華北石質山地側柏人工林C、N、P生態化學計量特征的季節變化
2019-04-17 12:58:12封煥英杜滿義辛學兵張連金孔慶云
生態學報 2019年5期
封煥英,杜滿義,辛學兵,高 旭,張連金,孔慶云,法 蕾,吳 迪
中國林業科學研究院華北林業實驗中心,北京 102300
碳(C)、氮(N)、磷(P)是植物的基本營養元素,參與細胞的結構與功能,與植物體生長和各種代謝過程存在密切聯系[1-2]。生態化學計量學通過分析生物系統多重化學元素和能量的交互作用,為探索C、N、P等元素的生態學過程和生物地球化學循環提供了一種新思路、新手段[3]。陸地植物生態化學計量學的研究起步較晚,但近年來取得了較大的進展[4],在開展不同時空尺度下森林[5]、草地[6]、荒漠[7]和濕地[8]的生態化學計量學研究中發現,植物各器官因養分儲存及功能性分化的差異性[9],可能導致同一植物不同器官間[10-11]及不同物候期下的化學計量特征存在差異[12-13]。
側柏(Platycladusorientalis)為多年生常綠喬木,具有耐干旱瘠薄,適應性強且壽命長的特性,是華北石質山區主要造林樹種之一,廣泛存在于我國北方地區,同時也是北京地區中低山的地帶性植被類型。目前,關于側柏人工林的研究多集中于其生物量及碳儲量[14-15]、水分利用策略與抗旱機理[16]、密度效應[17]、枯落物水文效應[18-19]、養分元素分配及循環特征[20],其生態化學計量學相關研究較少。盡管白雪娟等[21]和張海鑫等[22]對黃土高原地區側柏的“葉片-凋落物土壤”進行了生態化學計量特征和重吸收率的研究,但實驗僅是通過單次采集植物體光合器官完成的,研究亟待拓展至其他器官和整個生長季,且目前關于側柏不同器官養分元素間的相互作用和分配差異尚不明晰。華北石質山地是華北平原的重要生態屏障,屬于半干旱區域,然而該地區土層瘠薄、土壤儲水能力差、巖石滲漏性強,水土流失嚴重;且早期山地植被破壞嚴重,土壤退化顯著[23]。側柏是該區域典型造林樹種之一,對維持石質山區脆弱生態環境起到重要作用。因此,本文以華北石質山地幼齡側柏林人工林為研究對象,系統研究不同器官(葉、枝、根(0—10 cm、10—20 cm))在整個生長季節C、N、P生態化學計量學特征:1)明晰側柏人工林各器官生態化學計量特征的異同;2)揭示側柏人工林各器官化學計量特征在生長季節的變化規律及變異性;3)通過各器官化學計量同步、系統的研究,試圖找出不同器官間化學計量的關聯性。研究旨在解析生長季節內側柏人工林各器官生態化學計量特征,豐富我國植物種群化學計量學內容,為我國生態化學計量學的區域性研究與生物地球化學循環的模型整合提供基礎數據,同時也為指導幼齡側柏人工林經營提供理論支持。
1 研究方法
1.1 研究區概況
