香芹酚對耐甲氧西林金黃色葡萄球菌(USA300)抑菌機制的研究
2019-05-15 07:35:14甘盈盈袁中偉張天翼谷可欣申瀚君周能華尹立子
四川農業大學學報 2019年2期
關鍵詞:影響
甘盈盈,袁中偉,張天翼,谷可欣,李 超,申瀚君,周能華,尹立子
(四川農業大學動物醫學院,成都 611130)
金黃色葡萄球菌(Staphylococcus aureus)是一種極其常見的致病菌,數量大,分布廣,水、空氣、塵土中都可找到它的蹤影。金黃色葡萄球菌可以引起局部化膿性感染、全身炎癥反應綜合征和敗血癥等多種疾病,臨床體征較為嚴重[1]。耐甲氧西林金黃色葡萄球菌(Methicillin-resistant Staphylococcus aureus,MRSA)由金黃色葡萄球菌演變而來,毒力和致病力顯著增強,并且具有多重耐藥的特點,臨床治療的難度更大[2]。USA300 是一種具有高度致病性和全球流行性的社區獲得性耐甲氧西林金黃色葡萄球菌,它對紅霉素、左氧氟沙星、莫匹羅星和四環素等抗菌藥物的耐藥性增強。它主要引起皮膚和軟組織感染,還會造成嚴重的危及生命的疾病,如壞死性肺炎、骨髓炎、和膿毒性關節炎[3-4]。長期以來,萬古霉素是抗MRSA 最有效的藥物,由于萬古霉素的使用量漸增,導致MRSA 對其抵御能力增強,陸續出現多種耐藥菌株[5]。因此,為避免臨床上抗MRSA 陷入一種無藥可施的境地,開發新的抗MRSA 藥物迫在眉睫。
香芹酚,又稱香荊芥酚,是一種單萜酚類化合物。香芹酚廣泛分布于芳香植物中,是麝香草、牛至草、亞加菊等芳香植物揮發油的主要化學成分[6-9]。香芹酚既具有分子小、極性低和毒性低等特點[10],又具有抗菌、抗氧化和抗癌等生物活性[11-12],因而具有很大的開發價值。香芹酚的抗菌活性較好,對金黃色葡萄球菌、綠膿桿菌等均有較好的抑制效果[11],然而目前關于香芹酚對金黃色葡萄球菌的抑菌機理尚不完全明確。因此本文以MRSA 的標準菌株USA300 為試驗菌株,考察香芹酚對MRSA 的抑菌機制,為香芹酚開發為新型天然抑菌藥提供理論依據。
1 材料和方法
1.1 試驗材料
