Lager啤酒酵母起源的歷史足跡及基因組學研究
2019-05-23 05:17:52尹花賀揚侯曉平陳璐董建軍
食品與發酵工業 2019年9期
關鍵詞:酵母菌
尹花,賀揚,侯曉平,陳璐,董建軍
(啤酒生物發酵工程國家重點實驗室(青島啤酒股份有限公司),山東 青島,266100)
1 悠久的啤酒釀造歷史
啤酒是世界上歷史最悠久、飲用范圍最廣的酒精飲料之一,是典型的農業社會發展的產物。啤酒釀造的發展歷史與人類社會和文化的發展息息相關,并且受到人類遷徙的影響。據考古學家推測,蘇美爾人和巴比倫人早在公元前6 000年開始種植大麥后不久,便開始了原始的啤酒釀造,用于祭祀女神。而公元前3 000年逐步擴展到古波斯、古巴比倫和古埃及等地。在這些社會中,啤酒既是宗教儀式中的重要元素,也是人們日常生活中的常用飲料和藥物。而后啤酒傳入歐州北部國家,并在每個國家產生了自己特色的品種[2]。斯坦福大學在2016年《美國科學院院刊》(PNAS)的考古研究報道顯示,在一個出土于西安米家崖遺址,5 000年前的陶器中發現了用于釀造啤酒的大麥、黍米等作物的殘留物[3],表明中國人釀造啤酒的歷史同樣非常悠久。原始啤酒逐步發展,后來在世界各地釀制出各種各樣的啤酒,所采用的原料有所不同,口味也各異。
現代的啤酒釀造技術源于中世紀歐洲的釀造工藝。北歐和英國的溫帶海洋性氣候為種植小麥、燕麥和大麥等谷物提供了理想的條件[4]。谷物中的淀粉在發芽和糖化的過程中能夠轉變成可發酵性糖(如麥芽糖),隨后通過野生酵母的自然發酵生成酒精。但在當時人們對于負責發酵的物質本質仍然難以理解,將其命名為“yeast”。該詞來源于德語的“Gischt”或荷蘭語的“Gist”,指的是發酵結束時容器頂部的泡沫或絮狀物質。釀酒師在每一次發酵結束后將這些“泡沫”刮下,并接種到下一批啤酒的發酵液中。這種在發酵結束后會浮于發酵罐表面的酵母被稱為“上面發酵子(top fermenters)”,這實際上就是Ale啤酒(上面發酵啤酒)的釀造[5]。Ale啤酒興起較早,也被認為和人類早期生產的啤酒比較接近。由于其發酵溫度較高,導致產品均一性不好且不易儲存,所以啤酒的穩定性和純凈度成了中世紀啤酒釀造的主要問題。
隨著啤酒釀造工業的興起,為控制啤酒的純凈度和價格頒布了一系列的法律法規。1516年在巴伐利亞頒布了《純釀法》(Reinheitsgebot),規定了啤酒只能在較為寒冷的時間進行釀造(從9月29日的圣米迦勒日到4月23日的圣喬治日之間),且釀造原料只能包括大麥、酒花、水。而法規中并未提及酵母,因為當時酵母被認為是發酵過程的一個副產物而已。而在溫度較高的季節,則將啤酒存放在塞滿了冰塊的山洞中,這一過程則被稱為“Lagering”[6],經歷了這一儲存過程的啤酒則更加穩定。漸漸的人們發現,用這種方式釀造啤酒,在發酵結束后酵母并不會浮在表面,而是沉到發酵罐的底部,因此這些酵母被稱為“下面發酵子(bottom fermenters)”。從高溫發酵轉向低溫發酵,并結合低溫儲存過程,這就是我們今天所熟知的“下面發酵”及Lager啤酒的釀造工藝。