研究區位于北京市門頭溝區九龍山自然保護區(39°54′—39°59′N,115°59′—116°07′E),太行山低山丘陵區,海拔100—997 m;屬于暖溫帶大陸半濕潤季風氣候,年均氣溫11.8℃,年均降水量623 mm,主要集中在6—9月;年均蒸發量l 870 mm,無霜期216 d左右;土壤類型屬于山地褐土,土層普遍較薄,石礫含量高。九龍山有著華北石質山區典型的地形地貌、土壤和氣候條件及區域代表性地帶植被類型,九龍山進行封山育林始于20世紀60年代,并且營造了大面積的人工林,構成了以人工森林和灌叢為主的植被類型。目前天然植被以次生灌叢和灌草為主,包括荊條(Vitexnegundovar.heterophylla)、三裂繡線菊(Spiraeatrilobata)、酸棗(Ziziphusjujubavar.spinosa)等灌木和狗尾草(Spiraeaviridis)、黃背草(Themedajaponica)、茜草(Rubiacordifolia)、藎草(Arthraxonhispidus)等草本;喬木以人工林為主,主要包括側柏、油松(Pinustabulaeformis)、栓皮櫟(Quercusvariabilis)、華北落葉松(Larixprincipis-rupprechtii)、樟子松(Pinussylvestrisvar.mongolica)、白蠟(Fraxinuschinensis)、紫丁香(SyringaoblataLindl)等樹種[24]。
1.2 樣地選擇與采樣分析
2016年4月,在北京九龍山自然保護區共設置3個林齡35 a,面積20 m×20 m的干擾程度較小、立地條件一致、具有代表性的幼齡側柏人工林樣地,每個樣地間設置大于5 m寬的緩沖帶,3個樣地共計3次重復。樣地內側柏平均胸徑6.59 cm,平均樹高7.30 m,密度3000株/hm2,郁閉度0.70,土壤表層(0—10 cm)C、N、P含量分別為32.31、2.05、0.65 g/kg,土壤(0—30 cm)中大于2 mm的石礫含量約為40%。依據側柏各項生長指標的平均值,在樣地內確定5株中等大小、長勢及冠幅較一致的健康標準木,在2016年4月至2016年12月,每2個月的中旬利用高枝剪或爬樹收集植物葉、枝樣品,植物樣品分別從樹冠的上、中、下3個層次的4個方向進行采集,每株植株采集12個健康標準枝,每個樣地內共計獲得60個健康標準枝,收集其上的所有針葉和枝,分別混合后作為1個重復樣品。使用根鉆進行植物根系的采集,每個月中旬用根鉆(φ=10 cm)分別在所確定的各標準木根際約50 cm處鉆取0—10、10—20 cm樣木根(粗根與細根的混合樣品),每個樣地5次重復,同一樣地內相同層次混合后作為1個重復樣品。因此,在4—12月生長季節內,分別獲得側柏葉、枝和根系樣品各15份,共計45份植物樣品。
將采集的新鮮葉、枝和根系樣品洗凈裝入信封,于烘箱中105℃下殺青0.5 h,然后在65℃恒溫烘干48 h至恒重,烘干后的樣品用粉碎機粉碎(<0.5 mm),裝入棕色磨口瓶,用于測定葉片的全C、全N和全P含量,其中植物有機碳含量采用重鉻酸鉀-外加熱法測定、全氮含量采用半微量凱氏定氮法測定、全磷含量采用堿溶-鉬銻抗比色法測定[25]。
1.3 數據處理
數據統計分析在SPSS 16.0軟件下完成,正態分布性檢驗采用K-S檢驗(one sample Kolmogorov-Smirnov test)方法。首先對植物葉、枝及根的C、N、P含量及其化學計量比數據進行以10為底的對數轉換,使其符合正態分布,再求算其算術平均值。變異系數(cofficient of variation,CV)用公式CV=標準偏差/平均數×100%計算得出。不同器官,不同月份下數據變量的差異運用One-Way ANOVA分析,LSD進行方差分析。采用GLM模型中的雙因素方差方法分析不同季節和不同器官對植物C、N、P含量及其化學計量比的影響。植物各器官間不同化學計量參數相關性分析采用Peason相關分析。
