干旱和復水條件下小麥葉片 TaNCED1表達與ABA積累的關系
2019-05-15 06:21:20宋國琦張淑娟陳明麗李玉蓮張榮志韓小東李根英
麥類作物學報 2019年4期
宋國琦,李 瑋,張淑娟,陳明麗,高 潔,李玉蓮,張榮志,韓小東,李根英
(山東省農業科學院作物研究所,農業部黃淮北部小麥生物學與遺傳育種重點實驗室/小麥玉米國家工程實驗室,山東濟南 250100)
干旱是小麥產量的主要限制因子之一。挖掘和利用優異抗旱基因資源是提高小麥抗旱性的重要手段。研究表明,ABA是植物響應水分脅迫的重要信號分子,在抗旱性方面發揮著重要的調節作用[1-2]。干旱脅迫下,ABA不僅能夠促進氣孔的關閉以降低水分的蒸騰,而且能夠調控一些滲透調節物質基因的表達,從而提高植物對非生物脅迫的適應性[3-4]。在ABA的生物合成途徑中,9-順式-環氧類胡蘿卜素雙加氧酶(NCED)是關鍵的限速酶,能夠調控ABA的合成速度[5-9]。
目前,已在菜豆、擬南芥、鱷梨等植物中獲得NCED基因,并對其功能進行了分析[10-12]。研究結果表明,干旱脅迫處理不僅可誘導NCED基因的表達,同時也提高了內源ABA的活性[13-14]。但是其在不同脅迫或不同物種間的表達模式是不同的[15-17]。在對模式植物擬南芥的研究中發現,NCED為一個多基因家族,共有5個NCED基因,分別為AtNCED2、AtNCED3、AtNCED5、AtNCED6和AtNCED9,其中AtNCED3是應答水分脅迫的主要基因[15-16]。在對鱷梨的研究中發現,PaNCED1和PaNCED3能誘導果實成熟,并且PaNCED1受水分脅迫誘導表達,但是在脫水的葉片中未檢測到PaNCED3的表達[17]。在對小麥的研究中發現,NCED基因在普通小麥及其近緣種中在正常條件下表達量很低或幾乎不表達,但在逆境條件下的表達量明顯增強;脫水脅迫和鹽脅迫都能夠強烈誘導NCED基因的表達,并且在非生物脅迫條件下普通小麥及其近緣種之間NCED基因的表達量具有明顯差異[18]。在轉基因植物中NCED基因的過量表達能引起ABA積累和耐旱性增加[7,19]。上述研究表明,NCED基因在抵御干旱脅迫中起著重要的調控作用。
