外生菌根真菌對干旱條件下油松幼苗生長的影響
2019-05-21 08:26:56王秀麗柳昱旻高潤梅
中國水土保持科學 2019年2期
關鍵詞:質量
王秀麗, 柳昱旻, 高潤梅
(山西農業大學林學院,030801,山西晉中)
干旱脅迫是限制干旱與半干旱地區植物生長最重要的因子,森林作為中國北方自然生態系統的主體與生態屏障,尤其夏季干旱環境下,多數樹種難以實現更新[1],森林生態系統生產力下降,水土流失嚴重,生態恢復困難。我國植樹造林過程中存在苗木成活率低、適應性差和不能保持水土流失等問題。研究發現外生菌根真菌(ectomycorrhizal fungi)能增強樹木對干旱、病蟲害、鹽堿等傷害的適應性與抵抗力[2-3]。樹木在遭受干旱脅迫時,外生菌根真菌通過根外菌絲運送土壤中的礦質元素與水分到根內供其吸收[4],同時獲得其生長繁殖所需的碳水化合物,與宿主形成一種互惠共生的關系[5],利于提高樹木抗性,促進其生長發育[3]。外生菌根可有效改善樹木的水分生理狀態,使其在持續干旱下也可成活,或復水后很快恢復生長[6]。
油松(Pinustabuliformis)屬松科(Pinaceae)松屬(Pinus),為陽性樹種[3],對干冷氣候的適應能力強;其根系發達、生長迅速、耐干旱、耐貧瘠,抗風沙能力強[7],防治水土流失能力強,可促進土壤肥力的恢復與土壤結構的改善。作為華北地區低山的主要造林樹種,油松可保持水土、防風固沙,有效改善生態環境,對修復退化的生態系統有重要作用[8]。
雖然油松菌根資源豐富,但大部分仍未應用于生產實際,具優良促生效果的菌劑較少[3]。有研究表明,木霉(Trichodermaspp.)對油松幼苗具一定促生效果[9]。筆者通過室內盆栽試驗,研究木霉對油松幼苗抗旱能力的影響,以期為華北地區干旱半干旱低山地區的油松造林提供技術支撐,促進樹種更新,進而防治水土流失。
1 材料與方法
1.1 幼苗培養
2016年 10月,在山西省關帝山采取油松種子,室溫下自然晾干,冷藏在0~4 ℃冰箱中。2017年12月,挑選大小均勻顆粒飽滿的種子,于山西農業大學林學院樹木學實驗室室內育苗。試驗前,溫水浸種2 d,0.1%的KMnO4溶液振蕩消毒30 min后播種。培養基質由進口草炭與蛭石(體積比2∶1)組成,經高溫高壓蒸汽滅菌(121 ℃,0.1 MPa,2 h)后備用,基質裝入滅菌的黑色營養缽(長寬高為10 cm×10 cm×9 cm),每缽10粒,覆1 cm厚的基質,澆透水后,室溫培養,待種子萌發后,每缽定苗至4株,供試材料共64缽256株幼苗。
1.2 試驗設計
采用雙因素裂區試驗設計,重復8次,主處理T為菌根真菌,設接種木霉(T+)和不接種(T-)2個處理。木霉由山西農業大學樹木病理實驗室提供,PDA培養基上活化培養15 d后使用。播種同時進行接種處理。32缽每缽注射10 mL菌絲懸浮液;另32缽每缽注射10 mL滅活后冷卻的菌絲懸浮液(140 kPa高溫滅菌30 min)。副區處理D為干旱脅迫處理,設4個水分梯度:正常供水(control,CK),澆水間隔期3 d,土壤相對含水量(relative soil water content,RSWC)70%~80%;輕度脅迫(light drought,LD),澆水間隔期5 d,RSWC為50%~60%;中度脅迫(moderate drought,MD),澆水間隔期10 d,RSWC為35%~50%;重度脅迫(severe drought,SD),澆水間隔期15 d,RSWC為20%~35%[2]。以15 d為處理周期,連續進行3個周期水分處理,45 d后結束試驗,進行指標測定。
1.3 幼苗指標測定