2 結果與分析
2.1 側柏各器官C、N、P含量及其化學計量比
側柏不同器官間(葉、枝、根)C、N、P含量(圖1)及其化學計量比存在顯著性差異(圖2)。其中,葉、枝、根(0—10 cm)、根(10—20 cm)C含量平均值分別為(446.68±9.46) g/kg、(461.58±8.00) g/kg、(414.97±25.83) g/kg、(417.89±27.97) g/kg,相同生長季節下各器官間C含量差異顯著,均表現為枝最大,葉次之,根最小;N含量平均值分別為(14.28±1.68) g/kg、(6.57±0.63) g/kg、(9.26±1.13) g/kg、(8.05±1.26) g/kg,相同生長季節下各器官間N含量差異顯著,均表現為葉>根(0—10 cm)>根(10—20 cm)>枝; P含量平均值分別為(1.28±0.31) g/kg、(0.72±0.11) g/kg、(0.40±0.08) g/kg、(0.39±0.06) g/kg,相同生長季節下各器官間P含量差異顯著,均表現為葉最大,枝次之,根最小。C∶N平均值分別為31.76±4.39、70.98±7.91、45.50±6.46、53.19±9.61,相同生長季節下各器官間C∶N差異顯著,均表現為葉>根(10—20 cm)>根(0—10 cm)>葉;C∶P平均值分別為369.93±97.05、653.78±98.75、1087.80±301.81、1099.20±213.58,相同生長季節下各器官間C∶P差異顯著,均表現為根最大,枝次之,葉最低;N∶P平均值分別為11.50±1.64、9.21±0.96、23.81±4.81、20.99±4.01,相同生長季節下各器官間N∶P差異顯著,均表現為根最大,葉次之,枝最低。
2.2 側柏各器官C、N、P含量及其化學計量比的動態變化
側柏各器官的C、N、P含量(圖1)及其化學計量比在生長季節均呈現出一定程度的動態變化(圖2)。其中,葉C含量436.07—452.55 g/kg,N含量12.05—15.73 g/kg, P含量0.90—1.51 g/kg。枝C含量459.47—464.74 g/kg,N含量5.97—6.94 g/kg,P含量0.60—0.83 g/kg。根(0—10 cm)C含量392.18—435.02 g/kg, N含量8.76—10.04 g/kg,P含量0.32—0.47 g/kg。根系(10—20 cm)C含量397.67—443.87 g/kg,N含量7.85—8.25 g/kg,P含量0.37—0.42 g/kg。各器官中C元素最穩定(變異系數均小于7%),P元素變異性最大(變異系數均超過15%)。各器官C元素含量隨生長季節變化均不顯著;葉N元素含量隨生長季節變化顯著,表現出先上升,到達峰值后下降,最后趨于平穩的趨勢,而其他器官生長季節內N元素無顯著性變化;葉和枝P元素含量隨生長季節變化顯著,趨勢與葉片N元素相似。