2012年本課題組成功克隆了小麥NCED1基因的全長,并將其命名為TaNCED1,GenBank登錄號為JQ772528.1,該基因為單拷貝,長度為1 848 bp,無內含子。研究表明,在小麥中TaNCED1基因對脫水、鹽、冷和ABA脅迫均產生不同程度的響應;過表達TaNCED1的轉基因煙草抗旱性顯著提高[19]。但是目前在逆境脅迫下小麥中TaNCED1表達和ABA積累關系的研究尚未見報道。本研究擬通過研究不同小麥品種在干旱脅迫和恢復水分條件下TaNCED1的表達水平和ABA含量動態變化,尋找不同小麥品種中TaNCED1表達與ABA含量變化之間的關系,探討TaNCED1對水分脅迫的響應機理及其在小麥抗旱中的作用,以期為小麥耐旱理論研究及耐旱新品種選育提供理論依據。
1 材料與方法
1.1 植物材料
供試材料為濟麥22、魯麥21、和尚頭、臨旱2號、石4185、濰麥8號、青麥6號和鄭麥366。其中,濟麥22、石4185、濰麥8號和鄭麥366為中高肥水地塊種植的品種,魯麥21、和尚頭和青麥6號為抗旱品種,臨旱2號為耐旱品種。將小麥種子浸水至萌發后,置于4 ℃冰箱春化30 d,將芽苗修剪后移栽至直徑20 cm的花盆中,花盆基質為草炭土,每盆種植6棵,每個材料種植24盆。在人工氣候室的可控環境下生長,設置晝/夜溫度為22/16 ℃,光/暗周期為16/8 h,光照強度300 μmol·m-2·s-1,相對濕度為60%~70%。
1.2 方 法
1.2.1 水分處理
小麥幼苗移栽到花盆后,所有小麥水分管理一致,并保證小麥正常生長的水分需求。移栽15 d 以后(此時小麥處于5葉期),將所有材料澆水至飽和狀態,隨后分為干旱處理和對照兩個組。其中,干旱脅迫處理18盆,期間不再補充水分,并觀察植株的生長表現,同時利用水分測定儀測定花盆內土壤含水量的變化;6盆對照的土壤含水量始終保持在70%~80%范圍內。干旱脅迫處理的第5 d,濰麥8號和鄭麥366的展開葉片開始出現萎蔫,此時各干旱處理小麥土壤水分含量平均都在50%左右。在最早出現萎蔫現象時開始計時,依次分別在第6、12、24、48、72和96 h時對干旱處理和對照小麥進行取樣。