MRSA 標準菌株:USA300(ATCC BAA-1717),購自美國標準菌種庫;香芹酚(純度>98%,上海麥克林生物科技有限公司)用DMSO(分析純,索萊寶生物科技有限公司)配制成母液備用;pH 7.0 的磷酸緩沖鹽粉劑,購自索萊寶生物科技有限公司,按使用說明配制成2 L 的PBS 磷酸緩沖溶液備用;乳酸脫氫酶(lactate dehydrogenase,LDH)試劑盒,購自南京建成生物工程研究所;BCA 蛋白濃度測定試劑盒,購自上海碧云天生物技術有限公司;LB 肉湯和LB 營養瓊脂,購自北京奧博星生物技術有限責任公司;TSB 液體培養基和BHI 液體培養基,購自青島海博生物科技有限公司。
1.2 香芹酚對USA300 抑菌活性和生長曲線的測定
通過肉湯稀釋法和菌落計數法測定香芹酚在37 ℃下作用于USA300 24 h 的最低抑菌濃度(minimum inhibitory concentration,MIC)和最低殺菌濃度(minimum bactericidal concentration,MBC)[13-14]。試驗設立負對照組。試驗重復3 次,當試驗結果完全一致或者相差一個質量/體積濃度梯度時才被接受[15]。
將USA300 菌懸液置于37 ℃,150 r/min 的搖床中培養至OD600nm=0.3,混勻,均分于6 個50 mL的錐形瓶中,分別加入香芹酚,使其香芹酚的質量/體積濃度依次為0、32、64、128、256 和512 μg/mL;將其置于37 ℃、150 r/min 的搖床中培養,分別于0、1、2、4、6、8、16 和24 h 測定肉湯的OD600nm,采用Prism 7.0 繪制生長曲線。
1.3 香芹酚對USA300 細胞膜的影響
參考F.Bendali、張冠楠[16-17]等的試驗方法,將生長繁殖至對數期(OD600nm=1.8)的USA300 菌懸液4 500 r/min、4 ℃離心10 min,棄去上清液,收集下層的細菌沉淀,用5%葡萄糖溶液懸浮分散成107cfu/mL的菌懸液,混勻,均分于2 個250 mL 的錐形瓶中;加入香芹酚,使其質量/體積濃度分別為0 和512 μg/mL,混勻,置于4 ℃恒溫培養箱中靜置培養。于0、1、2、4、6 和8 h 取出菌懸液,4 500 r/min、4 ℃離心10 min,測定上清液的電導率。試驗設立負對照組。試驗重復3 次,計算平均值。
1.4 香芹酚對USA300 DNA 外滲量的影響