因此,依據發酵工藝和酵母菌種的差異,現代啤酒主要分為兩大類:Ale啤酒(上面發酵啤酒)和Lager啤酒(下面發酵啤酒)。Ale啤酒發酵溫度較高,口感濃厚,香氣豐富,但發酵過程不易控制,產品均一性不好且不易儲存,因此在現代的工業啤酒生產中占比不大。與Ale啤酒相比,Lager啤酒發酵溫度低,發酵周期長,口味更加淡爽純凈,其發酵過程容易控制,產品均一性好,生產效率高,更適合標準化的工業生產。19世紀中葉,Lager啤酒逐步替代Ale啤酒成為現代工業啤酒的主流,也是全球生產和消量最大的啤酒品類。現代Lager啤酒在19世紀末引入中國,雖然初期發展緩慢,但后期迅速增長,并于2002年超越了美國,連續16年成為全球第一大啤酒生產國。根據啤酒的種類不同,發酵啤酒使用的酵母種類也不同。生產Ale啤酒的酵母即是生物學上重要模式生物——釀酒酵母(Saccharomycescerevisiae,上面發酵酵母,Ale酵母)。而Lager啤酒主要由一種雜交酵母(S.pastorianus、S.carlsbergensis、S.monacensis),統一稱為巴氏酵母(S.pastorianus,下面發酵酵母, Lager酵母)發酵而成[7]。
2 Lager酵母起源及遺傳學背景
Lager酵母是自然界雜交所產生的異源多倍體菌株。在大規模進行釀酒酵母全基因組測序之前,就已通過單基因序列和微衛星序列的對比分析證實了S.cerevisiae是Lager酵母其中的一個親本來源[8-9]。
近年來,隨著測序技術的發展和測序成本的降低,共有超過1 000株S.cerevisiae的全基因組序列陸續完成測序[10]。通過對工業釀酒酵母菌株(S.cerevisiae)基因組水平的研究,揭示了酵母菌是如何在人類的長期釀造過程中被“馴化”,以及啤酒美味背后的生物學基礎。來自比利時魯汶大學的研究團隊搜集了不同來源及工業用途多樣的157株工業釀酒酵母菌株,并完成了不同菌株的全基因組測序。通過比較工業釀酒酵母基因組及已發表的釀酒酵母基因組(包括葡萄酒酵母和面包酵母等),進一步構建了157株工業釀酒酵母的系統發生樹(圖1)[11]。顯然,S.cerevisiae可以按照其工業用途(葡萄酒、蘋果酒、面包、啤酒、清酒、蒸餾酒、臨床、實驗室)及地理來源(亞洲、北美、非洲、歐洲等)被區分開。