取樣時,選擇干旱脅迫處理中表現相近的6盆材料,每次每盆分別取1株材料,并將6株材料不同部位的葉片混合,置于-70 ℃下保存備用;干旱計時96 h后,開始對剩余干旱脅迫處理材料進行復水,分別在復水后的第6、12、24、48、72和96 h時再次對各處理小麥進行取樣,取樣方法同上,將6株材料不同部位的葉片混合后置于-70 ℃下保存備用。
1.2.2 小麥總RNA的提取和cDNA的合成
分別取0.1 g樣品于液氮研磨后,利用UNIQ-10柱式總RNA抽提試劑盒(生工,上海)進行RNA的提取,利用Nanodrop 2000/2000 C分光光度計(Thermo,美國)進行RNA濃度的測定與樣品的定量,利用PrimeScript RT reagent Kit with gDNA Eraser(TaKaRa,大連)進行反轉錄,然后取10×稀釋液為模板進行qRT-PCR。
1.2.3TaNCED1基因的實時熒光定量PCR分析
TaNCED1定量PCR引物為NCED1-P2F/R,以小麥Actin為內參,引物序列如下: NCED1-P2F/R,TCATCGCCGTCCACCAGG/GGGCCG CTCCAGGACATT;TaActin-F/R,GCCACAC TGTTCCAATCTATGA/TGATGGAATTGTA TGTCGCTTC(AB181991)。按照Light Cycler 480 SYBR Green I(Roche,德國)說明書,在Light Cycler?480實時定量PCR儀(羅氏,德國)上進行反應。反應結束后分析熒光值變化曲線和融解曲線。每個反應3個技術重復,采用2-ΔΔCt法分析試驗結果。
1.2.4 ABA含量的測定
ABA含量參照張 煒等[20]的方法測定。
1.3 數據處理
采用Microsoft Excel 2007軟件進行試驗數據處理及作圖,采用DPS 15.10進行顯著性檢驗(Duncan多重比較,P<0.05)。
2 結果與分析
2.1 TaNCED1基因表達水平分析