取培養至對數生長期(OD600nm=1.8)的USA300菌懸液,4 500 r/min 離心10 min,棄去上清液,收集下層的細菌沉淀,用PBS 磷酸緩沖溶液重新完全分散成107cfu/mL 的菌懸液,混勻,均分于2 個250 mL的錐形瓶中;加入香芹酚,使其質量/體積濃度分別為0 和512 μg/mL,混勻,置于37 ℃的恒溫培養箱中靜置培養。于0、1、2、4、6 和8 h 取出菌懸液,4 500 r/min 離心10 min,收集上清液,用微量分光光度計NanoDrop One(Thermo,America)測定上清液中DNA 含量[18]。試驗設立負對照組。試驗重復3次,計算平均值。
1.5 香芹酚對USA300 可溶性蛋白質含量的影響
制備生長繁殖至對數期(OD600nm=1.8)的USA 300 菌懸液,混勻,均分于2 個200 mL 的錐形瓶中,加入香芹酚,使其香芹酚的質量/體積濃度分別為0 和512 μg/mL,混勻,置于37 ℃,150 r/min 的搖床中培養。于2、6、10 和16 h 取出菌懸液,用新鮮的LB 肉湯稀釋至OD600nm=0.6,并定容至50 mL,4 500 r/min 離心10 min,棄去上清液,收集下層的細菌沉淀。用1 mL 的PBS 磷酸緩沖液將細菌沉淀重新完全分散,進行超聲破碎,混勻,得到超聲破碎液;向超聲破碎液中加入5×上樣緩沖溶液(超聲破碎液體積:5×上樣緩沖溶液體積=4∶1),混勻,在沸水浴中煮沸5 min,4 500 r/min 離心10 min,取上清液進行SDS-PAGE 試驗[19];按照BCA 蛋白濃度測定試劑盒的試驗方法檢測超聲破碎液中可溶性蛋白質的含量。試驗設立負對照組。試驗重復3 次,計算平均值。
1.6 香芹酚對USA300 乳酸脫氫酶含量的影響
制備生長繁殖至對數期(OD600nm=1.8)的USA300菌懸液,混勻,均分于2 個200 mL 的錐形瓶中,加入香芹酚,使其香芹酚的質量/體積濃度分別為0和512 μg/mL,置于37 ℃,150 r/min 的搖床中培養。于0、2、4、6 和8 h 取出菌懸液,用新鮮的LB肉湯稀釋至OD600nm=0.6,并定容至50 mL,4 500 r/min離心10 min,棄去上清液,收集下層的細菌沉淀,用1 mL 的PBS 磷酸緩沖液重新完全分散,超聲破碎后4 500 r/min 離心10 min,用LDH 試劑盒測定上清液中LDH 的含量[20]。試驗設立負對照組。試驗重復3 次,計算平均值。