葉C∶N比值范圍27.77—37.41,C∶P為302.32—503.60,N∶P為10.21—13.49,變異系數分別為14%、26%、14%;枝C∶N比值范圍66.56—77.17,C∶P為563.19—771.26,N∶P為8.26—10.01,變異系數分別為11%、15%、10%;根(0—10 cm)C∶N比值范圍39.24—48.67,C∶P為867.21—1421.10,N∶P為21.09—30.27,變異系數分別為14%、28%、20%;根(10—20 cm)C∶N比值范圍50.86—55.99,C∶P為957.03—1228.91,N∶P為18.72—21.85,變異系數分別為18%、19%、19%。同一器官化學計量比在不同生長季節間均無顯著性差異,各器官中C∶N和N∶P較C∶P更為穩定。
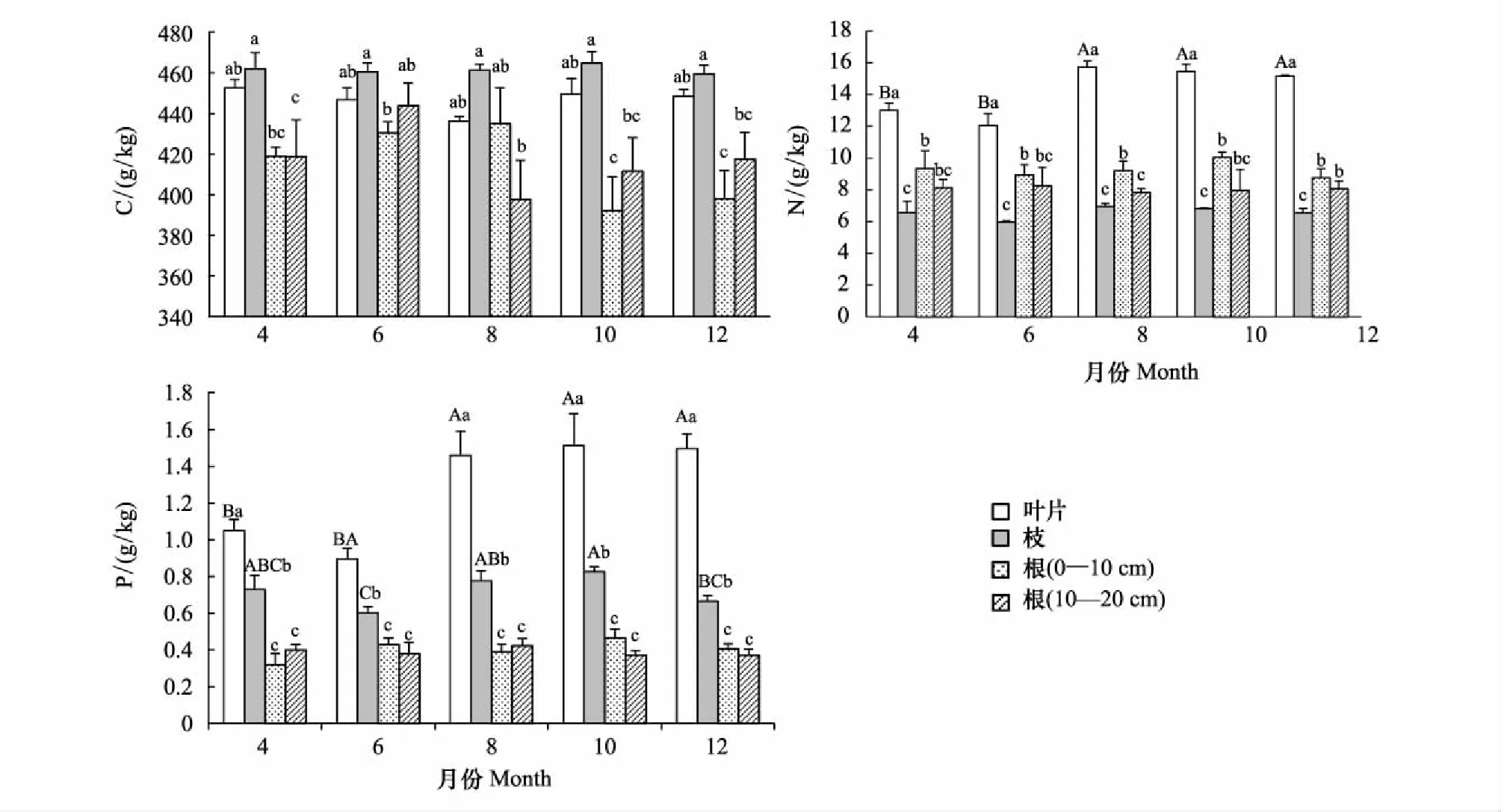
圖1 側柏各器官C、N、P含量及其動態變化(平均值±標準偏差)Fig.1 Seasonal dynamics of C, N, P concentration in different organs of Platycladus orientalis (mean±SD)不同小寫字母表示各器官間差異,不同大寫字母表示各生長季節間差異(P<0.05)
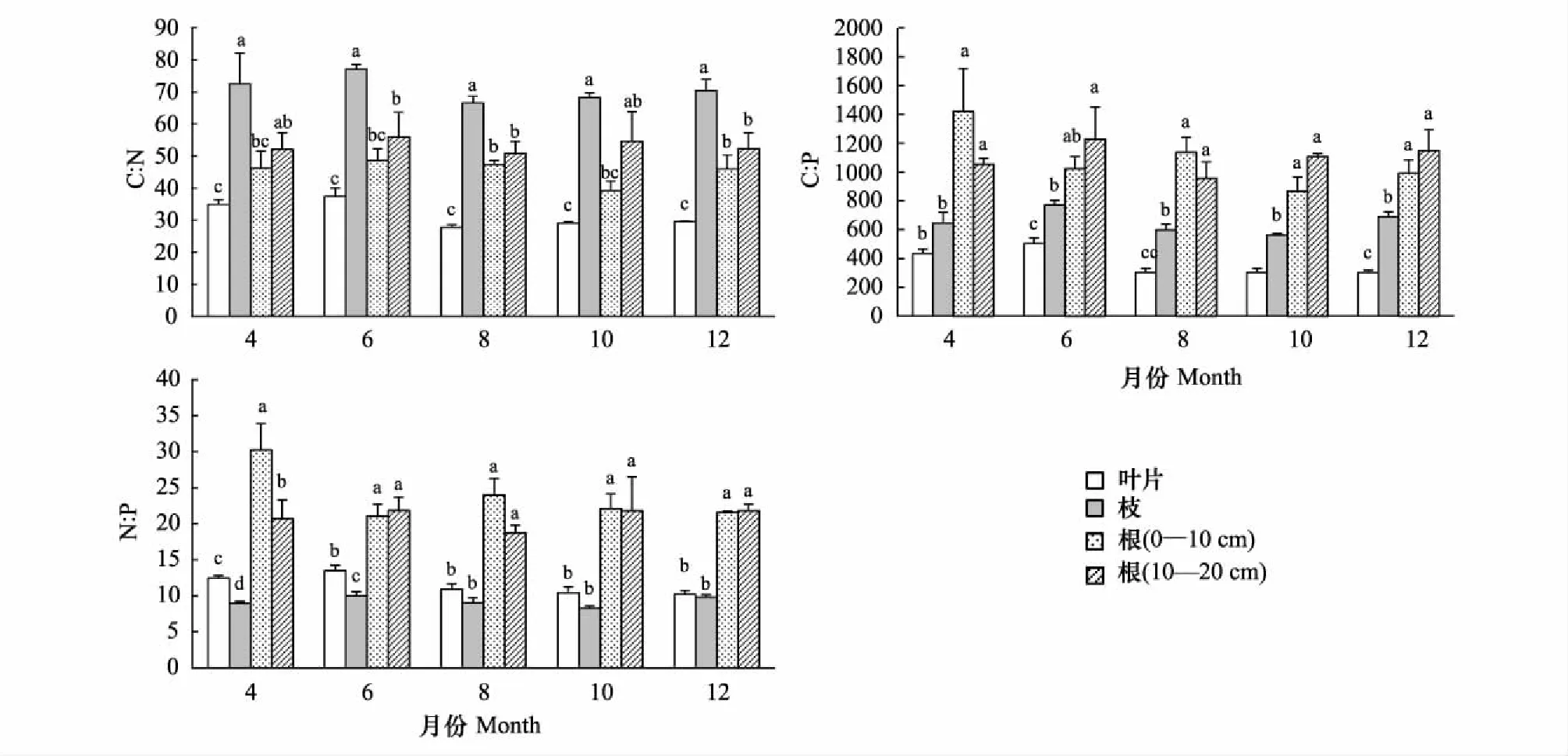
圖2 側柏各器官C∶N、C∶P、N∶P含量及其動態變化(平均值±標準偏差)Fig.2 Seasonal dynamics of C∶N, C∶P, N∶P ratio in different organs of Platycladus orientalis (mean±SD)
2.3 C、N、P含量及其比值間相關性
通過SPSS 16.0 統計分析軟件對側柏植株總體C、N、P含量及其化學計量比值進行Person相關分析,結果表明(表1):C、N含量與P含量總體上均呈現極顯著正相關(P<0.01),相關系數分別是0.402和0.762,而C、N之間相關系數不明顯(P>0.05),這在一定程度上反映出N、P在植物體內存在較高的耦合度。C∶N與P呈現極顯著負相關(P<0.01),這主要是因為N作為C∶N的分母,N越大,C∶N越小,而N和P又正相關,所以C∶N與P呈現顯著正相關;C∶P與N呈現極顯著負相關(P<0.01),與C∶N呈現顯著正相關(P<0.05)。作為C∶P的分子,C與C∶P理論上應為極顯著正相關,然而結果表明C∶P與C為極顯著負相關,這說明在側柏中的C∶P變化主要由P來決定;N∶P與C和P均呈現顯著負相關(P<0.01),與N不存在顯著相關性(P>0.05),這說明在側柏中N∶P的變化主要由P變化決定。

表1 C、N、P含量及其化學計量比之間的相關系數
*,相關系數顯著水平為5% (P<0.05);**,相關系數顯著水平為1% (P<0.01)
2.4 側柏各器官間C、N、P含量及化學計量比相關性
側柏各器官C、N、P含量相關性多數未達到顯著性水平(表2),僅有葉與枝P元素含量顯著相關;器官間化學計量比的相關性多數也未達到顯著性水平,僅有葉與枝C∶P比值顯著相關。
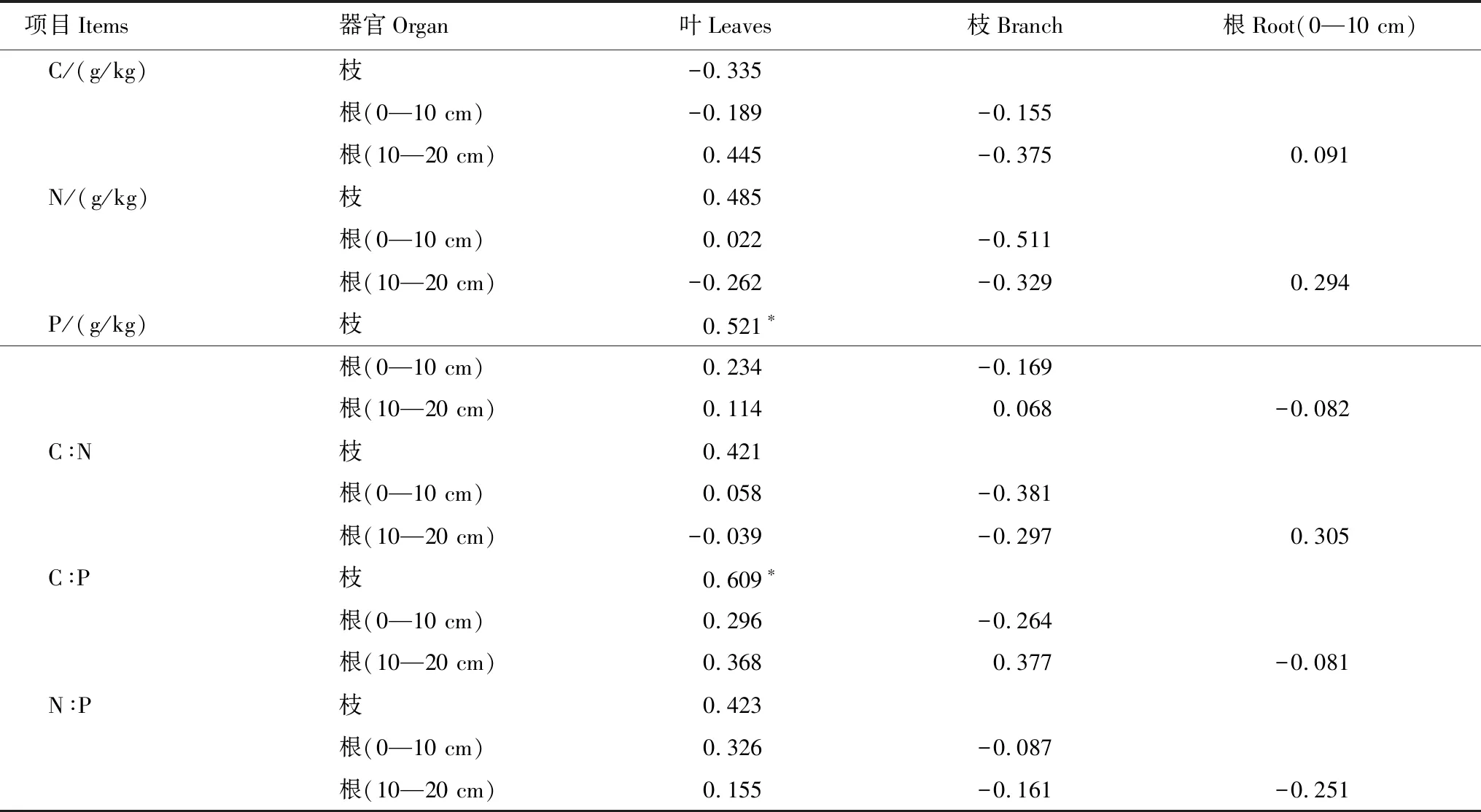