1.3.1 生長指標 每個處理挑選長勢基本一致的10株幼苗,測量每株幼苗的株高、針葉長度(選最長針葉測定),精確到0.01 cm;生物量測定:每處理取30株幼苗,洗凈擦干,分地上與地下部分,105 ℃下,殺青10 min[10],80 ℃下烘干至恒質量,分別稱量地上和地下部分的干質量[11];根冠比:根冠比(R/T)=地下部分干質量/地上部分干質量。
菌根真菌依賴性(Mycorrhizal fungi dependence,MFD)反映植物對菌根真菌依賴性大小,數值越大,依賴性越強,接種菌根后對植物的促生效果越好(MFD≥300%,屬強依賴性;MFD≥200%,屬中等依賴性;MFD<200%,屬弱依賴性)[12]。MFD計算方法為:MFD=(接種株干質量/對照株干質量)×100%
1.3.2 生理指標 剪取幼苗針葉,進行生理指標測定:可溶性蛋白質含量用考馬斯亮藍法測定;蒽酮比色法測定可溶性糖含量;蒽酮硫酸法測定淀粉含量。具體操作參照文獻[13]。
1.4 數據處理
數據統計分析采用SPSS 21.0,以多因素方差分析(Univariate)和最小顯著差數法(least significant difference,LSD)比較油松幼苗不接種木霉各個水分處理組、接種木霉各個水分處理組、相同水分處理組接種與不接種木霉油松幼苗各指標的差異顯著性,差異水平為0.05。
2 結果與分析
2.1 接種木霉對干旱脅迫下油松幼苗形態的影響
正常水分下,接種木霉顯著促進油松幼苗的地上生物量、地下生物量、根冠比(P<0.05,表1);菌根苗的地上生物量、地下生物量、根冠比分別是非菌根苗的2.05、2.38、1.16倍;對木霉的菌根依賴性屬中等依賴性。

表1 油松幼苗對木霉的菌根依賴性Tab.1 Effect on the growth ofPinus tabulaeformisseedlings inoculatedTrichodermaspp. under drought stress
注: *表示同列的差異顯著(P<0.05); T+、T-分別表示接種木霉和不接種木霉。下同。* indicates significant difference between the same column (P<0.05); T+, T-respectively indicate inoculationTrichodermaspp. or not. The same as below.
研究發現接種木霉與干旱脅迫對油松幼苗的株高和針葉長度均具極顯著影響(P<0.01),但交互作用影響不顯著(P>0.05)。不接種時,油松幼苗在CK、LD、MD、SD下的株高分別為3.88、3.51、3.25和2.96 cm,即隨脅迫加劇,幼苗株高遞減,與CK相比,LD、MD、SD分別減少9.47%、16.24%、23.65%;接種木霉后,隨干旱脅迫加劇,株高仍呈下降趨勢,LD、MD、SD分別下降7.70%、10.15%、17.11%(圖1(A))。表明隨著干旱脅迫加劇,接種木霉的油松幼苗株高降低程度減緩。
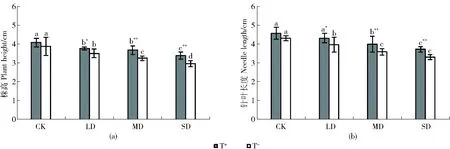
CK、LD、MD、SD分別表示對照、輕度脅迫、中度脅迫與重度脅迫;柱形圖上方不同的小寫字母表示不同水分處理間差異顯著(P<0.05);**表示接種木霉與不接種在相同水分處理下差異極顯著(P<0.01),*表示接種木霉與不接種在相同水分處理下差異顯著(P<0.05)。下同。