1.7 電鏡觀察香芹酚對USA300 形態結構的影響
制備生長繁殖至對數期(OD600nm=1.8)的USA300菌懸液,混勻,均分至2 個100 mL 的錐形瓶中,加入香芹酚,使其香芹酚質量/體積濃度為0 和512 μg/mL,置于37 ℃,150 r/min 的搖床中培養。于1、10 和16 h 時取出菌懸液,4 500 r/min 離心10 min,收集下層細菌沉淀,用PBS 磷酸緩沖液清洗3 次,棄去上清液,得到菌體沉淀[21],并制成掃描電鏡、透射電鏡樣品[22-24],于S-3400N 掃描電子顯微鏡(日立,日本)和H-600IV 型透射電鏡(日立,日本)下觀察細菌表觀形態與結構的變化。試驗設立負對照組。試驗重復3 次。
1.8 亞抑菌濃度下的香芹酚對USA300 生物被膜形成能力的影響
制備香芹酚質量/體積濃度依次為0、16、32、64和128 μg/mL 的BHI 液體培養基(各50 mL),各接種0.5 mL USA300 菌懸液,置于37 ℃,150 r/min 的搖床中培養至OD600nm=0.6。向96 孔板中加入上述菌懸液10 μL,再對應加入含相同質量/體積濃度香芹酚的3%蔗糖BHI 液體培養基290 μL,混勻(混合液中香芹酚保持最初的質量/體積濃度),置于厭氧條件、37 ℃的培養箱中靜置培養18 h。清除96 孔板中所有液體后,每孔加入100 μL10%的甲醛溶液,常溫靜置培養18 h;移除所有液體,加入100 μL 質量濃度為1 mg/g 的結晶紫溶液,常溫靜置30 min;清除所有液體,用雙蒸水沖洗,晾干,加入200 μL 33%的乙酸溶液,混勻,用酶標儀(Thermo,America)測量OD490nm,以此反映USA300 菌株生物被膜的形成能力[25]。試驗設立負對照組。試驗重復3 次,計算平均值。
1.9 統計學分析
采用Prism 7.0 對試驗數據進行差異顯著性分析(t 檢驗),結果用“平均值±標準誤”表示,P<0.05為差異顯著,P<0.01 為差異極顯著。
2 結果與分析
2.1 抑菌活性及生長曲線的測定
香芹酚具有良好抑菌活性,MIC 是128~256 μg/mL,MBC 是512 μg/mL。香芹酚對USA300 生長曲線的影響結果如圖1所示,香芹酚濃度為128、256 和512 μg/mL 時,對USA300 的生長有著明顯抑制作用;香芹酚濃度為32 和64 μg/mL 時,對USA300 的生長沒有顯著的影響。
