埃博拉病毒分子生物學研究進展
2019-05-15 01:10:46孫英堯
中國獸藥雜志 2019年4期
關鍵詞:功能
孫英堯,董 浩
(吉林農業大學生命科學學院,長春 130118)
埃博拉病毒(Ebola virus, EBOV)是一種十分罕見的病毒,它是在1976年同時暴發的兩起疫情中首次出現的,一起在現在的南蘇丹恩扎拉,另一起在剛果民主共和國揚布庫。后者發生在位于埃博拉河附近的一處村莊,該病由此得名。感染埃博拉出血熱(Ebola virus disease, EVD)通常潛伏期為4~10 d,初期癥狀與流感類似,表現為嘔吐與腹瀉,隨后身體內外出血,伴隨有突發休克,最后是多器官衰竭。病人一般在出現癥狀后的6~16 d死亡。盡管目前已研發了一些疫苗可對埃博拉病毒進行防護,但我們對埃博拉病毒的了解還不夠。本文就埃博拉病毒的分子生物學研究進展作一綜述,以期為更深入的研究提供參考。
1 埃博拉病毒的分類學地位
埃博拉病毒屬于單股反意病毒目(Mononegavirales)絲狀病毒科(Filoviridae)埃博拉病毒屬(Ebolavirus)。該屬又包含5個種:本迪布焦埃博拉病毒(BundibugyoEbolavirus,BEBOV)、雷斯頓埃博拉病毒(RestonEbolavirus,REBOV)、蘇丹埃博拉病毒(SudanEbolavirus,SEBOV)、大森林型埃博拉病毒(TaiForestEbolavirus;IvoryCoastEbolavirus,ICEBOV)、扎伊爾型埃博拉病毒(ZaireEbolavirus,ZEBOV)。絲狀病毒科下還有兩個屬,為馬爾堡病毒(Marburgvirus,MAEV)與奎瓦病毒(Cuevavirus)[1]。
2 埃博拉病毒分子生物學研究進展
埃博拉病毒(圖1)直徑80 nm,長度為970~1200 nm,進入細胞后病毒的長度可達14000 nm[2]。埃博拉病毒的反意基因長達19 kb,基因組呈線狀,無分段,包含有7個基因與兩個調節區段。兩個調節區段位于3’頭端與5’尾端。3’頭端有雙向復制的啟動子,也是第一個基因的轉錄起始位點。5’尾端編碼復制啟動子的互補序列[3]。7個基因分別對應編碼埃博拉病毒的7種病毒蛋白,分別為:核蛋白(nucleoprotein,NP)、糖蛋白(glycoprotein,GP)、L聚合酶蛋白(L-polymerase protein,L)、病毒蛋白24(viral protein24,VP24)、病毒蛋白30(viral protein30,VP30)、病毒蛋白35(viral protein35,VP35)與病毒蛋白40(viral protein40,VP40)[4]。7種蛋白的分子生物學特征綜述如下:
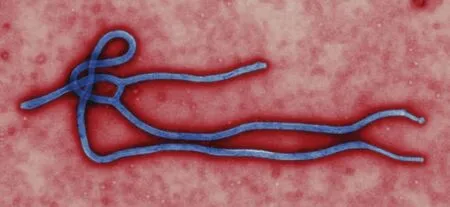
圖1 埃博拉病毒電鏡照片[5]Fig 1 Electron microscope photography of Ebola virus[5]
2.1 病毒蛋白24(VP24) VP24是絲狀病毒的常見蛋白。以往的研究表明,該蛋白的功能是抑制宿主機體產生α、β與γ干擾素,阻礙干擾素基因的翻譯過程。VP24能夠通過結合真核細胞核孔的核轉運蛋白以及抑制KappaB(NF-KB)通路來阻止編碼干擾素基因的轉錄、信號轉導與磷酸化過程[6]。