釀造來源的S.cerevisiae形成一個獨特的群,其中大部分的啤酒酵母(85.3%)聚集在兩個子群(Beer 1和Beer 2),但這兩個子群的親緣關系較遠。Beer 1由來自不列顛群島(英國和愛爾蘭)、美國、德國和比利時的S.cerevisiae菌株組成,其中美國來源的菌株與英國菌株在系統發生樹上關系較近,表明早期是由英國定居者把釀酒酵母引入了美國。而Beer 2則與葡萄酒酵母的親緣關系更加密切[11]。
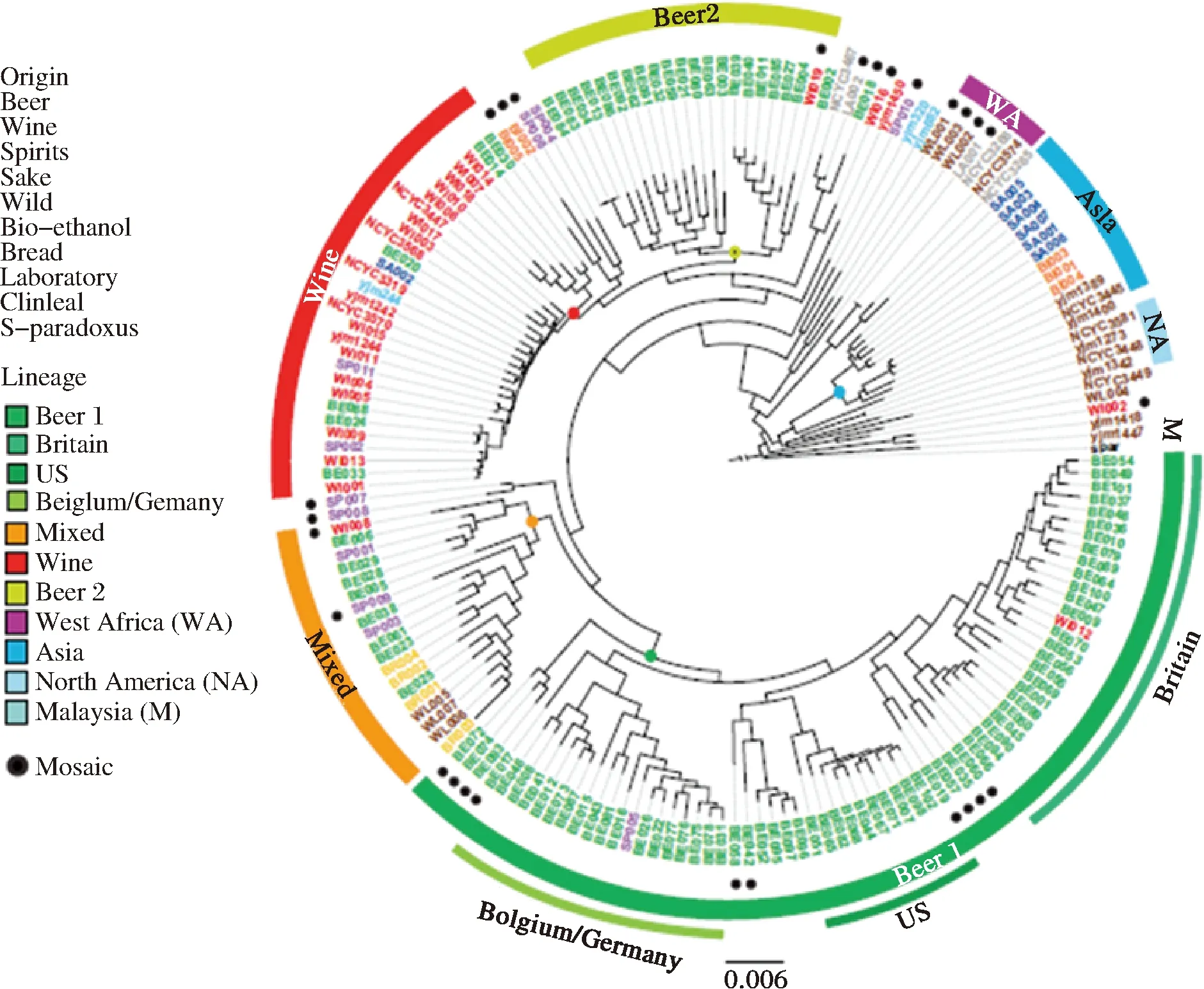
圖1 工業釀酒酵母菌株的系統發生樹Fig.1 Phylogeny structure of industrial S. cerevisiae strains
比較這兩類酵母的分支可以明顯看出,啤酒用釀酒酵母,無論是Beer 1還是Beer 2,都有較長的“樹杈”,而葡萄酒用酵母的“樹杈”則偏短[11]。這表明與葡萄酒酵母相比,啤酒酵母在工業化過程中經歷了漫長的馴化,與它們的祖先已經相去甚遠。主要表現在工業啤酒酵母的異味生成量降低(4-乙烯基-愈創木酚,4-VG)、麥芽三糖的利用率增加,且在自然條件下的存活率降低。另外,啤酒酵母在長期的工業馴化下幾乎都已失去交配能力,而葡萄酒酵母則仍保存了有性生殖的形式[11]。這與兩種酒的釀造模式是分不開的。啤酒的釀造條件受季節限制較少,一年四季都可以釀造。人們常常會用上一批啤酒使用過的酵母去釀造下一批啤酒,這無形中就完成了對啤酒酵母的人工馴化。并提出了啤酒酵母的馴化假說:啤酒酵母經歷了兩次單獨的馴化過程,第一次馴化發生在公元1573-1604年間;第二次馴化事件發生在公元1645年至1671年之間[11],這期間正是啤酒釀造從家庭手工業轉型成工業化釀造的過程。相比之下,葡萄酒則只能在特定的季節生產。在生產季節外的時間里,尚未掌握微生物學原理和操作技術的中世紀釀造人員只能將酵母留在地窖中,而它們此時則有機會和野生酵母交配,因此展示出的馴化痕跡也較少。
在另一項系統發生研究中,GON?ALVESET等將啤酒S.cerevisiae酵母分為3個進化支:小麥啤酒酵母、英國愛爾蘭啤酒酵母和德國Alt-Kolsch啤酒酵母[12]。將Lager酵母S.cerevisiae的亞基因組(Sc-亞基因組)與面包酵母、啤酒酵母、葡萄酒酵母及野外來源的90種S.cerevisiae酵母菌株進行系統發生樹分析后,Lager酵母Sc-亞基因組與啤酒S.cerevisiae的親緣關系更加密切[12]。這些結果都證實了Lager酵母中S.cerevisiae的親本來源是Ale酵母。