CK, LD, MD, and SD respectively refers to control, light drought, moderate,sever drought; different lowercase letters above the histogram showed significant differences between different water treatments (P<0.05). ** indicated that the difference betweenPinus tabulaeformisseedlings inoculatedTrichodermaspp. or not at the same water treatment was very significant(P<0.01). * indicated that the difference betweenPinus tabulaeformisseedlings inoculatedTrichodermaspp. or not at the same water treatment was significant(P<0.05). The same as below.圖1 接種木霉對干旱脅迫下油松幼苗的株高與針葉長度的影響Fig.1 Effect on plant height and needle length ofPinus tabulaeformisseedlings inoculatedTrichodermaspp. under drought stress
不接種時,CK、LD、MD、SD組油松幼苗的針葉長度分別為4.30、3.95、3.58和3.30 cm,整體呈下降趨勢,LD、MD、SD分別下降8.14%、16.86%、23.26%;接種木霉后,油松幼苗針葉長度均有所增長,但隨著干旱脅迫的加劇,整體仍呈遞減趨勢,LD、MD、SD分別減少5.43%、12.55%、18.64%(圖1(b))。與不接種相比,隨著干旱脅迫的加劇,接種木霉的油松幼苗針葉長度降低程度減緩,具有明顯的增長作用。
2.2 接種木霉對干旱脅迫下油松幼苗生理指標的影響
滲透調節是植物在干旱環境下的重要生理機制[2],參與滲透調節的溶質稱為滲透調節物質,可溶性蛋白質、可溶性糖與淀粉作為主要的滲透調節物質,干旱脅迫下,其含量變化可以反映出植物對逆境的抗性[14]。接種木霉與干旱脅迫對油松幼苗針葉的可溶性蛋白質、可溶性糖與淀粉含量影響極顯著(P<0.01),但交互作用對針葉的可溶性蛋白與淀粉含量影響不顯著(P>0.05), 對可溶性糖含量影響極顯著(P<0.01)。
2.2.1 可溶性蛋白質質量分數 油松幼苗針葉CK組的可溶性蛋白質質量分數最高,隨著干旱脅迫的加劇,質量分數逐漸降低。不接種時,LD、MD與SD分別比CK降低66.73%、77.91%、84.81%;相同水分梯度下,接種木霉后,油松幼苗針葉的可溶性蛋白質質量分數顯著高于不接種木霉的幼苗,且與CK相比,LD、MD與SD分別減少59.79%、66.92%與75.64%(圖2),即接種木霉的油松幼苗,其針葉的可溶性蛋白質質量分數降低趨勢減緩。

圖2 接種木霉對干旱脅迫下油松幼苗可溶性蛋白質質量分數的影響Fig.2 Effect on content of soluble protein inPinus tabulaeformisseedlings inoculatedTrichodermaspp. under drought stress
2.2.2 可溶性糖質量分數 干旱脅迫下,油松幼苗針葉的可溶性糖質量分數均高于CK組,隨著干旱脅迫的加劇,整體呈“先升后降”的趨勢;未接種和接種木霉的幼苗可溶性糖質量分數分別在LD和MD達到最高值,MD和SD幼苗針葉的可溶性糖質量分數開始下降,與不接種相比,接種木霉后,油松幼苗耐旱閾值增大,抗性更強(圖3)。
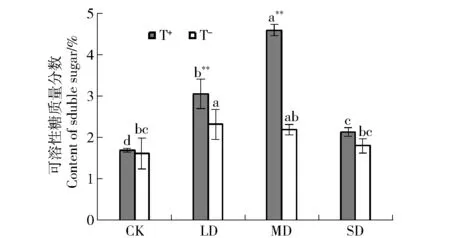