VP24也是病毒組裝衣殼的重要蛋白,它增強了不同結構蛋白之間的協同催化作用。通過觀察C、N末端缺失的VP24,發現N末端編碼的VP24具有調控衣殼蛋白形成的作用[7],說明VP24在病毒衣殼合成過程中起的作用應該為催化作用或者為調控VP35與NP。另有研究表明,VP24與VP40的共同表達能夠形成大量的類病毒粒子(VLPs),遠比VP40單獨表達時多得多,推測可能是VP24的小干擾RNA(siRNA)降低了病毒的釋放量,使大量的病毒蛋白保留在被感染的細胞內。此外還有研究表明,VP24結合在VP35的病毒衣殼外表面,起到了調節NP層、增強衣殼機械強度穩定性的作用,增強了NP、VP35、VP24的協同關系[8]。
2.2 病毒蛋白35(VP35) VP35的主要作用是影響宿主的免疫系統,使病毒免于被宿主的免疫機制清除。VP35是1型干擾素的拮抗物,通過與雙鏈RNA結合抑制干擾素調節因子3,同時抑制1型干擾素RIG-1的信號傳導從而降低α型與β型干擾素的生成[9]。VP35還能通過多種途徑影響宿主整體的免疫系統。VP35通過增加活躍的STAT1(轉錄活化蛋白1)調節干擾素調節因子7的活性,抑制干擾素生成toll樣受體與RIG-1的活性。已有證據表明,VP35還可以解除RNA沉默,可以使先天免疫信號傳導與宿主的抗病毒反應失效[10]。
此外,VP35也有協助病毒復制的功能。VP35的C末端可以結合雙鏈RNA,而N末端可以結合NP。當VP35的N末端與NP解離時,NP會活化,隨后與病毒RNA結合,高度激活基因組的轉錄活性,隨后的NP解離,并出現寡聚現象,形成寡聚NP。但NP單體并沒有寡聚的傾向,僅在上述的生化反應中才會出現寡聚。此外,其他病毒中VP35與NP的作用反而會降低NP的寡聚化。對這一現象的研究可能是研發治療埃博拉出血熱的突破口。
2.3 病毒蛋白30(VP30) VP30是埃博拉病毒的一種次要核蛋白質,主要作用是起始病毒基因組的轉錄,并調節細胞的動態磷酸化過程[11]。最新的報道發現,VP30還具有解除RNA干擾的功能,但是具體機制尚不明確。當存在siRNA時,VP30會影響主要參與RNA干擾的蛋白的帽子結構,這時即使是VP35的N末端RNA結合域已經結合,但VP30的干涉仍會啟動。這表明VP30蛋白的RNA結合功能非常廣泛,而且不止限定于宿主或者病毒自身的RNA,似乎存在一定的堿基結構選擇性,但是這一功能是否為病毒RNA轉錄所必須的,目前尚不可知。
2.4 病毒蛋白40(VP40) VP40是病毒組裝的基石[12],其主要作用是形成六聚物——這一結構被認為是埃博拉病毒衣殼的基礎結構,功能為埃博拉基因組復制與RNA的結合[13]。
以往的研究多針對于該蛋白作用調節生成VLPs,而最近的研究表明,VP40還可以產生外泌體結構,殺死免疫細胞。外泌體的出現一般是在病毒感染細胞之后, VP40導致的外泌體接觸到未成熟的T細胞和單核白血球細胞后,會引起這兩者的凋亡及活性下降。關于VP40在致病機制中的作用,有可能是開發新治療手段的切入點。已有研究用酪氨酸磷酸激酶對其進行酪氨酸殘基加聚磷酸基團,降低VP40的活性。還有使用氧四環素作為其他治療手段[4],它可以導致VP40相關的分泌小體減少,并顯著降低接受了分泌小體的受體細胞的生存能力,這一成果對治療埃博拉出血熱有著很大的研發空間。
2.5 糖蛋白(GP) GP是埃博拉病毒蛋白中最主要的毒性蛋白。GP并非指單獨的GP糖蛋白,而是由編碼GP基因經過轉錄編輯形成的三種產物的總稱:其包括全長的GP蛋白(包含GP1受體結合域,GP2病毒融合域兩個亞基);可溶性GP(sGP,缺少橫跨膜域)與小可溶GP(ssGP)。sGP上存在福林酶位點,經過福林酶處理后的sGP可以產生一個碎片片段稱為△-肽[14]。
GP與病毒感染細胞的途徑密切相關。通過GP介導的胞飲作用,GP會將病毒與宿主細胞的前溶酶體結構相融合,促使病毒進入細胞。在毒性方面,GP通過黏蛋白類似域影響細胞的絲裂原活化蛋白激酶(MAPK)通路,降低了ERK2的磷酸化作用與催化活性,降低宿主細胞的粘附性,使細胞剝離且不能維持正常的球形,繼而使細胞死亡[15]。此外,sGP的碎片產物△-肽有可能還扮演著埃博拉病毒致病機理中病毒孔蛋白的作用,其可以在哺乳動物的原生質膜上形成孔道,增加離子通透性,破壞細胞的內環境平衡[16]。
