異分支酸合成酶, 參與合成途徑的第一步, 使得分支酸轉變為異分支酸。2016年,amoG、amoH也被證實與鐵載體的合成相關, 其中amoG基因失活導致鐵載體合成缺陷,amoH基因缺失株只能合成2種結構的鐵載體[15]。鐵載體的合成過程十分復雜, 通常需要一套完整的專用基因來編碼所需的生物合成酶。本研究首先用RT-PCR初步了解amoA、amoE, 以及amoF基因在富鐵和低鐵條件下的轉錄水平, 再利用融合PCR和基因同源重組原理, 以自殺性質粒pRE112為載體構建基因缺失株ΔamoA、ΔamoE和ΔamoF, 確認這3個蛋白在嗜水氣單胞菌鐵載體合成途徑中的作用。
1 材料與方法
1.1 菌株及質粒
嗜水氣單胞菌4LNC202由本實驗室于2008年6月在患病鰱魚肝臟中分離保存[16], 具有氨芐青霉素抗性(表1); 大腸桿菌S17-1λpir及pRE112質粒均由中國科學院水生生物研究所謝海俠老師饋贈[17]。

表1 本試驗中使用的菌種和質粒Tab. 1 Strains and plasmids used in this study
1.2 培養基
TSB培養基、10×MM9培養基和CM9培養基。10×MM9培養基和CM9培養基制備方法如下:10×MM9培養基: KH2PO43 g、NaCl 5 g、NH4Cl 10 g、ddH2O 100 mL, 121℃高壓滅菌20min; CM9培養基: Na2HPO46 g、KH2PO43 g、NaCl 0.5 g、NH4Cl 1 g、ddH2O 1000 mL、調節pH=7.4、121℃高壓滅菌20min, 待液體冷卻后依次加入以下無菌物質: 20%葡萄糖10 mL、1 mol/L MgSO42 mL、1 mol/L CaCl20.1 mL、10%去鐵水解酪蛋白20 mL。
1.3 主要的試劑及儀器
DNA Marker購自北京擎科新業生物技術有限公司, 2×Taq MasterMix購自北京康為世紀生物科技有限公司; 細菌基因組提取試劑盒、質粒提取試劑盒購于OMEGA公司; 細菌RNA提取試劑盒購自QIAGEN公司, 一步法無縫克隆試劑盒購自南京諾唯贊生物科技有限公司; 限制性內切酶購自Fermentas公司; PrimeSTAR Max DNA Polymerase、反轉錄試劑盒購自大連TaKaRa公司; 蔗糖、氯霉素、氨芐青霉素購自Sigma公司。PCR儀購于Thermo公司; 核酸電泳儀購于Bio-Rad公司。
1.4 引物的設計與合成
采用生物軟件Primer 5.0設計實時熒光定量PCR引物對(表2)和構建基因缺失株所用引物對(表3), 引物均由北京擎科新業生物技術有限公司合成。
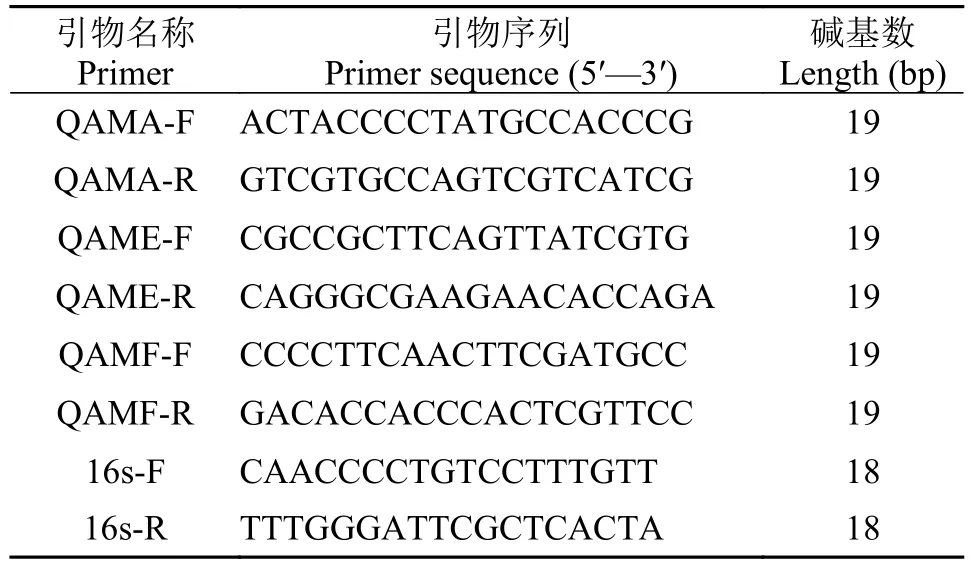
表2 實時熒光定量PCR引物Tab. 2 Primers used in Real-time quantitative PCR
1.5 在富鐵、低鐵條件下amoA、amoE、amoF基因轉錄水平檢測
嗜水氣單胞菌4LNC202劃線接種于TSA平板,28℃培養過夜。挑單菌落分別接種于TSB培養基和CM9培養基中, 在28℃、180 r/min條件下培養至對數期。參照QIAGEN RNA提取試劑盒說明書抽提細菌RNA, 1%瓊脂糖凝膠電泳檢測RNA質量, 超微量分光光度計NanoDrop2000測定RNA濃度。取1 μg RNA進行反轉錄, 具體步驟參考試劑盒說明書, 反轉錄后的cDNA于-20℃保存備用。