表2 側柏各器官間碳(C)、氮(N)、磷(P)含量及化學計量比的相關關系
2.5 側柏C、N、P含量及其化學計量比的整體變異分析
通過生長季節和不同器官對側柏C、N、P含量及其化學計量關系影響的一般線性模型分析(GLM)發現:不同器官對C、N、P含量及其化學計量關系的影響均達到極顯著水平(P<0.05);生長季節對側柏N含量存在顯著影響(P<0.05),對P含量存在極顯著影響(P<0.01),而對其他化學計量指標影響不顯著;不同器官與生長季節間的交互作用只對P含量存在極顯著影響(P<0.01)。相對于生長季節以及生長季節和不同器官交互作用而言,器官對側柏各化學計量的影響更為顯著(表3)。
表3季節與器官對側柏C、N、P含量及其計量比影響的一般線性模型(GLM)分析
Table3Generallinearmodel(GLM)analysisofeffectsofseasonsandorgansonC,NandPconcentrationsandtheirratiosofPlatycladusorientalis
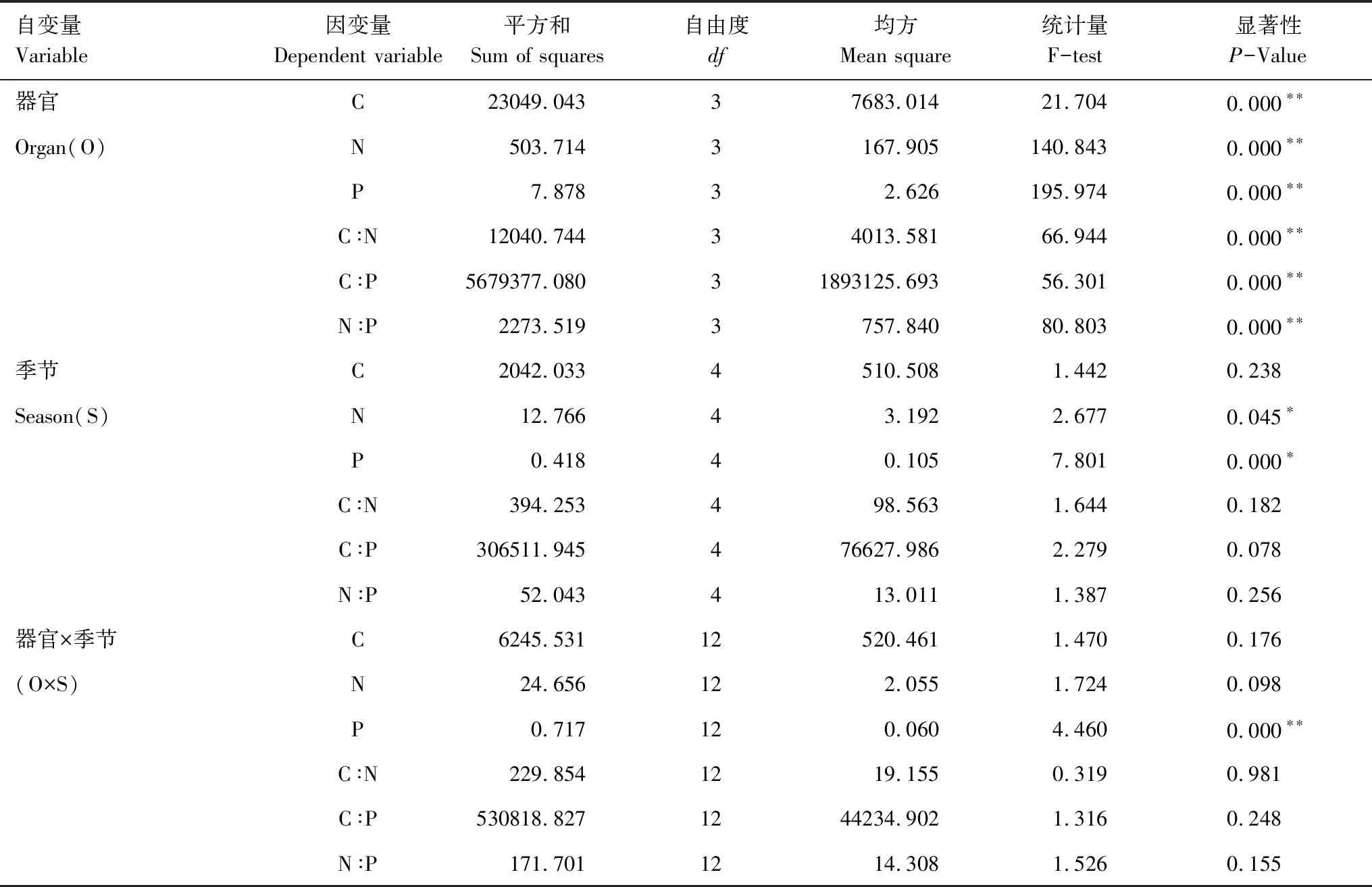
自變量Variable因變量Dependent variable 平方和Sum of squares自由度df均方Mean square統計量F-test顯著性P-Value器官C23049.04337683.01421.7040.000??Organ(O)N503.7143167.905140.8430.000??P7.87832.626195.9740.000??C∶N12040.74434013.58166.9440.000??C∶P5679377.08031893125.69356.3010.000??N∶P2273.5193757.84080.8030.000??季節C2042.0334510.5081.4420.238Season(S)N12.76643.1922.6770.045?P0.41840.1057.8010.000?C∶N394.253498.5631.6440.182C∶P306511.945476627.9862.2790.078N∶P52.043413.0111.3870.256器官×季節C6245.53112520.4611.4700.176(O×S)N24.656122.0551.7240.098P0.717120.0604.4600.000??C∶N229.8541219.1550.3190.981C∶P530818.8271244234.9021.3160.248N∶P171.7011214.3081.5260.155
*,P<0.05;**,P<0.01
3 討論
3.1 側柏各器官化學計量及其整體變異分解
目前森林生態系統化學計量學的研究內容基本上是以植物葉片為研究對象[26]。與已有研究成果對比發現(表4)。
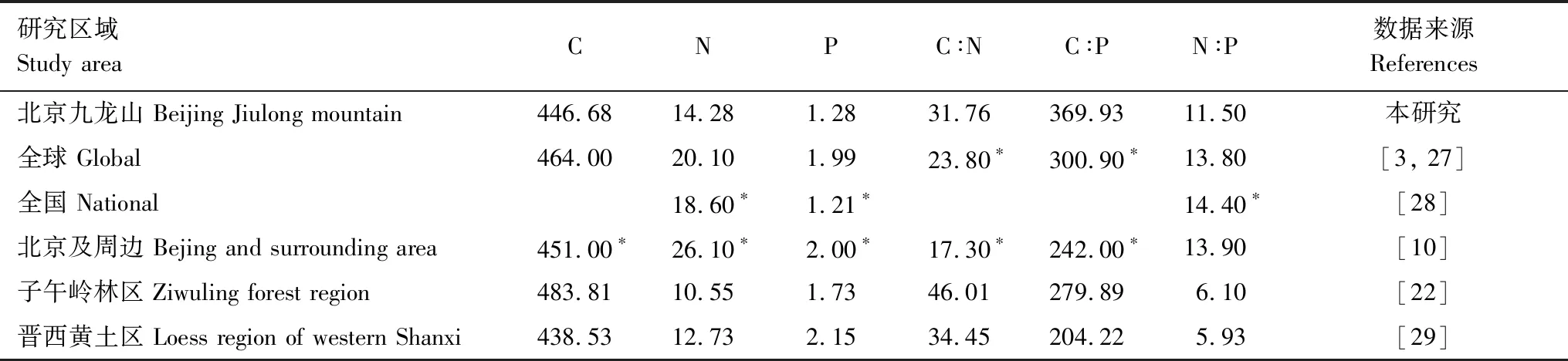
表4 側柏葉片C、N、P含量及其化學計量比與其他已有研究結果的對比
*, 幾何平均值