GP作為位于病毒衣殼表面的蛋白,與埃博拉病毒的侵染選擇性也有很深的淵源。在2014-2016年的西非埃博拉出血熱爆發中,檢測到的埃博拉病毒的GP蛋白在A82V中的高頻變異。這一變異增加了GP的膜融合活性,并且使得大量其他型的細胞變得可以被侵染:包括黑猩猩的纖維母細胞(S008842)、恒河猴上皮細胞(FRhK4)、非洲綠猴上皮細胞(Vero)以及人樹突細胞。這一信息表明新種埃博拉病毒已經開始高度適應人類作為宿主[17-18]。
2.6 核蛋白(NP) NP是埃博拉病毒的復制循環中的重要蛋白——核糖核蛋白復合物的亞基,其主要功能是保護病毒RNA免于被降解,保證病毒基因組在裝配過程中正確進入到病毒粒子的衣殼中。目前NP的新功能并沒有發現。
2.7 聚合酶蛋白(L) L是依賴RNA的RNA聚合酶,是病毒多聚酶復合物的元件之一。主要功能是病毒的轉錄與復制過程中對病毒RNA進行反轉錄與翻譯,但是L作為一種病毒的RNA聚合酶,其功能與真核細胞的RNA聚合酶差異很大。以GP基因為例子,L將其翻譯的結果是sGP而不是GP[19]。L的翻譯也是實現調節GP、sGP、ssGP不同水平的原因。在連續培養的組織細胞中,L被發現能夠在GP基因中連續的7個U中再添加一個U,從而調節GP與sGP的表達比率為80∶20。在豚鼠的實驗中,該基因回復突變再次使這個基因又變回了7個U,導致GP∶sGP=20∶80[20]。這一特點有可能是病毒進行免疫回避的一種獨特手段。作為對比,在人的肝癌細胞系(Hun7)中的病毒復制則是一個該基因為9U的變異體,其保持了高水平的sGP表達,同時增強的還有ssGP的表達。通過豚鼠與人肝癌細胞的兩個實驗中不難發現L除了翻譯的功能之外,還存在有調節GP與GP相關蛋白比例的功能。這一現象據推測可能與病毒為了適應不同的宿主而進行的調節有關,從而實現在宿主體內的快速復制。
3 結 語
埃博拉出血熱目前尚無有效的治療手段。對埃博拉疫苗的研究始于2004年,由美國衛生與公眾服務部(State Department of Health and Human Services)發起的Project Bioshield,最終于2014年推出了能夠100%保護的疫苗,這為后來的埃博拉病毒疫苗提供了模板。隨后的口腔炎病毒(rVSV)與扎伊爾埃博拉病毒(ZEBOV)糖蛋白的重組病毒(rVSV-ZEBOV)疫苗在2016年的埃博拉疫情中效果顯著,但是仍然有患者在痊愈后1~2年中抗體顯著下降,有著復發的潛在危險[21]。目前來看,針對于埃博拉出血熱的疫苗開發形式仍然嚴峻。
猜你喜歡
鐘表(2023年5期)2023-10-27 04:20:44
中華詩詞(2022年6期)2022-12-31 06:41:24
當代陜西(2021年21期)2022-01-19 02:00:26
中學生數理化(高中版.高考數學)(2020年1期)2020-02-20 13:23:44
經濟技術協作信息(2018年11期)2019-01-14 03:07:20
中國科技論壇(2017年7期)2017-07-25 08:49:53
制造技術與機床(2017年3期)2017-06-23 08:11:33
媽媽寶寶(2017年2期)2017-02-21 01:21:24
國際漢語學報(2016年1期)2017-01-20 08:21:20
中國中醫藥現代遠程教育(2014年22期)2014-03-01 04:32:55