選取穩定表達的16S rRNA基因作內參, RTPCR檢測amoA、amoE、amoF基因在富鐵和低鐵條件下表達量的變化。
20 μL PCR反應體系如下: SYBR GreenⅠMix 10 μL, 上、下游引物(10 μmol/L)各0.8 μL, 模板cDNA 1 μL, ddH2O 7.4 μL。
PCR反應條件為: 95℃預變性2min; 95℃變性10s, 58℃退火30s, 72℃延伸20s, 39個循環。

表3 構建基因缺失突變株所用引物Tab. 3 The primers used to construct the gene deletion mutant strain
1.6 基因缺失突變株的構建與鑒定
根據Ho等[18]的方法構建基因缺失株。 以菌株4LNC202全基因組DNA為模板, 用引物P1/P2 和P3/P4 分別擴增目的基因上、下游同源臂。通過融合PCR的方法, 以上、下游同源臂為模板, 使用P1/P4 引物, 融合上下游同源臂。
用限制性內切酶XbaⅠ和NdeⅠ將pRE112線性化, 根據一步法無縫克隆試劑盒說明書將線性化pRE112質粒與融合上下游同源臂進行連接。方法簡述如下: 20 μL反應體系包含5×CE Ⅱ Buffer 4 μL、Exnase Ⅱ 2 μL、線性化PRE112 1 μL、融合上下游同源臂1 μL、ddH2O 12 μL。37℃條件下反應30min, 反應結束后立即將離心管置于冰上。取上述10 μL連接產物轉化至大腸桿菌S17-1λpir感受態細胞中, 將培養物涂布TSA+Cm平板上, 篩選陽性克隆并進行菌落PCR鑒定。鑒定后的陽性質粒送北京擎科新業生物技術有限公司測序。
將含有重組質粒的大腸桿菌S17-1λpir作為供體菌, 以嗜水氣單胞菌4LNC202 為受體菌, 通過接合轉移的方式將重組質粒從大腸桿菌S17-1λpir轉移到嗜水氣單胞菌4LNC202 中, 經2次同源重組得到基因缺失突變株。具體操作如下:
(1)接合。供體菌S17-1λpir和受體菌4LNC202分別培養至OD600達到0.5, S17-1λpir和4LNC202按照2∶1比例混勻, 取100 μL轉移至新鮮濕潤的TSA平板中央, 28℃ 靜置培養24h; 用PBS沖洗TSA平板,取200 μL菌液涂布于TSA+Cm+AmP平板, 28℃培養24—48h。以P1/P4為引物做菌落PCR篩選接合成功的重組子, 此時重組質粒已整合到嗜水氣單胞菌4LNC202 基因組上。
(2)二次交換及缺失株篩選。選取成功結合的重組子, 接種至 5 mL TSB液體培養基中, 28℃靜置培養24h, 進行二次交換。然后將菌液梯度稀釋涂布于TSA+Amp+10%蔗糖的平板上, 28℃培養至長出單個菌落, 挑取單克隆同時劃線接種于TSA+Cm+Amp、TSA+Amp兩類平板, 28℃培養 24h。選擇只在TSA+Amp平板生長的單克隆, 劃線接種于TSA+Cm平板, 如若不生長, 則可能是突變株或是恢復到野生型的菌株。以P1/P4為引物PCR檢測,選擇產生正確目的條帶的單克隆保種。
1.7 RT-PCR鑒定基因缺失突變株