然而,賦予Lager酵母能夠在低溫進行發酵的非釀酒酵母來源基因序列卻一直存在爭論。嘉士伯實驗室的Morten Kielland-Brand利用單染色體轉移的技術將Lager酵母的III號染色體轉移至S.cerevisiae中,發現染色體上很大一部分無法與釀酒酵母染色體發生同源重組[13]。這暗示了Lager酵母不是一個簡單的釀酒酵母多倍體,而存在除S.cerevisiae外的其他基因來源。直至20世紀末, TAMAI等通過Southern blot雜交技術發現了S.pastorianus有兩組不同的染色體,一組跟S.cerevisiae類似,另外一組則與貝氏酵母(S.bayanus)類似[14]。因此長久以來,Lager酵母被認為是S.bayanus與S.cerevisiae雜交的產物[9,15-16]。然而日本三得利的研究學者們在2009年完成的首株Lager酵母全基因組測序結果表明,非S.cerevisiae來源的序列與S.bayanus的同源性僅為92.7%[17],說明Lager酵母可能還存在除真貝氏酵母外的其他親本來源。此外,研究發現S.bayanus是一個復雜的物種,存在純合體和雜交體兩種形式及至少兩種基因型:S.bayanusvar. bayanus和S.bayanusvar. uvarum或S.uvarum[18]。直至2011年首次于南美洲獲得了真貝氏酵母(S.eubayanus)后發現,Lager酵母中非S.cerevisiae來源的序列與S.eubayanus的序列相似性達到99.5%[19]。最終明確了工業Lager酵母的親本來源,是由S.cerevisiae與S.eubayanus通過自然雜交獲得。
真貝氏酵母(S.eubayanus)是一種耐寒的野生酵母,主要分布在南美巴塔哥尼亞地區的假山毛櫸樹和智利南洋杉、亞洲地區的櫟樹以及北美地區的雪松、紅橡樹等地,而作為Lager酵母起源地的歐洲則沒有發現野生的S.eubayanus[20-22]。S.eubayanus最早(2011年)在南美地區發現,因此提出S.eubayanus是通過歐洲殖民者在新大陸建立的貿易路線傳入歐洲的假說[23]。近年來又在中國、北美和新西蘭均分離到野生的S.eubayanus菌株[20-24],因此作為Lager酵母親本之一的S.eubayanus起源地則變得更加復雜。
2014年在中國西藏野生酵母種群及其豐富,涵蓋了發現的所有種類,多樣性遠高于美歐。中國西藏地區發現的S.eubayanus與Lager酵母中非S.cerevisiae來源基因組的序列同源性高達99.8%[23],比來自南美巴塔哥尼亞的S.eubayanus菌株相似性更高。這一結果暗示了來源于西藏的S.eubayanus菌株是Lager酵母中非Ale酵母來源的亞基因組的貢獻者。