從表1可以看出,在土壤含水量降至50%之后,TaNCED1基因的表達量隨干旱脅迫時間的延長呈現出先增加后降低的趨勢,但各材料間的變化趨勢不同,其中,濟麥22、青麥6號、石4185、濰麥8號和鄭麥366呈現明顯的單峰曲線,魯麥21、和尚頭和臨旱2號呈現明顯的雙峰曲線;材料間TaNCED1表達量峰值出現的時間也存在明顯的差異,其中,臨旱2號和石4185在干旱脅迫6 h后即達到最高值,濟麥22、青麥6號、和尚頭、濰麥8號和鄭麥366在干旱脅迫12 h時達到最高值,而魯麥21則在脅迫96 h達到峰值。
在復水過程中,除臨旱2號外,其余7個小麥材料的TaNCED1表達均表現為先快速下降,之后緩慢上升的“V”字形變化趨勢,且這7個材料TaNCED1的表達量均在復水24~48 h內下降至最低值,之后緩慢上升;臨旱2號TaNCED1的表達水平則表現出降-升-降的變化趨勢,在復水48 h時達到峰值,之后快速下降。分析水分處理與對照TaNCED1表達水平之間的關系發現,石4185在整個復水過程中始終顯著高于對照;臨旱2號在復水6~72 h內顯著高于對照,在復水96 h時降至對照的水平;濟麥22、青麥6號、魯麥21及和尚頭在大部分復水時間段與對照的差異都不顯著;濰麥8號和鄭麥366在整個復水過程中始終低于對照,在復水12 h后與對照的差異達到顯著水平,在復水96 h時的表達量僅為對照的50%左右。
從整個水分脅迫和復水過程來看,水分脅迫導致小麥TaNCED1的表達水平顯著升高,復水后其表達水平相應下降。表明小麥TaNCED1具有水分快速響應的特點,即水分脅迫能夠誘導小麥TaNCED1的表達。不同材料間,TaNCED1對水分變化的響應快慢程度及其表達水平存在顯著差異。
2.2 ABA含量變化分析
從表2可知,在土壤含水量降至50%左右之后,隨干旱脅迫時間延長,所有品種葉片中的ABA含量均呈現增加趨勢,但達到峰值以后,有的品種緩慢下降,有的品種快速下降然后再升高。不同品種ABA積累量到達峰值的時間也不相同:石4185出現在干旱脅迫計時后第6 h,魯麥21、和尚頭、臨旱2號和濰麥8號出現在第12 h,濟麥22和青麥6號出現在第24 h,鄭麥366出現在第96 h。