RNA與cDNA的制備方法、RT-PCR的反應體系和反應條件同1.5, PCR反應結束后, 取5 μL擴增產物經1%瓊脂糖凝膠電泳檢測, 凝膠成像系統記錄結果并照相。預期如下: 用短引物對擴增疑似基因缺失突變株ΔamoA、ΔamoE和ΔamoF不能擴增出任何條帶, 而擴增野生株WT獲得的條帶大小分別約為164、141和180 bp。
經上述PCR、RT-PCR篩選到的陽性基因缺失株送北京擎科新業生物技術有限公司測序, 確認嗜水氣單胞菌突變株ΔamoA、ΔamoE和ΔamoF序列是否正確。
1.8 生長特性
不同濃度2,2-聯吡啶作用下野生株與突變株在CM9培養基中的生長實驗嗜水氣單胞菌野生株與突變株菌液劃線接種于TSA平板上, 在28℃恒溫培養箱中過夜培養。挑單菌落接種于TSB液體培養基中, 28℃、180 r/min搖床培養至對數期,PBS洗滌3次后重懸, 調整OD600為0.5, 按照1∶1000(v/v)的比例將菌液接種于含有不同濃度鐵螯合劑2,2-聯吡啶的CM9培養基中, 28℃、180 r/min搖床培養18h, 在600 nm波長下測吸光值, 作為后續實驗2,2-聯吡啶添加濃度的依據。
細菌在富鐵、低鐵培養基中的生長實驗根據上述方法調整野生株突變株OD600為0.5, 按1∶100(v/v)的比例將上述菌液接種于50 mLCM9培養基(2,2-聯吡啶: 50 μmol/L)中, 28℃ 180 r/min振蕩培養, 每種菌體設置3個重復, 每隔1h用分光光度計檢測一次菌液吸光值(OD600), 連續6h, 記錄數據并繪制曲線。
1.9 鐵載體的檢測
微生物鐵載體根據其螯合基團的化學特性進行分類, 一般分為三大類, 即異羥肟酸類鐵載體、兒茶酚類載體、羥基羧酸鹽類鐵載體。Arnow[19]在1937年提出兒茶酚類鐵載體的檢測方法, Arnow法檢測鐵載體的實驗原理是: 含有苯環或者羥基的化合物在酸性條件下與亞硝酸鈉鉬酸鈉溶液混合可以生成一種紅色的物質, 并在510 nm處有最大光吸收值。鐵載體活性檢測采用CAS固體平板檢測,該方法由Schwyn等[20]于1987年提出, CAS實驗檢測鐵載體的原理: 鐵載體能夠與Fe3+形成更穩定的鰲合物, 伴隨著Fe3+從其結合緊密CAS-Fe3+-HTDMA藍色絡合物轉移到鐵載體中發生顏色變化。
Arnow實驗根據1.8的方法調整野生株突變株OD600為0.5, 按1∶1000(v/v)的比例接種于CM9培養基(2,2-聯吡啶: 50 μmol/L), 28℃、180 r/min搖床培養24h, 4000 r/min 4℃離心10min, 吸取上清液用0.22 μm的過濾器進行過濾, 濾液于-20℃冰箱保存, Arnow實驗參照文獻[19]方法檢測。
CAS固體平板檢測挑單菌落接種于TSB液體培養基中, 28℃、180 r/min搖床過夜培養, 吸取5 μL菌液接種于CAS固體平板上, 28℃培養箱靜置培養24h。CAS平板制備方法如下:
CAS-HDTMA溶液配制: (1)取6.0 mL十六烷基三甲基溴化銨(HDTMA)溶液(10 mmol/L)加入100 mL容量瓶中; (2) 1.5 mL FeCl3溶液(1 mmol/L, 10 mmol/L鹽酸作溶劑)和7.5 mL鉻天青溶液(2 mol/L)混勻后沿玻璃棒加入容量瓶中; (3) 4.307 g無水雙甲胺溶于30 mL ddH2O, 小心加入6.25 mL濃鹽酸, 所得溶液加入上述容量瓶中, 最后用雙蒸水定容至100 mL。