圖3 接種木霉對干旱脅迫下油松幼苗可溶性糖質量分數的影響Fig.3 Effect on content of soluble sugar inPinus tabulaeformisseedlings inoculatedTrichodermaspp. under drought stress
2.2.3 淀粉質量分數 與干旱脅迫下的幼苗相比,CK組油松幼苗針葉淀粉質量分數最高,淀粉質量分數隨著干旱脅迫的加劇,整體呈下降趨勢;不接種木霉時,LD、MD與SD分別降低15.56%、26.88%、57.79%;接種木霉后,LD、MD與SD分別減少8.68%、17.35%、38.08%(圖4);與不接種相比,隨著干旱脅迫的加劇,接種木霉的油松幼苗針葉淀粉質量分數降低程度減緩,表明接種木霉對于油松幼苗抗旱能力的提高具有明顯的促進作用。
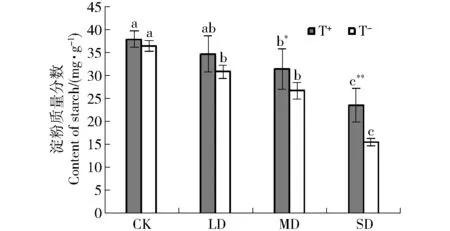
圖4 接種木霉對干旱脅迫下油松幼苗淀粉質量分數的影響Fig.4 Effect on content of starch inPinus tabulaeformisseedlings inoculatedTrichodermaspp. under drought stress
3 討論
植物苗期較易受逆境影響[15],筆者發現干旱脅迫下,接種木霉可促進油松幼苗生長,這與王藝等[4]對馬尾松的研究結果一致。接種木霉后,顯著促進油松幼苗的株高、針葉長度與生物量等形態指標,主要原因在于菌根菌可擴大幼苗根系的吸收面積和范圍,增加植株對礦質元素的吸收,改善植株礦質營養,減少水分散失,使植株更好地利用和保持土壤水分[16]。其次,真菌通過自身或利用周圍微生物及植物等分泌多種酶類和次生代謝產物來分解有機質,活化土壤中的營養物質[17],改善土壤結構與環境等。
可溶性蛋白質是針葉中氮的重要存在形式,其質量分數下降是針葉衰老的主要標志[18]。接種木霉后,油松幼苗針葉的可溶性蛋白質質量分數均顯著高于對照;但隨著脅迫加劇,可溶性蛋白質質量分數呈遞減趨勢;可能在于干旱脅迫抑制蛋白質合成并誘導其降解,引起蛋白質質量分數下降[19]。可溶性糖是植物體內碳水化合物代謝和暫時貯藏的主要形式[20],其質量分數變化可反映植株碳水化合物的合成與運輸情況[18]。接種木霉后,油松幼苗針葉可溶性糖質量分數增加,利于植株保持較高的滲透壓,以抵御干旱造成的不利影響[19]。其中:MD條件下可溶性糖質量分數達到最高值,此時油松幼苗抗性較強;SD條件下可溶性糖質量分數下降,可能由于重度脅迫下氣孔關閉,導致碳攝取受阻,如果脅迫持續可能影響幼苗存活[21]。淀粉和可溶性糖作為主要的非結構性碳水化合物,其質量分數變化可表征植物對干旱脅迫的緩沖能力。接種木霉后,油松幼苗淀粉質量分數升高;隨著干旱脅迫的加劇,其質量分數呈遞減趨勢;當植物遭受干旱脅迫時,土壤含水量減少,油松幼苗針葉積累的淀粉轉化為可溶性糖,使油松在低水勢下保持較高膨壓,降低干旱對植物組織的傷害[22]。總之,干旱脅迫顯著抑制油松幼苗的生長,但接種木霉后脅迫作用顯著減緩,在形態與生理層面均有表征。
4 結論
1)外生菌根真菌促進了干旱脅迫下油松幼苗的吸收功能,增加生物量的積累,進而促進了植株的生長。
2)中度干旱脅迫下,接木霉對油松幼苗生長的促進作用效果最顯著,推廣使用過程中,應根據實際情況把握好干旱脅迫的閾值,在閾值內進行復水處理。
3)相同水分處理下,菌根苗比非菌根苗的抗旱能力增強,菌根化苗木復水后也能更快恢復生長。
猜你喜歡
中學生數理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(2018年6期)2018-04-22 03:16:54
產品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