本研究中側柏葉片C含量為446.68±9.46 g/kg,落在國際上公認的植物平均C含量45%—50%區間內[30],與晉西黃土區側柏葉片C含量相近,而略低于子午嶺林區側柏葉片C含量。側柏葉片N含量為14.28±1.68 g/kg,低于全球、全國及北京周邊范圍內葉片N含量,而高于兩個黃土區域側柏葉片N含量。這一方面因為不同生活型葉片養分含量具有明顯差異,與落葉樹種和闊葉樹種相比,常綠樹種和針葉樹種往往具有較低的N、P含量[31-32],側柏屬于常綠針葉樹種,葉片N含量相對其他樹種偏低;另一方面,在研究對象均為側柏的情況下,對比發現側柏葉片N含量在三地年均降雨量基本相近的情況下隨研究區域年均氣溫增大而增大,即本研究年均氣溫(11.8℃)大于晉西黃土區(10.0℃)和黃土高原子午嶺林區(7.4℃)。“溫度-植物生理假說”(TPPH)認為,植物在低溫下會提升自身元素含量來補償低溫的光合速率[27],顯然本研究并不符合此假設,這是因為植物體攝取的營養元素來源于土壤,其體內N和P元素含量與土壤的N和P可給性有關[33]。本研究中側柏葉片P含量為1.28±0.31 g/kg,與全國陸地植被P平均含量(1.21 g/kg)相近,而低于全球植被P平均值,這是因為與世界其他地區相比,我國陸地生態系統缺磷的現象更為明顯[34]。與黃土高原兩個地區的側柏相比,本研究中側柏葉片P含量偏低,這可能是因為葉片P含量會隨著平均氣溫的降低極顯著的增加,也會隨著經度增大而表現出降低的趨勢[30,35]。本研究中側柏葉片C∶N和C∶P均表現出與N和P含量相反的趨勢,這是因為植被間C含量差異性不大,而C∶N和C∶P的相對大小更取決于N和P含量的高低。本研究區及黃土高原區域的側柏葉片N∶P均小于全球、全國及北京周邊地區植物葉片N∶P平均值,這說明與其他植被相比側柏更大程度上受到N元素的限制;與黃土高原區域相比本研究中側柏葉片N∶P比值較高,說明本區域內葉片中N元素相對充足,這可能與土壤中N、P的含量密切相關。
由于本研究所采集樣品均在同一小地點,取樣地點環境的空間異質性可以忽略不計,這進一步突出了器官和生長季節對植物自身化學計量的影響。對不同器官和生長季節的側柏C、N、P含量及其化學計量比影響的一般線性模型分析顯示(表3),側柏化學計量不僅與器官類型有關,在不同生長季節也表現出較大的差異性。其中,C、C∶N、C∶P和N∶P的變異主要受不同器官的影響;N的變異主要受不同器官和生長階段的影響;P的變異受不同器官、生長階段以及兩者的交互作用的影響;總體而言,側柏C、N、P含量及其化學計量比主要受到不同器官的影響,器官對C、N、P含量及其化學計量比均達到了極顯著水平(P<0.01),這進一步說明側柏器官分化過程中各器官對元素的吸收利用具有特異性。側柏N、P元素的變異同時受到生長季節的影響,除了植物自身生物學特性的原因,還與生長季節中溫度、降雨等環境因子變化有關,夏季高溫多雨,冬季低溫干旱,影響了側柏對N和P的吸收利用。本研究發現,側柏作為多年生的常綠針葉樹種,其化學計量特征在一定程度上受到了生長季節的影響,但相較之下側柏枝條在整個生長季節中C∶N,C∶P和N∶P化學計量特征變異性均最小,且植物樣品相對容易獲取,可以作為側柏不受生長季節影響的化學計量特征,以用于生態系統問題的分析。
3.2 側柏器官間化學計量的差異與關聯