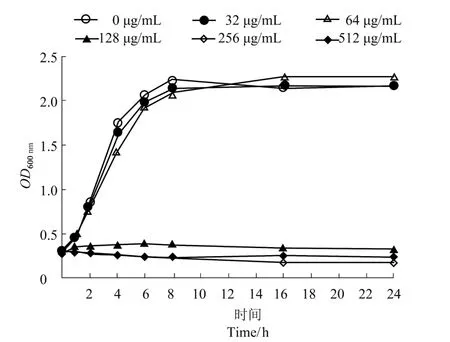
圖1 香芹酚對USA300 生長的影響Figure 1 The effect of carvacrol on the growth of USA300
2.2 香芹酚對USA300 細胞膜通透性的影響。
當細胞膜被藥物破壞時,其滲透性增加,無機鹽外漏,導致菌懸液的電導率增加[26],因此本試驗通過考察香芹酚作用于USA300 菌懸液后電導率的變化,研究香芹酚對細胞膜通透性的影響。由圖2可知,香芹酚作用顯著、迅速;與0 h 相比,香芹酚作用1 h 后,菌懸液電導率增長5.57%±0.40%(P<0.01);香芹酚作用8 h 后,菌懸液電導率增長4.22%±0.71%(P<0.01)。對照組的電導率在0~8 h 里無顯著變化。相同培養時間下,給藥組與對照組組間差異極顯著。
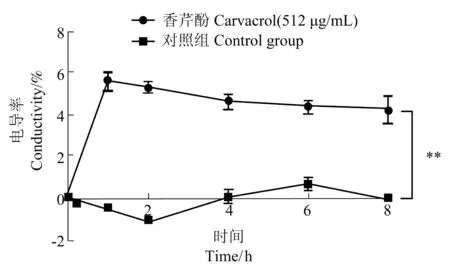
圖2 512 μg/mL 的香芹酚對USA300 菌懸液電導率的影響Figure 2 The effect of carvacrol at 512 μg/mL on the conductivity of USA300 bacterial suspension
2.3 香芹酚對USA300 DNA 外滲量的影響
本試驗測定了香芹酚作用于USA300 后DNA的外滲量,以此驗證香芹酚對USA300 細胞膜滲透性的影響。如圖3,與0 h 相比,加入香芹酚1 h 后,USA300 的DNA 外滲量增加(16.27±1.86)μg/mL(P<0.01);加入香芹酚8 h 后,USA300 DNA 外滲量增加(21.80±2.00)μg/mL(P<0.01)。相同培養時間下,給藥組與對照組組間差異極顯著。該試驗結果表明香芹酚可以顯著影響USA300 的DNA 外滲量。
2.4 香芹酚對USA300 可溶性蛋白含量的影響
BCA 蛋白濃度測定試劑盒測定結果顯示,與相同培養時間的對照組相比,香芹酚作用于USA300菌體10 h 和16 h 后,可溶性蛋白含量分別增加61.13%±7.63%(P<0.01)和85.76%±1.84%(P<0.01)。SDS-PAGE 結果和BCA 蛋白濃度測定試劑盒測定結果共同表明了香芹酚可以刺激USA300 可溶性蛋白含量增加。如圖4,SDS-PAGE 蛋白譜帶結果顯示,香芹酚可以影響USA300 可溶性蛋白含量。