由于Lager發酵起源于1 400年的巴伐利亞,而跨大西洋貿易始于1 500年哥倫布發現新大陸后,因此S.eubayanus從南美通過貿易途徑進入歐洲的假說難以成立。而亞歐位于同一大陸,地理位置接近,商貿交往歷史悠久。中國古代與歐洲通過絲綢之路的聯系一直存在,野生酵母可以通過人員或貨物的交流而散播。因此來源于西藏的S.eubayanus被認為是Lager啤酒酵母非Ale基因組真正的野生親本。通過絲綢之路傳播到歐洲,在Lager啤酒低溫釀造時與S.cerevisiae進行雜交,賦予Lager酵母耐寒的特性。
3 Lager酵母分型
19世紀的PASTEUR和HANSEN提倡運用純種發酵來生產Lager啤酒,以避免啤酒的腐敗和異味。1909年,LINDNER首次分離了2株不同的Lager酵母菌株,分別以它們的起源地波西米亞(今捷克)Saaz跟德國Frohberg命名,稱為Group Ⅰ型(Saaz型)與Group Ⅱ型(Frohberg型)[25]。后來,兩種類型的菌株陸續傳到歐洲,并最終在世界各地進行使用[26]。Saaz型酵母現主要在捷克地區(如皮爾森啤酒廠)、丹麥的嘉士伯啤酒廠廣泛使用;而Frohberg型酵母則在德國(如Weihenstephan啤酒廠)、丹麥的其他非嘉士伯啤酒廠、荷蘭地區(如喜力啤酒廠,Oranjeboom啤酒廠)等地進行使用[8]。
這兩種菌株在發酵性能上存在顯著差異:Group Ⅰ型比Group Ⅱ型更加適合低溫發酵,Group Ⅱ型在較高的溫度下發酵更快。兩種酵母在發酵末期的存活率相當,但Group Ⅱ型菌株產生呼吸缺陷型菌株(嬌小突變)的幾率更高。芬蘭VTT技術中心通過對比在小試發酵條件下溫度等對兩種類型酵母發酵性能的影響,發現Group Ⅰ(Saaz)型酵母菌株對麥芽三糖的利用能力弱于Group Ⅱ(Frohberg)型,且Group Ⅰ型的酯類生成量較Group Ⅱ低[26]。
在2008年,DUNN和SHERLOCK等利用競爭性基因組雜交技術(CGH)分析了Lager酵母中S.cerevisiae來源、S.bayanus(現定義為S.eubayanus)來源基因的拷貝數及同源染色體的重組位點,結果也從基因組層面驗證了Lager酵母的這兩個分類(圖2)[8]。這兩種類型的Lager酵母在基因組層面上的主要區別是Group Ⅰ(Saaz型)型相對于Group Ⅱ(Frohberg型)型整體上缺失了很大一部分S.cerevisiae來源的基因組。大體上,Group Ⅰ型為三倍體,包含單倍體的Sc-亞基因組和雙倍體的S.eubayanus亞基因組(Seb-亞基因組);而Group Ⅱ型為四倍體,包含雙倍體的Sc-亞基因組和雙倍體的Seb-亞基因組[27]。這種基因組水平差異的進化原理目前尚不明確,推測其原因可能是在低溫發酵下的人為選擇。