在1 L錐形瓶中依次加入NaOH 6.0 g、ddH2O 750 mL、PIPES 30.24 g、10×MM9培養基100 mL、瓊脂20 g, 調節pH=6.8, 121℃高壓滅菌20min, 待溶液冷卻至50℃左右依次加入下列無菌試劑: 10%去鐵水解酪蛋白30 mL、20%葡萄糖10 mL、1 mol/L MgCl21 mL、1 mol/L CaCl20.1 mL, 然后緩慢加入100 mL CAS-HDTMA溶液, 混勻后倒平板。
1.10 數據統計分析
實驗結果圖通過GraphPad Prism 7繪制, 采用雙尾T檢驗方法。顯著性差異定義為:*P<0.05,**P<0.01,***P<0.001。
2 結果與分析
2.1 在富鐵、低鐵條件下amoA、amoE、amoF基因轉錄水平檢測結果
嗜水氣單胞菌4LNC202分別在富鐵培養基TSB及低鐵培養基CM9中培養至對數期, 提取細菌RNA, 通過RT-PCR實驗檢測amoA、amoE、amoF基因在富鐵及低鐵條件下的轉錄水平。由圖1可知, 在低鐵條件下,amoA、amoE、amoF基因極顯著上調, 實驗結果表明amoAEF基因的表達受到鐵的調控。
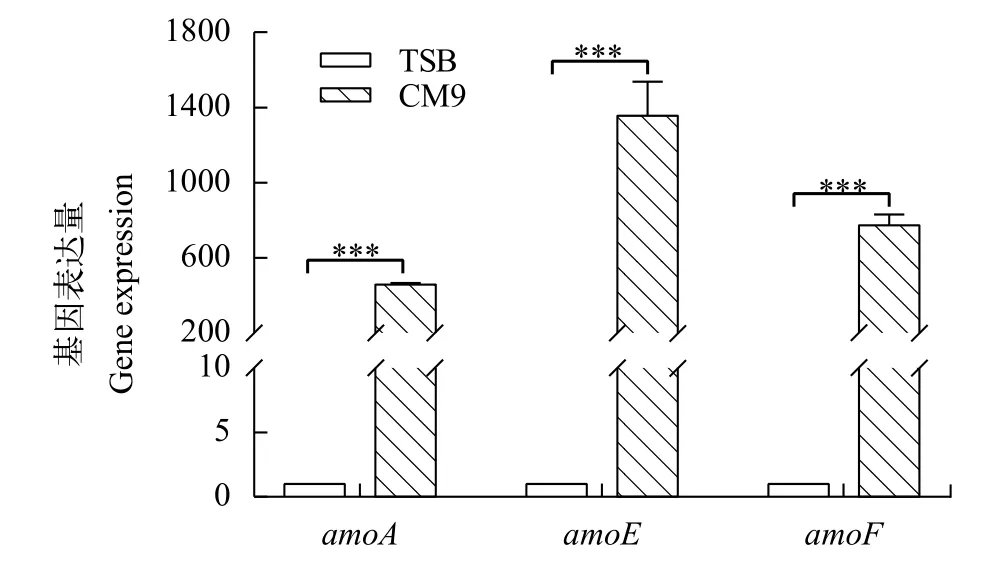
圖1 amoA、amoE、amoF表達量變化Fig. 1 amoA, amoE, amoF gene expression level
2.2 基因缺失突變株菌落PCR鑒定
重組缺失質粒送北京擎科新業生物技術有限公司測序后, 通過NCBI的BLAST功能在線比對確認序列正確無誤。
分別用目的基因上下游同源臂引物P1/P4, 對缺失株進行PCR驗證, 實際片段大小與理論大小相符合, 結果表明成功構建基因缺失株ΔamoA、ΔamoE、ΔamoF。AMA-upF-P1/AMA-doR-P4、AME-upF-P1/AME-doR-P4、AMF-upF-P1/AMF-doR-P4對野生株基因組模板進行PCR擴增產物大小分別1459、2442、3806 bp, 對各自對應基因缺失株基因組模板進行PCR擴增產物大小分別為727、837、783 bp (圖2)。
2.3 基因缺失突變株RT-PCR鑒定
進一步采用RT-PCR在RNA水平上驗證基因缺失突變株ΔamoA、ΔamoE和ΔamoF, 提取RNA、反轉錄后用引物對QAMA-F/R、QAME-F/R和QAMF-F/R分別對嗜水氣單胞菌基因缺失突變株ΔamoA、ΔamoE和ΔamoF與野生株WT進行PCR擴增。結果顯示突變株ΔamoA、ΔamoE和ΔamoF均未擴增出條帶, 而對野生菌株擴增分別得到164、141 和180 bp大小的條帶。RT-PCR結果進一步證實基因缺失突變株ΔamoA、ΔamoE和ΔamoF構建成功(圖3)。
2.4 生長能力測定
不同濃度2,2-聯吡啶作用下野生株與突變株在CM9培養基中的生長實驗嗜水氣單胞菌野生株與突變株分別在2,2-聯吡啶終濃度為50、75、100 μmol/L的CM9培養基中培養18h后, 測600 nm波長下的吸光度值(表4)。

表4 細菌在不同濃度2,2-聯吡啶下的生長實驗Tab. 4 Growth experiment of bacteria under 2,2-dipyridine at different concentrations