2.5 香芹酚對USA300 乳酸脫氫酶的影響
LDH 乳酸脫氫酶試劑盒測定結果如圖5所示,香芹酚作用于USA300 1、2、4、6 和8 h 后,與相同培養時間的對照組相比,乳酸脫氫酶的含量分別增加了40.49%±2.77%(P<0.01)、20.13%±2.28%(P<0.01)、19.10%±2.82%(P<0.01)、13.59%±1.35%(P<0.01)和11.65%±2.85%(P<0.05)。此試驗結果表明香芹酚可以刺激USA300 的LDH 含量增加。
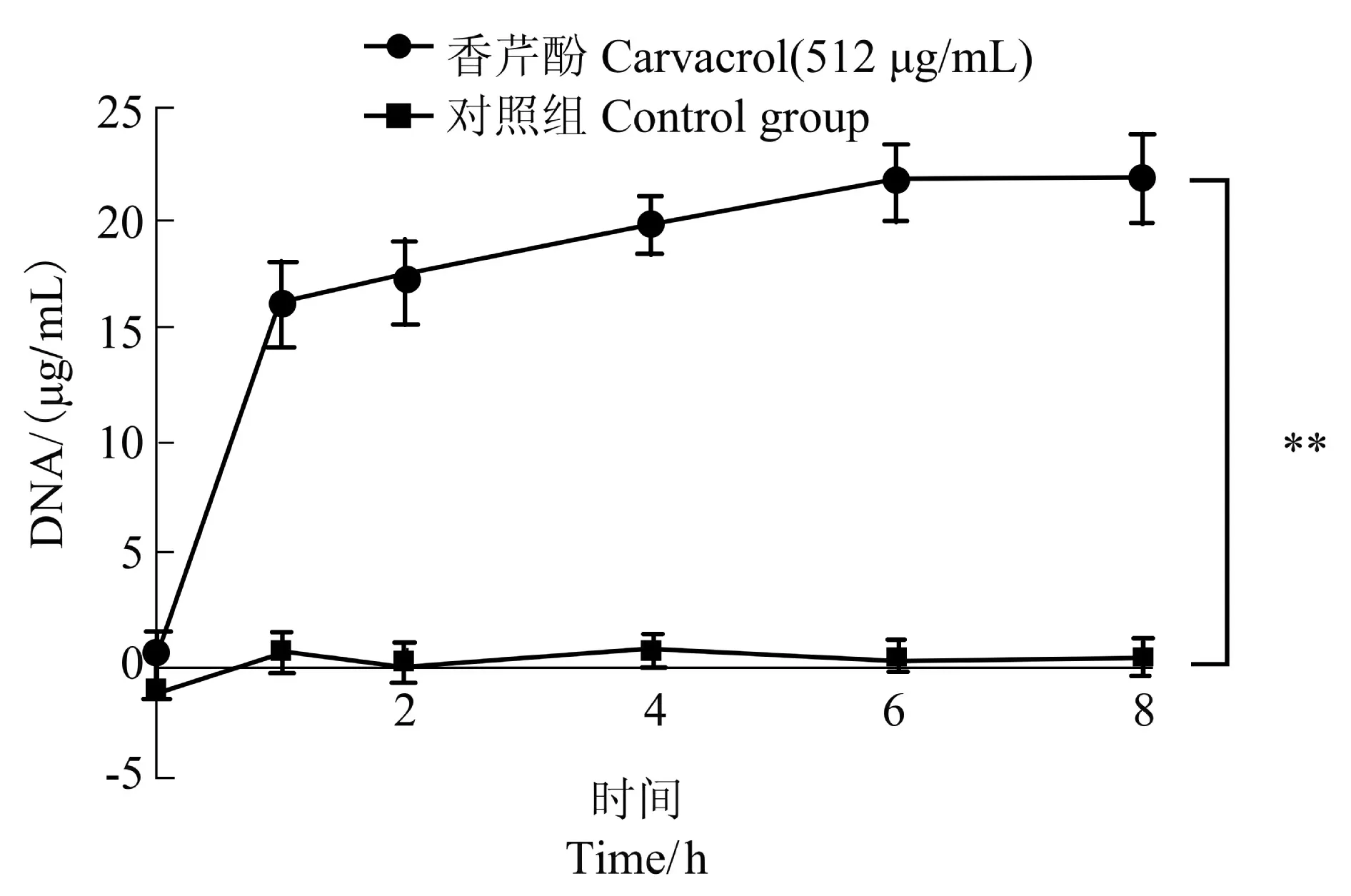
圖3 512 μg/mL 香芹酚對USA300DNA 外滲量的影響Figure 3 The effect of carvacrol at 512 μg/mL on DNA exosmosis amount of USA300
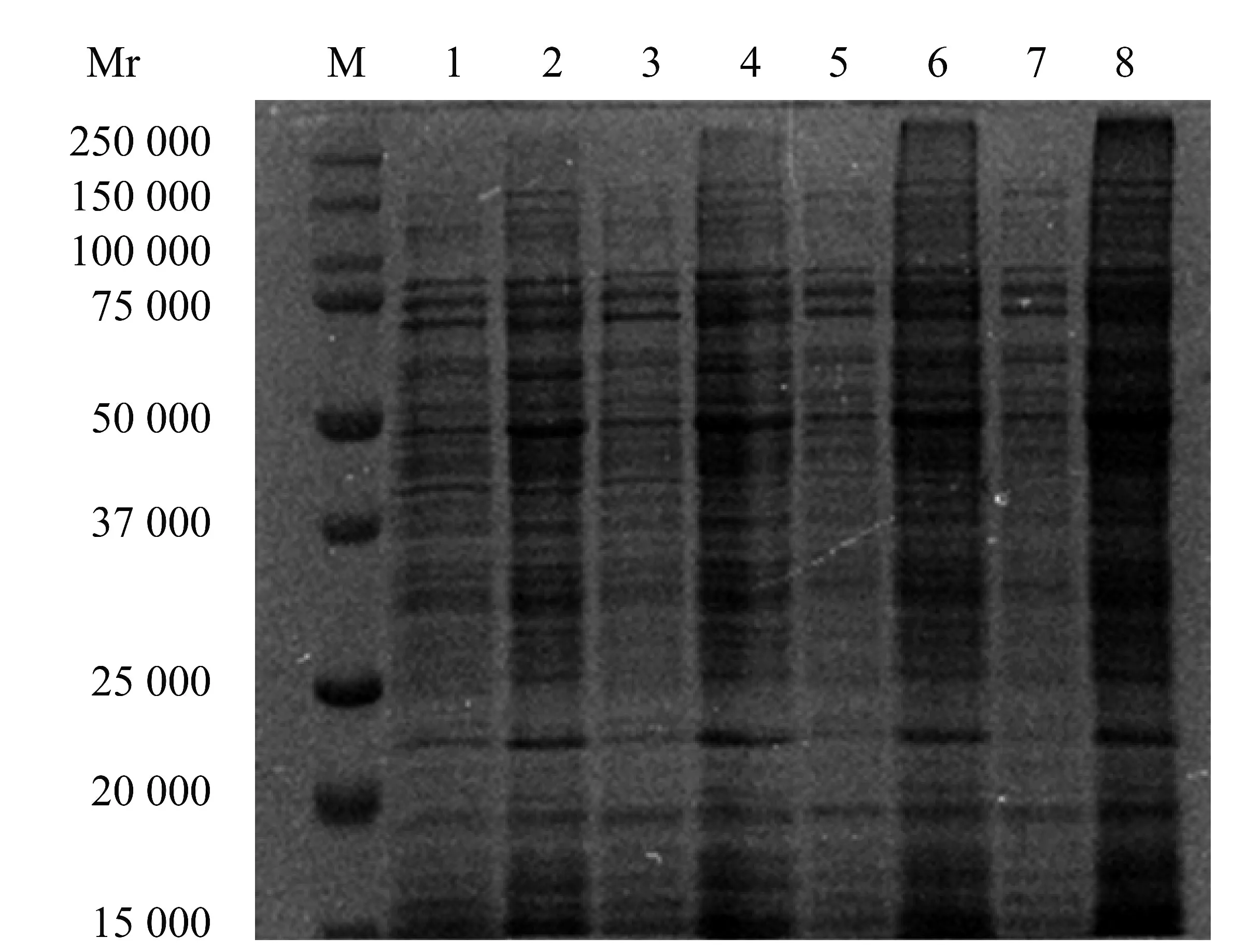
圖4 512 μg/mL 香芹酚對USA300 可溶性蛋白質含量的影響Figure 4 The effect of 512 μg/mL carvacrol on the soluble protein content of USA300
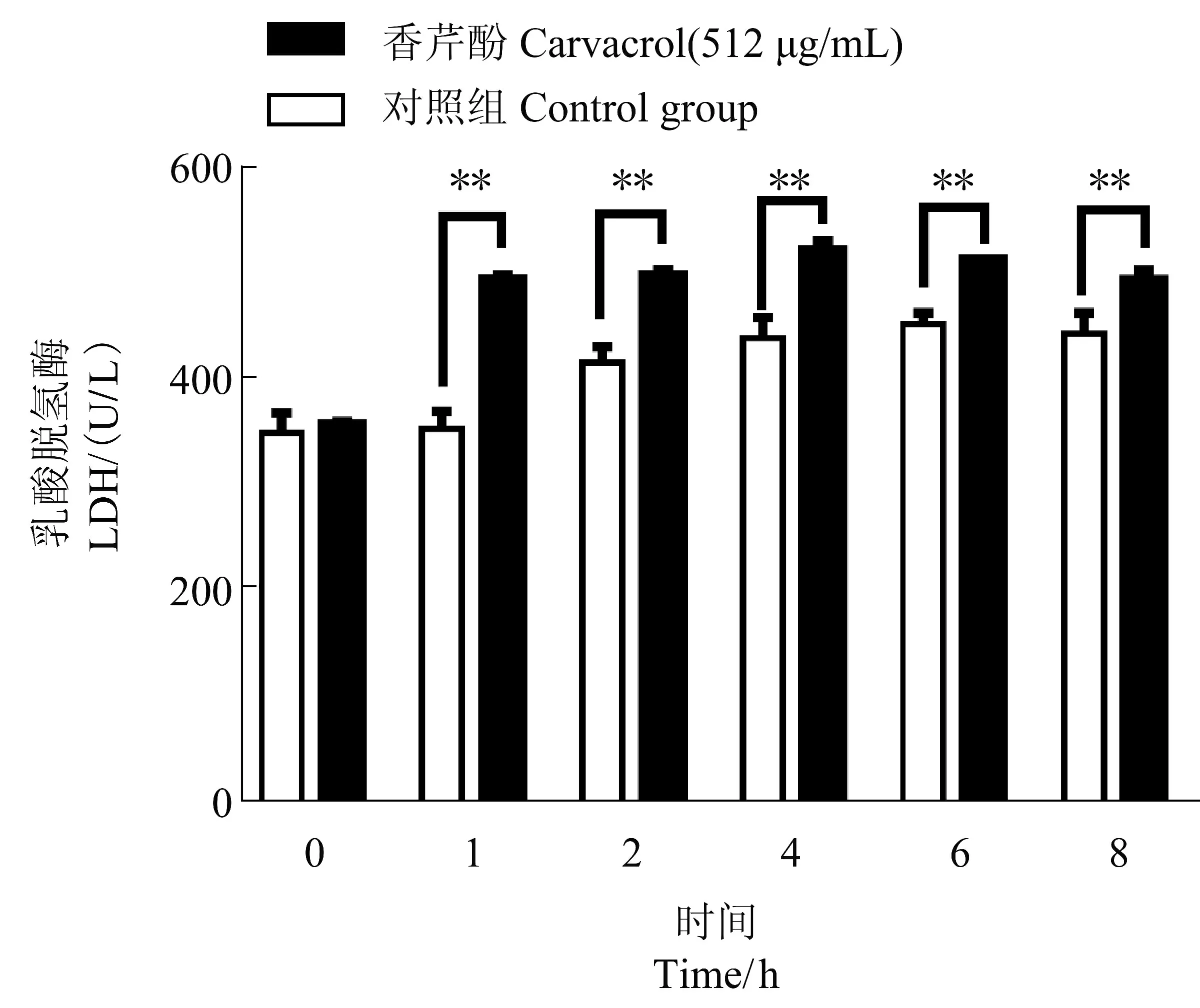
圖5 512 μg/mL 香芹酚對USA300 的LDH 含量的影響Figure 5 The effect of carvacrol at 512 μg/mL on LDH content of USA300
2.6 香芹酚對USA300 形態結構的影響
掃描電鏡圖像顯示,對照組(圖6A1)的USA300表面光滑,具有正常的表觀形態;512 μg/mL 香芹酚作用于USA300 1 h 后,菌體表面褶皺,菌體萎縮變形(圖6A2)。透射電鏡圖像顯示,對照組10 h(圖6B1)和對照組16 h(圖6C1)菌體表觀正常、結構清晰并具有完整的細胞壁;512 μg/mL 香芹酚作用于USA300 10 h(圖6B2)后,蛋白質異常沉淀,內容物固縮,出現空泡化的現象;作用16 h 后(圖6C2),菌體細胞壁結構被破壞,質壁分離,出現細胞壁溶解、脫落的現象。