在復水過程中,小麥葉片中ABA含量的變化趨勢與TaNCED1的表達基本一致。除臨旱2號外,其余7個品種葉片中ABA的積累量均是先下降,然后再緩慢上升,但ABA總體變化幅度較小。濟麥22和魯麥21在整個復水過程中ABA的積累量始終與對照的水平相當;青麥6號及和尚頭在復水96 h后恢復到對照的水平;其他品種則在多數復水階段都顯著低于對照。
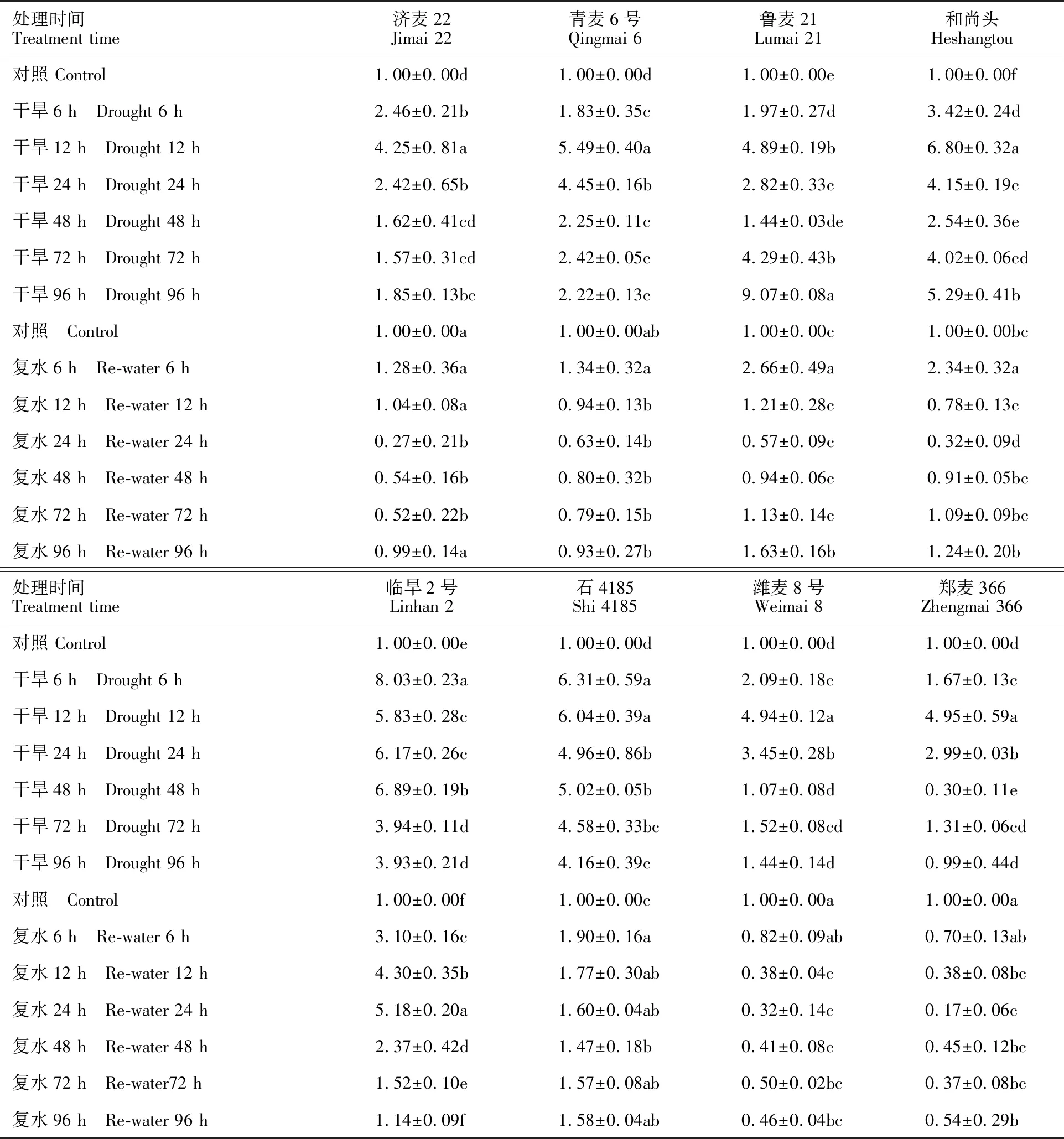
表1 TaNCED1基因在干旱脅迫和復水條件下的表達水平Table 1 Expression level of TaNCED1 gene under drought stress and rehydration
數據用“均值±標準差”表示,同一列數據后的不同字母表示處理間差異顯著(Duncan's多重比較,P<0.05)。下表同。
The data are expressed as “mean value ±standard deviation”, and the different letters within same column indicate significant differences among the different treatments(P<0.05). The same in table 2.
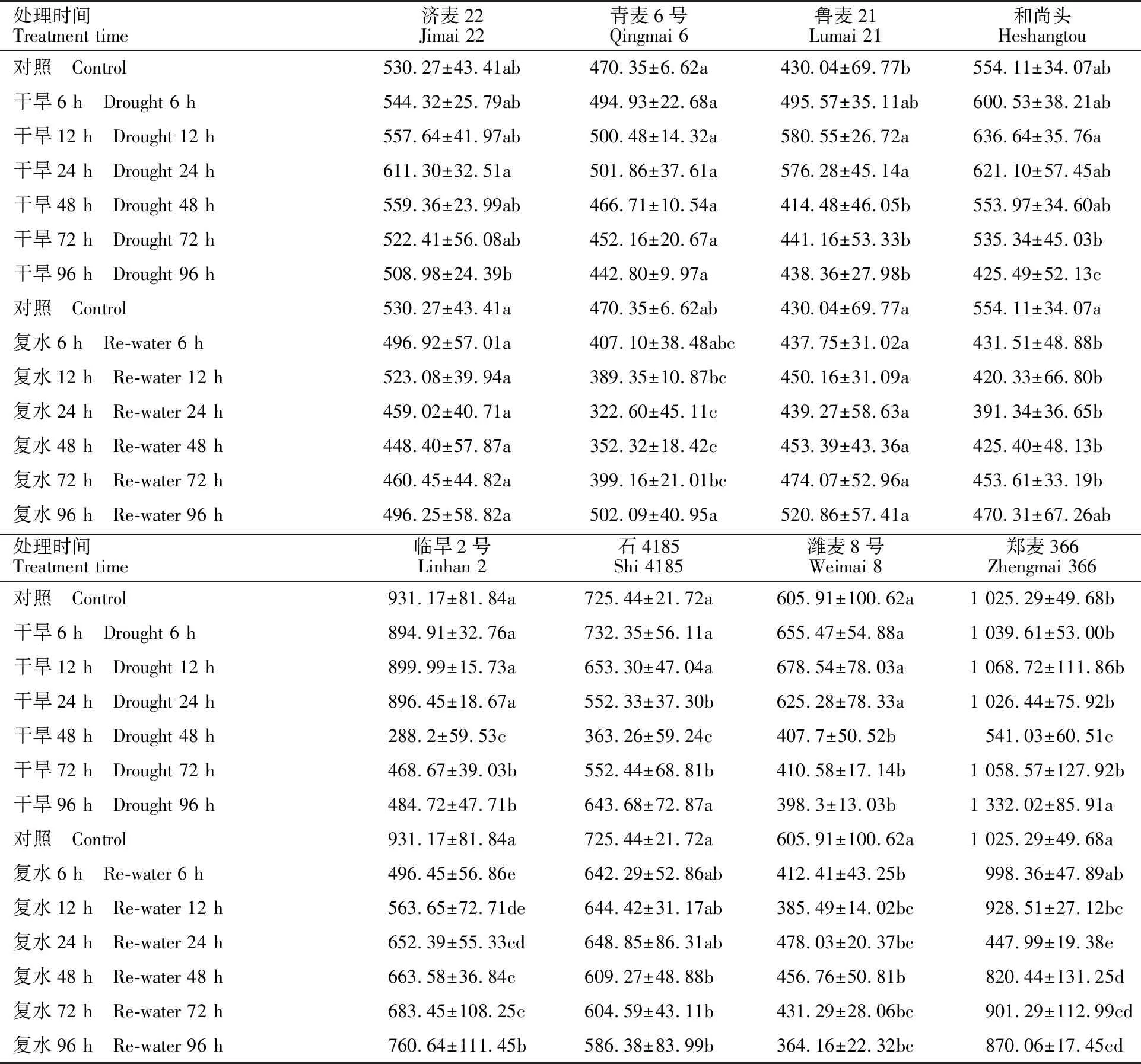
表2 ABA在干旱脅迫和復水條件下的積累情況Table 2 Accumulation of ABA under drought stress and rehydration ng·g-1 dry weight
2.3 TaNCED1表達和ABA積累的相關性
由于小麥中ABA的絕對含量很低,其含量的變化在各處理中表現不夠明顯。鑒于此,在分析中,分別對不同小麥品種各個處理的ABA含量進行了歸一化處理,即將各處理的ABA含量數值分別除以其各自對照的ABA含量值。用相同方法,將各材料不同處理TaNCED1的表達水平也進行了歸一化處理。對歸一化后的TaNCED1表達量與ABA含量進行分析發現,除臨旱2號外,不同小麥品種葉片TaNCED1的相對表達量與ABA相對含量之間均符合y=aln(x)+b的對數相關關系(圖1),系數a表示ABA的相對值隨TaNCED1的相對值變化的變幅大小。相關分析表明,不同小麥品種之間TaNCED1對ABA影響程度的大小有較明顯的差異。