在不同濃度下, 野生株與突變株ΔamoA、ΔamoE和ΔamoF生長均存在極顯著性差異。當2,2-聯吡啶終濃度為50 μmol/L時, 突變株ΔamoA、ΔamoE和ΔamoF相較于嗜水氣單胞菌野生株WT生長緩慢; 當濃度大于75 μmol/L時, 突變株ΔamoA、ΔamoE和ΔamoF基本停止生長。以本實驗為依據,后續實驗中2,2-聯吡啶終濃度均為50 μmol/L。
細菌在富鐵、低鐵培養基中的生長實驗嗜水氣單胞菌野生株與突變株分別在富鐵培養基TSB和低鐵培養基CM9(2,2-聯吡啶終濃度為50 μmol/L)中培養6h, 每隔1h在波長600 nm下測吸光值。結果如下, 在TSB培養基培養時, 野生株WT與突變株ΔamoA、ΔamoE和ΔamoF生長無差異(P>0.05, 圖4A)。而用CM9(2,2-聯吡啶終濃度為50 μmol/L)培養基培養時, 野生株WT與突變株ΔamoA、ΔamoE和ΔamoF生長均存在顯著性差異(P<0.05), 突變株ΔamoA、ΔamoE和ΔamoF之間生長不存在顯著性差異(P>0.05, 圖4B)。
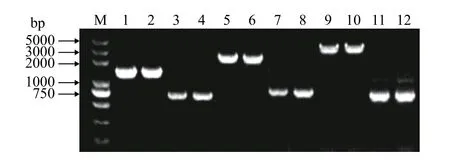
圖2 基因缺失株ΔamoA、ΔamoE、ΔamoF菌落PCR鑒定Fig. 2 PCR identification of gene deletion strains ΔamoA, ΔamoE and ΔamoF

圖3 基因缺失株ΔamoA、ΔamoE、ΔamoF RT-PCR鑒定Fig. 3 RT-PCR identification of gene deletion strains ΔamoA,ΔamoE and ΔamoF
2.5 鐵載體的檢測
Arnow實驗為了比較野生株與突變株鐵載體合成差異, 采用Arnow法進行檢測。野生株WT在波長510 nm的吸光值為0.14±0.03, 突變株ΔamoA為0.006±0.008, 突變株ΔamoE為0.277±0.013,突變株ΔamoF為0.279±0.019。野生株WT與突變株ΔamoA、ΔamoE和ΔamoF存在極顯著差異。ΔamoA相較于野生株WT極顯著降低, ΔamoE和ΔamoF相較于野生株WT極顯著升高(圖5)。
CAS固體平板實驗為了比較野生株與突變株鐵載體合成情況, 采用CAS固體平板方法進行檢測。野生株WT形成的黃色暈圈直徑為(12.907±0.381) mm, 突變株ΔamoA、ΔamoE和ΔamoF均不產生黃色的暈圈, 野生株WT與突變株ΔamoA、ΔamoE和ΔamoF存在極顯著差異(P<0.001)。實驗結果表明基因缺失突變株ΔamoA、ΔamoE和ΔamoF鐵載體合成缺陷(圖6)。

圖4 細菌在TSB (A)和CM9 (B)培養基中的生長實驗Fig. 4 Growth experiment of bacteria in TSB (A) and CM9 (B)

圖5 Arnow實驗Fig. 5 Arnow assays
3 討論
雖然嗜水氣單胞菌鐵載體的生物化學結構在30年前已經闡釋得十分清楚, 但關于其合成途徑的相關研究信息卻相對貧乏。1989年研究人員提出嗜水氣單胞菌鐵載體的合成分為兩步, 第一步合成2,3-二羥基苯甲酸(DHBA), 第二步以DHBA為前體,在此基礎上加以組裝加工[21]。Seshadri等[22]通過基因組比對后發現, 霍亂弧菌與嗜水氣單胞菌關于鐵載體的合成途徑十分相似, 推測出嗜水氣單胞菌鐵載體合成基因簇amoCEBFAGH, 這個基因簇與弧菌的vibCEBFADH基因簇相對應。