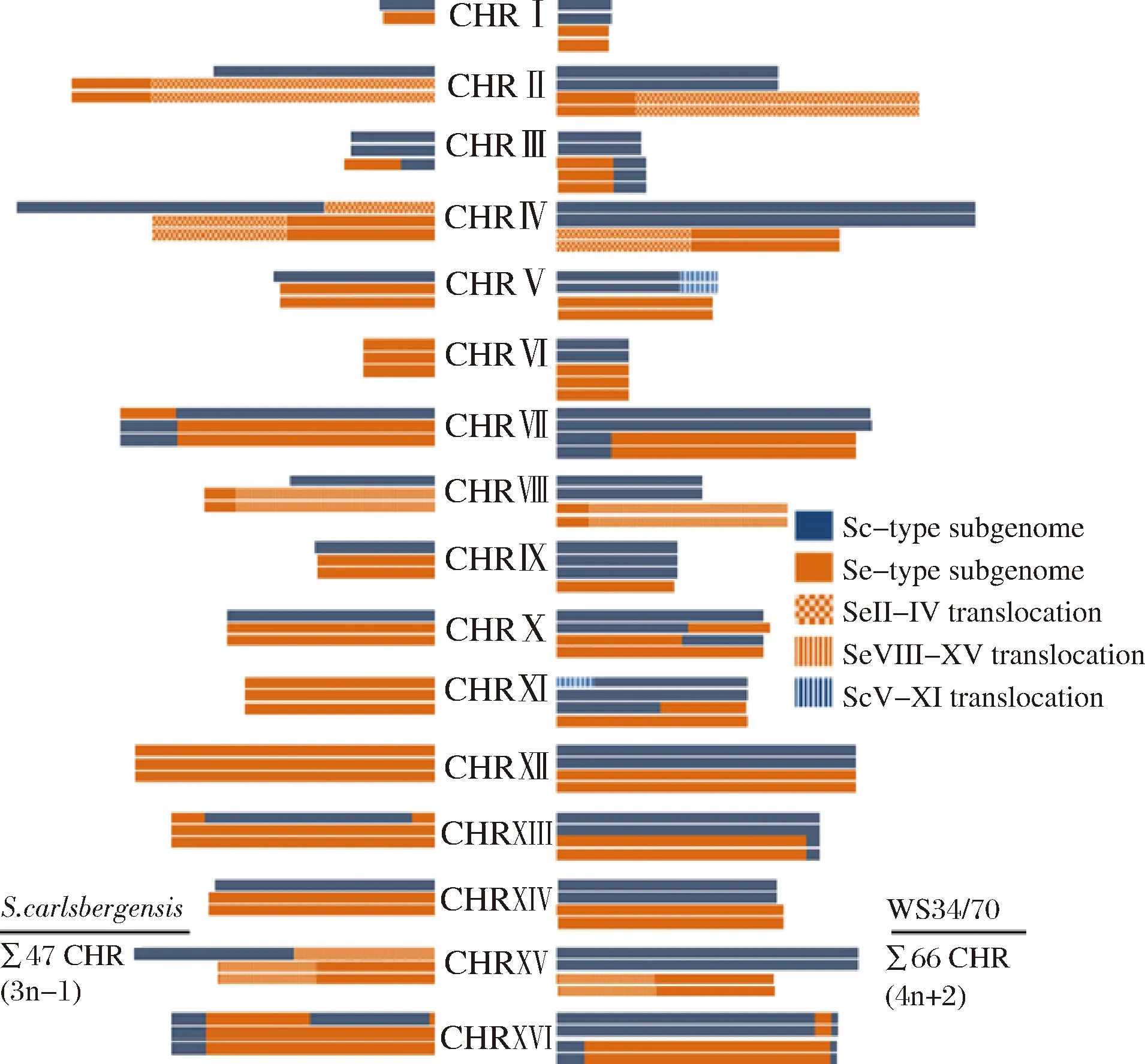
圖2 不同類型啤酒酵母(Group Ⅰ和Group Ⅱ)的染色體結構和拷貝數[27]Fig.2 Map of the chromosome structures and copy numbersof different types of Lager yeast (Group Ⅰ and Group Ⅱ)
人們對不同類型Lager酵母明確的祖先和進化過程仍然有爭議。目前主要存在3種假說,不同假說都擁有基因組學分析結果的支持[28]。第一種假說提出,Saaz型和Frohberg型的產生是起源于兩個完全獨立的雜交事件,所涉及的Ale酵母和野生S.eubayanus菌株親本基因組均存在差異(圖3-a),包括:轉座子的分布、染色體缺失模式、染色體重排等[14, 18, 29-31]。第二種假說提出,Saaz型和Frohberg型由S.cerevisiae和S.eubayanus一次雜交后分別演化而來。這種假說的依據是在這兩種不同類型的Lager酵母中發現了保守的染色體易位[32](圖3-b)。也就是說在Saaz型和Frohberg型獨自進化之前,在共同的親本染色體中發生易位,并通過雜交保留在不同類型的酵母中。在后期的進化過程中,Frohberg型酵母中Sc-亞基因組中經由雜合性丟失而降低了同源染色體之間的差異,而Saaz型則丟失了一條S.cerevisiae來源的染色體。第三種假說是結合以上兩種假說所提出,認為Saaz型和Frohberg型起源于單倍體S.cerevisiae和二倍體S.eubayanus之間至少一個共有的雜交事件。首先單倍體S.cerevisiae和二倍體S.eubayanus之間的單一雜交事件產生三倍體,進一步分化為Saaz型;而Frohberg型是由三倍體進一步與單倍體S.cerevisiae發生二次雜交所產生的[33](圖3-c)。
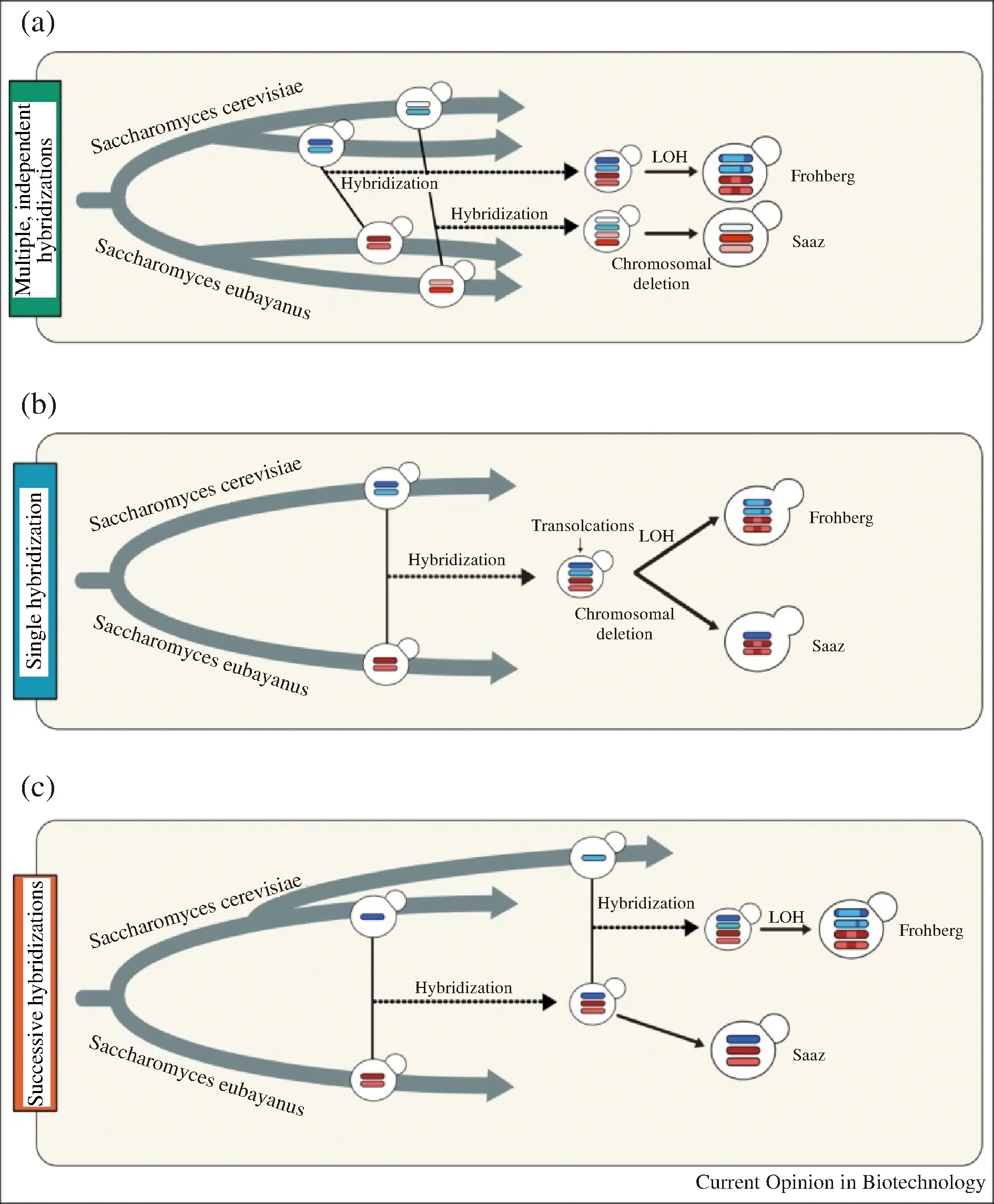
圖3 Saaz型和Frohberg型起源的現有模型[28]Fig.3 Current models for the origin of Frohberg andSaaz lineages
4 Lager酵母的基因組結構