3 討 論
本研究通過分析不同類型抗旱小麥品種在干旱脅迫和復水條件下葉片中TaNCED1表達量和ABA積累量的變化規律及其兩者之間的對應關系,發現水分脅迫能夠誘導小麥TaNCED1的表達和ABA的積累,與水稻[21]、枸杞[22]和煙草[23]中的研究結果一致。但是在水稻和枸杞中發現NCED的表達和ABA的積累是一致的,而在煙草模擬干旱試驗中,NCED3的表達水平在脅迫處理12 h時達到最高值,而ABA的積累量在脅迫72h范圍內隨著脅迫時間的延長持續升高[23]。
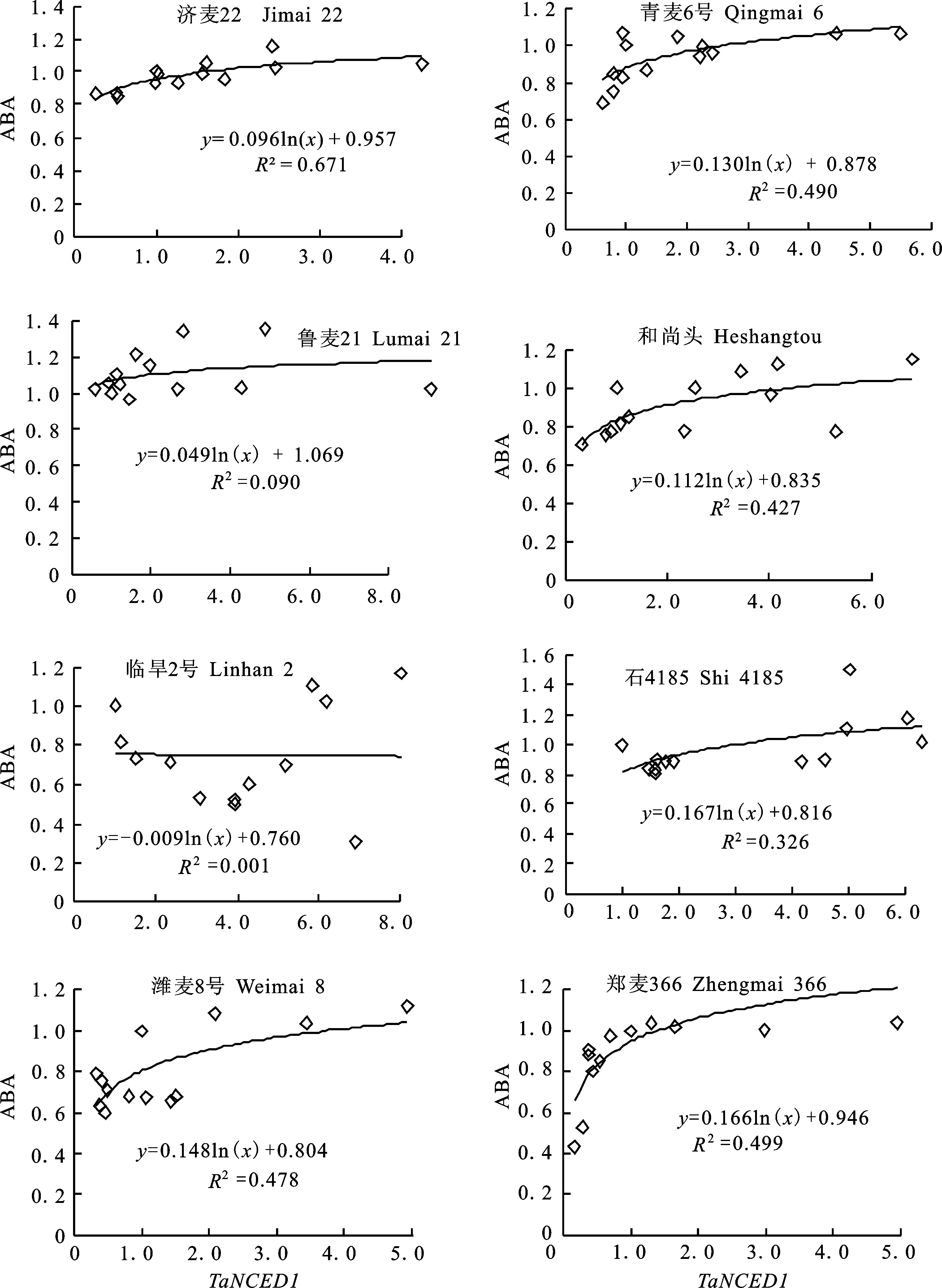
圖1 不同小麥品種在干旱脅迫和復水條件下 TaNCED1相對表達量和ABA相對含量的相關關系
表明不同作物之間NCED的表達和ABA的積累對于干旱脅迫的響應并不是完全對應的關系。本研究發現,除臨旱2號外,其他7個小麥品種ABA的積累量與TaNCED1的表達量均呈對數相關關系。臨旱2號在受到水分脅迫后與其他7個品種的主要差異在于,在水分脅迫48 h時,TaNCED1還在高水平表達,而ABA的積累量卻下降到最低值;在復水過程中,臨旱2號在復水24 h以后TaNCED1表達呈下降的趨勢,而ABA的積累則呈上升的趨勢,這也與其余7個小麥品種不同。臨旱2號為什么和其他材料存在差異?是臨旱2號與其他品種的抗旱機制不同,還是其他因素造成的,本課題組正在從基因序列差異上進行研究。
通過分析干旱脅迫和復水過程中不同小麥品種間TaNCED1表達和ABA積累的變化規律,發現不同品種間TaNCED1對水分變化的響應快慢程度及其表達水平存在顯著差異。NCED基因作為ABA合成間接途徑的關鍵限速酶,其表達水平調控著植物體內的ABA含量[13-14, 24],而ABA在逆境響應中起著重要的調控作用,因此,不同品種間TaNCED1對水分變化的響應快慢程度及其表達水平的差異,是否是導致不同品種抗旱性不同的原因,有待進一步研究。