本研究中側柏不同器官(葉、枝、根)C、N、P含量及其化學計量比存在顯著性差異,這種差異不僅受到植物基本生理過程需求的影響,還會受到器官間組織結構和功能分化的影響[9]。葉片是植物同化和代謝的主要器官,植物在生長階段會將大量養分集中傳送給葉片以滿足植物的生長需求[36],而枝和根作為養分的吸收和輸送通道,較少儲存養分,因此葉片中N(14.28±1.68 g/kg)、P(1.28±0.31 g/kg)兩種營養元素含量最高。枝作為支撐和疏導器官,主要由木質素、纖維素等富含C的多糖物質組成,這一結構性質決定其C濃度較高,因此在側柏葉、枝、根中,枝的C含量(461.58±8.00 g/kg)最高,與已有研究結論一致[37-38]。C∶N和C∶P分別為枝和根最大,這主要是由于不同器官間C元素含量差異不大,影響C∶N、C∶P值的主要因素是N、P的含量[27],而枝中N元素和根中P元素含量均明顯小于其他器官;葉和枝的N∶P比值均小于14,根(0—10 cm)和根(10—20 cm)N∶P比值均大于18,即相同發育階段不同器官的C∶N、C∶P和N∶P化學計量特征不同,這與器官的選擇性吸收和養分元素分配格局密切相關。趙一娉等[39]在黃土高陵溝壑區的研究發現森林生態系統喬木層平均C、N、P含量顯著低于葉片水平,葉片不能全面代表喬木層整體養分含量情況,這是由于葉片與其他器官(枝、干、根)化學計量參數不同而引起的,與本文研究結果一致。N∶P是衡量生物體營養狀況和判斷植物群落受養分限制情況的重要指標,根據當N∶P>16表示P限制,N∶P<14表示N限制的判斷標準[40],說明華北石質山區側柏在生長季節里更多受到N元素的限制。目前對植物體限制性元素的判斷多采用葉片為研究對象,這是因為葉片是植物的光合器官,也是最能反映植物生長狀況的器官,但葉片并非唯一、最全面的判斷標準,更多情況下需要拓展至其他器官便于綜合性的判斷,這在后期研究中需要有所突破。此外,N∶P臨界值的閾值受到研究區域、生態系統以及植被類型等眾多因素的影響[41],且一般在肥沃的土壤和某種具有較高養分含量的物種中使用N∶P比值進行養分限制性判斷應當謹慎,相關養分限制診斷指標的敏感性和適應性因研究對象不同而存在差異[34],目前施肥實驗是確定N或P存在限制作用及N∶P臨界值的最佳辦法[42]。
植物各器官由于養分儲存及功能差異性導致同一植物不同器官間的C、N、P化學計量存在顯著差異,但器官間的C、N、P濃度密切相關,且不同器官對養分的利用具有一定相關性[43]。以往更多基于大尺度的研究表明,植物不同器官間功能性具有一致性,即不同器官間N、P含量和N∶P一致相關,且這種相關不隨物種、土壤等因子的變化而變化[44-45]。在種群和物種水平上,研究表明溫帶優勢草本植物間N、P含量以及N∶P在各器官之間一致呈顯著正相關[11]。本研究中葉與枝的P元素含量以及C∶P比值呈現顯著正相關,其他器官間化學計量相關性不明顯(表2)。這表明葉與枝之間的養分利用效率是最緊密聯系的,這是因為實驗所采集到的枝、葉位于同一枝干上,枝作為葉最相鄰的養分輸送通道,其汲取和吸收的養分是同步的[46]。周紅艷等[2]研究發現單葉蔓荊各器官的化學計量相關性主要集中在地上部分,其中尤以枝條和匍匐莖之間表現明顯。