第一個完成全基因組測序的Lager酵母Weihenstephan 34/70 (WS 34/70),是Lager啤酒釀造中被廣泛使用的Frohberg型菌株,由日本三得利在2009年完成[17]。其多倍體基因組大小為25 Mb,包含36條染色體,分別由釀酒酵母(S.cerevisiae)和貝氏酵母(S.bayanus)兩個亞基因組構成,而后來貝氏酵母的亞基因組被確認是來自真貝氏酵母(S.eubayanus)[19]。從DNA含量的預測結果來看,該Lager酵母基因組為四倍體,但實際的染色體數量卻較低,這也意味著在其染色體上存在基因的多拷貝現象[17]。其中,Sc-亞基因組與S.cerevisiae模式菌株S-288C的序列同源性高達99.2%,而S.bayanus來源的基因序列與當時唯一完成測序的S.bayanus菌株CBS 7001的序列相似性僅有92.7%。進一步通過分析基因組序列證實了Lager酵母中包含3種類型的染色體:類S.cerevisiae染色體、類S.bayanus染色體和雜合型染色體(來自于前兩種的同源重組)[17]。同時還發現WS 34/70的線粒體基因組與S.bayanus菌株CBS 7001同源性很高,表明Lager酵母的線粒體DNA(mt DNA)應該是來源于非S.cerevisiae[17]。
在獲得了WS 34/70的基因組序列后,近年來又有多株Lager酵母菌株陸續完成了全基因組測序,重點在于對不同菌株進行比較基因組學分析,并為進一步探索不同類型Lager酵母起源提供了基因組學基礎[31-37]。Lager酵母基因組學的研究能夠從生物進化的角度來闡明雜交菌株是如何通過基因的融合來實現對環境的適應。通過比較3株Lager酵母的染色體斷點位置及序列發現,不同親本來源的序列大多是在基因編碼區發生染色體重組,而不是在基因組的重復元件(Repetitive elements)上。重組所產生的大量嵌合基因會減弱系統發生的信號,這才導致了在Lager酵母起源的研究上存在大量爭議[35]。通過對Lager酵母染色體拷貝數的變化研究表明,Group Ⅰ型菌株的染色體組型較為一致,而Group Ⅱ型菌株之間則表現出較大的差異。此外,基因組序列基本一致的Lager酵母菌株在染色體拷貝數上也存在明顯的變化[36]。
BAKER等指出,不同類型Lager酵母中Sc-亞基因組在與Seb-亞基因組發生雜交之前就已產生差異,在兩個親本的亞基因組中中性差異位點的數量存在差異。因此支持在Lager酵母進化過程中發生兩次獨立的雜交事件的假說[29]。與2009年對WS 34/70線粒體基因組分析結果不同的是,OKUNO等通過對Lager酵母mt DNA的系統發生學分析,揭示了Lager酵母的線粒體基因組可以從兩個親本的任意一方獲得。此外還發現,Group Ⅰ型菌株和Group Ⅱ型菌株的mt DNA具有相同的來源(西藏)[37]。此外,研究發現在兩種類型的Lager酵母基因組上至少存在3處相同的染色體間易位,認為Group Ⅰ型和Group Ⅱ型是由相同的祖先進化而來;但通過SNVs的分析卻證明了不同類型Lager酵母基因組中含有不同的Sc-亞基因組,說明Group I型和Group Ⅱ型在其進化史上存在獨立的進化起源[37]。因此認為兩種類型Lager酵母至少經歷了一次共同的雜交事件,進一步證明了第三種進化假說的可信性。