2.7 香芹酚對USA300 生物被膜形成能力的影響
低于MIC 的藥物濃度稱為亞抑菌濃度。由圖7可知,加入亞抑菌濃度的香芹酚后,USA300 生物膜的形成比對照組顯著減少。香芹酚質量/體積濃度為16、32、64 和128 μg/mL 時,形成的生物被膜比對照組分別減少27.55%±2.33%(P <0.01)、35.28%±3.67%(P<0.01)、68.86%±0.53%(P<0.01)和77.47%±1.74%(P<0.01)。該試驗結果表明,香芹酚能抑制USA300 生物被膜的形成。
3 討論與結論
隨著抗菌藥物的大量使用,細菌耐藥性呈上升趨勢,MRSA 現已成為臨床上十分常見的致病菌[27-28]。MIC、MBC 和生長曲線試驗結果顯示香芹酚對USA300 具有較好的抑菌作用,表明香芹酚具有開發為抗MRSA 的天然抑菌藥的潛在價值。

圖6 512 μg/mL 香芹酚不同作用時間對USA300 菌體細胞形態結構的影響Figure 6 Effect of different treating time of carvacrol at 512 μg/mL on the morphology and structure of USA300 cells
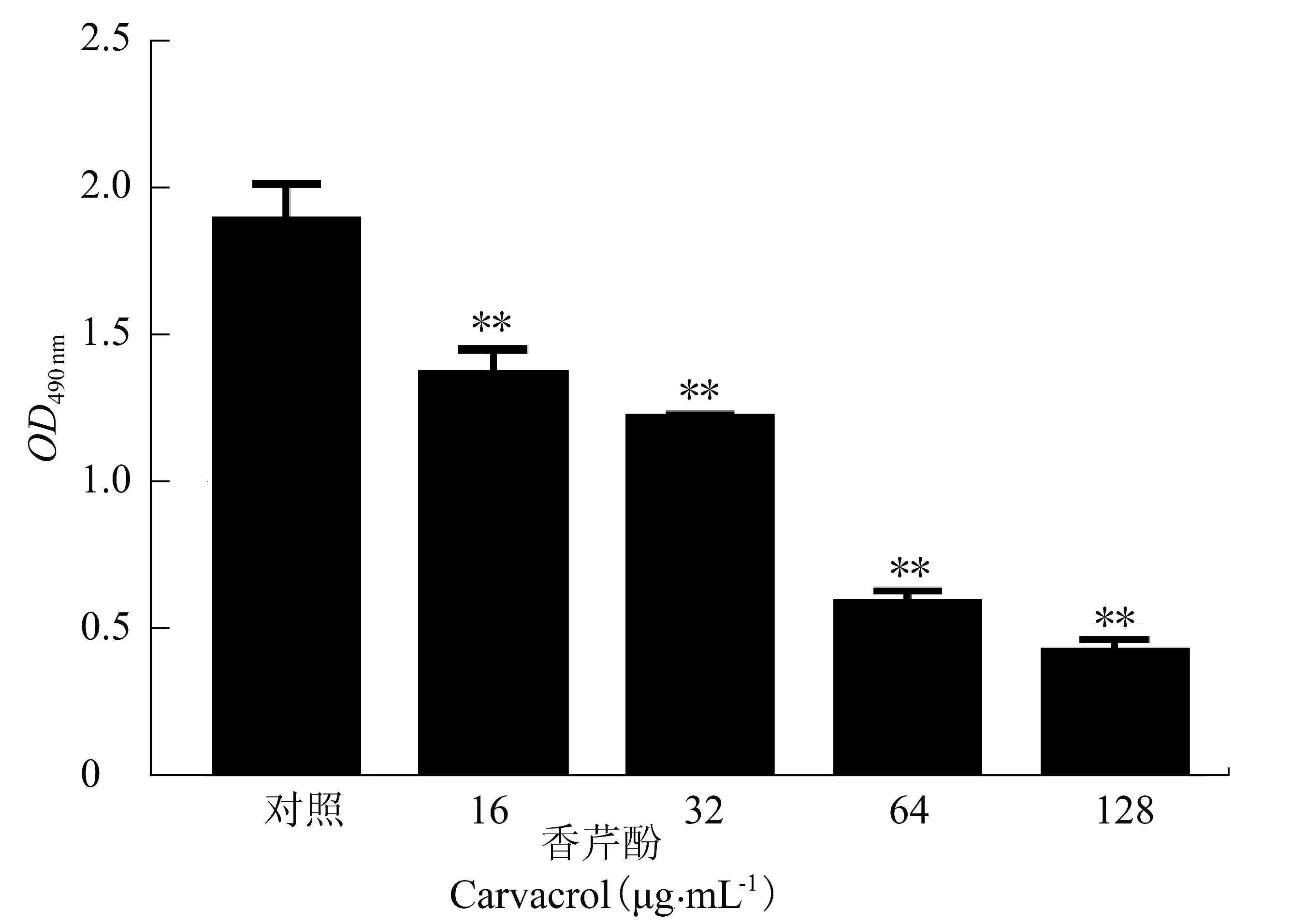
圖7 不同質量/體積濃度的香芹酚對USA300 的生物被膜形成能力的的影響Figure 7 The effect of carvacrol with different mass concentrations on biofilm formation ability of USA300
細胞膜是保護細菌的天然屏障。許多抗菌藥物可以通過改變或損傷細胞膜的結構使細菌生長受到抑制乃至死亡,從而達到抑菌、殺菌的目的[29-30]。本研究中電導率試驗表明,香芹酚作用迅速、明顯,與對照組相比,加入香芹酚1 h 后,菌懸液電導率顯著上升,說明香芹酚可以迅速影響USA300 菌株的細胞膜通透性。DNA 是一種高分子聚合物,細菌正常生長情況下,DNA 幾乎不會通過細胞膜外滲而出。DNA 外滲量測定結果表明,香芹酚作用于USA300 菌株1 h 內,DNA 外滲量顯著增加,說明香芹酚可以在短時間內破壞細胞膜的結構,再次驗證電導率的試驗結果。