vibABC主要參與DHBA的合成[23], 而vibBDEFH在組裝加工過程中發揮作用。張從剛[24]詳述了DHBA的合成過程:分支酸在異分支酸酶合成酶VibC的作用下轉變為異分支酸, 異分支酸在異分支酸酶VibB的作用下轉化為2,3-二羥-2,3二羥基苯甲酸, 接著2,3-二羥-2,3二羥基苯甲酸在2,3-二羥-2,3二羥基苯甲酸脫氫酶的作用下脫氫轉變為2,3-二羥基苯甲酸(DHBA)。本研究對野生株WT與基因缺失突變株ΔamoA、ΔamoE和ΔamoF進行Arnow實驗時, 實驗結果顯示,amoA基因缺失后所測得的A510極顯著降低, 而amoE、amoF基因缺失后使得A510值極顯著升高, 但CAS檢測實驗結果顯示基因缺失突變株ΔamoA、ΔamoE和ΔamoF都存在鐵載體合成缺陷。由此可以推斷, 基因缺失突變株ΔamoA不能合成前體2,3-二羥基苯甲酸, 而基因缺失突變株ΔamoE和ΔamoF仍然可以正常合成DHBA, 但不能對DHBA正常裝配, 導致DHBA胞外大量堆積, 從而使得A510值顯著性升高。因此,amoA基因參與鐵載體合成第一步-2,3-二羥基苯甲酸的合成,amoE、amoF參與鐵載體合成第二步, 本研究結果與張從剛結論一致。對于amoE、amoF是參與鐵載體的組裝加工具體機制還有待更進一步的研究。
在生長實驗中, 野生株WT與突變株ΔamoA、ΔamoE和ΔamoF在TSB培養基中長勢基本一致, 說明在富鐵條件下,amoA、amoE、amoF基因的缺失不影響細菌的生長; 用CM9培養基培養時, 突變株ΔamoA、ΔamoE和ΔamoF的生長能力相較于野生株WT變弱, 說明在低鐵條件下,amoA、amoE、amoF基因的缺失使得細菌的生長受到抑制。

圖6 CAS固體平板實驗Fig. 6 CAS plate assays
目前, 氣單胞菌屬不僅是魚類和蛙類的重要病原菌, 同時也被認為是引起免疫缺陷人群感染各種并發癥的致病菌[25]。鐵離子的獲取是病原菌感染過程中的重要步驟, 能夠顯著影響細菌的毒力, 而微生物從環境中攝取鐵離子的主要機制是合成分泌鐵載體。鼠疫桿菌基因突變株Δirp2鐵載體合成缺陷, 且不能在無鐵培養基中生長, 當給小鼠皮下接種時, 鼠疫桿菌野生株LD50為1.3×102, 突變株Δirp2為1.3×106, 突變株Δirp2毒力極顯著降低[26]。Heesemann等[27]、Abdelhamed等[28]、Oh等[29]、Takase等[30]研究表明鐵離子的攝取與細菌的毒性顯著相關。Zywno等[31]也曾提出鐵載體的合成與否可以作為毒性嗜水氣單胞菌的判定依據。未來可以通過進一步的攻毒實驗來比較野生株與嗜水氣單胞菌鐵載體合成缺陷株毒力的差異, 闡明鐵載體在嗜水氣單胞菌致病中的作用, 為重組弱毒疫苗的研制奠定理論基礎。此外, 對于這幾個基因在鐵載體合成中的具體分工協作機制還需進一步闡明。
本研究利用同源重組成功構建了嗜水氣單胞菌基因缺失突變株ΔamoA、ΔamoE和ΔamoF, 發現amoA、amoE和amoF基因是嗜水氣單胞菌鐵載體合成的關鍵基因, 其缺失會導致細菌在低鐵環境中的生長能力受到抑制。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
發明與創新(2016年38期)2016-08-22 03:02:52