在Lager酵母基因組中,大多數的基因存在Sc-和Seb-兩個同源拷貝。但研究發現,在某些基因中,一個來源的基因拷貝由于發生了移碼突變或提前終止密碼子會導致功能缺失[38-39]。由于有些基因是酵母生存的必需基因,因此推測這種突變是在兩個親本雜交之后產生的[17]。對雜交菌株來說,它平等地繼承了親本雙方的基因及其特性,但隨時間的推移及雜交菌株所受到的進化壓力,一個親本的基因組則會表現出其主導地位,從而改變菌株的性狀[40-44]。這種性狀的改變是由雜交菌株的基因組不穩定性所引起。許多研究也正是利用了Lager酵母的這種特性,通過人為引導進化(適應性進化)獲得了性能優良的酵母菌株[45-50],包括高濃釀造Lager酵母、抗脅迫Lager酵母等。
5 結束語
在漫長的啤酒釀造歷史中,人們無意識地對酵母進行了人工馴化及篩選,并釀造出美味的啤酒。與其他發酵產業不同,大規模啤酒釀造中所使用的Lager酵母菌株仍存在局限性。在生命科學領域早已步入基因組時代的今天,如何利用全新的基因組學知識指導優良酵母菌株的篩選也是當今的研究重點。Lager酵母菌株的單倍體分離及雜交技術現已十分成熟,實現Lager酵母菌株的定制化也不再是夢想。目前,國際許多研究機構都在進行大規模菌株雜交的方式篩選新的酵母菌株,并用來釀造啤酒。利用現代生物學知識和高通量實驗技術,通過人工選擇獲得的酵母菌株與它們的祖先相比是否能夠生產出口味更佳、更具競爭力的啤酒呢?還是讓我們拭目以待!
猜你喜歡
發明與創新·初中生(2024年6期)2024-06-16 00:00:00
昆明醫科大學學報(2022年1期)2022-02-28 07:43:38
中國民間療法(2021年8期)2021-07-22 05:53:28
軍事文摘·科學少年(2021年1期)2021-02-04 08:03:45
當代水產(2019年3期)2019-05-14 05:43:24
天然產物研究與開發(2018年9期)2018-10-08 03:25:32
煙草科技(2015年8期)2015-12-20 08:27:04
藥學與臨床研究(2015年4期)2015-06-05 11:35:51
閱讀與作文(小學低年級版)(2015年8期)2015-05-30 10:48:04
食品工業科技(2014年9期)2014-03-11 18:15:31