蛋白質是生命中至關重要的物質基礎,可分為可溶性蛋白和非可溶性蛋白兩大類。可溶性蛋白是細菌重要的滲透壓調節物質,它可以提高細菌的保水能力,保證細菌的生命活動正常進行[31]。SDSPAGE 試驗結果和BCA 蛋白濃度測定結果顯示,香芹酚作用于USA300 菌體后,菌體的可溶性蛋白含量顯著增加,分析原因可能是香芹酚誘導了USA300 抗性蛋白及其相關蛋白的增加,菌體通過此生命活動來抵抗香芹酚的傷害,暗示了干擾蛋白質代謝可能不是香芹酚的抑菌機制。LDH 是一種重要的可溶性蛋白,它對于金黃色葡萄球菌保持致病力、逃避宿主的先天性免疫是極其重要的[32]。LDH含量測定試驗結果表明,香芹酚作用于USA300 后,菌體內LDH 含量明顯增多,其原因可能是香芹酚誘導了USA300 生物酶的表達,菌體以此來抵抗香芹酚的損傷和維持自身的生長代謝,暗示影響酶的活性可能不是香芹酚發揮抑菌作用主要途徑,再次驗證了SDS-PAGE 試驗的試驗結果。
掃描電鏡和透射電鏡在抑菌機制的研究中被廣泛應用。本試驗的圖像表明,香芹酚可以直接損傷菌體的細胞壁,影響其結構的完整性。細胞壁可以維持細菌正常的形態結構,當香芹酚損傷細胞壁后,菌體形態發生改變,出現形變萎縮的現象。
細菌生物被膜既具有天然屏障的作用[33],又可以改變被膜內微環境[34],因此極大地增加了細菌對抗生素的耐藥性。生物被膜試驗結果顯示,亞抑菌濃度下,香芹酚抑制生物被膜形成的能力與香芹酚濃度呈正相關,且都不影響細菌的正常生長。此試驗結果表明,亞抑菌濃度下,香芹酚可以有效抑制MRSA 生物被膜的形成,降低耐藥性的產生,進一步說明香芹酚具有開發為抗MRSA 感染藥物的潛力。
綜上所述,香芹酚具有良好的抗MRSA 活性,其抑菌機制主要是通過破壞USA300 的細胞壁、改變細胞膜的通透性來抑制USA300 的生長繁殖,并且其在亞抑菌濃度下能有效抑制生物被膜的形成。其他抑菌機制還需要進一步研究。
猜你喜歡
英語文摘(2022年4期)2022-06-05 07:45:18
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
知識經濟·中國直銷(2016年3期)2016-02-27 16:15:49
現代檢驗醫學雜志(2014年6期)2014-02-02 03:02:04