葉片和根作為植物地上和地下最重要的營養器官,在功能上聯系緊密,頗受關注。Yuan等[47]發現陸地植物根與葉N∶P相似,但葉片N、P含量絕對值大于根系N、P含量絕對值。本研究中葉與根化學計量未達到顯著性相關,一個可能是因為土壤與根系直接接觸,植物通過根系從土壤中吸收和利用有效的N、P,土壤理化特性對根系生態化學計量特征產生重大影響[48],另一個可能的原因是尺度效應,在小尺度上植物生境條件相對一致,種內個體間功能性狀因變異幅度較小而關聯不明顯[49]。
3.3 側柏各器官化學計量的季節動態
本研究中側柏各器官的C、N、P含量呈現出一定程度的動態變化,其中C在各器官的含量高且相對穩定(變異系數均小于7%),N、P含量較低且變異系數均較大。原因可能是C的轉化主要通過植物光合作用,C是構成植物骨架的基本結構物質,且為植物新陳代謝、生長發育和繁殖等生理活動提供能源的物質,需求量大,所以在植物體內含量高且變異小[37]。葉片中N元素以及葉片和枝的P元素含量均隨生長季節變化顯著,表現出先上升,到達峰值后下降,最后趨于平穩的趨勢。這是因為葉片作為主要的光合器官能夠合成大量的蛋白質和核酸,在生長初期,植物葉片生長速度緩慢,細胞分裂能力一般,所需形成的蛋白質和核酸較少,因此對N、P的選擇吸收較少;在生長旺季時期,需要大量的蛋白質和核酸來滿足植物逐漸增長的生長速率,從而對N、P的選擇性吸收較多;在生長末期,葉片生長基本停止,對葉片N、P營養元素的需求又略有降低并逐漸趨于平穩。對草本[50-51]和灌木[52]的相關研究認為,在植物生長最旺盛的時候對N、P元素的需求最大,但由于葉片生物量迅速增加,植物對營養的吸收趕不上細胞膨脹的速率,致使各器官元素被“稀釋”,最終表現為N、P元素含量下降,與本文得出結論部分上不一致,這可能是因為相對于草本和灌木而言,針葉樹側柏屬于慢生樹種,生長周期長,葉片生長速率相對緩慢,即使在一年中的生長旺季,N和P元素也不會因細胞分裂的“稀釋作用”而降低,這一現象在同為針葉樹的落葉松研究中也得到了印證[12]。
本研究中側柏各器官C∶N、C∶P及N∶P化學計量隨生長季節變化均存在一定變異性,總體而言,C∶N和N∶P較C∶P更為穩定。一方面是因為C元素相對穩定,C∶N、C∶P的季節變異主要由N和P元素決定,而P元素的變異性大于N元素,所以C∶P變異大于C∶N的變異;各器官N∶P比值相對穩定,說明N和P在植物體具有相對一致性(r=0.762**),但N∶P的變化主要取決于P的變化,這是因為N∶P比值與N元素相關性不明顯,而與P元素極顯著負相關(r=-0.687**)。葉與枝的C∶N、C∶P比值在溫度偏低的月份相對較高,這是因為溫度能夠影響植物葉片的N、P元素含量,低溫降低了土壤微生物活性,從而降低土壤中有機物的分解和養分釋放速度;此外低溫還會阻礙養分元素的運輸和植物對養分元素的更新[43,53]。根系(0—10 cm)C∶N、C∶P比值4月均較高,這是因為4月份溫度相對較低,根系生理活動剛剛開始,對土壤中N、P等營養元素的吸收處于起步階段,供應相對不足,因此導致低溫環境下植物根系養分含量偏低而C∶N、C∶P較高的結果。研究還認為較高的C∶N和C∶P同時代表對N、P的利用率較高,以往研究也證實了植物在養分元素比較短缺的情況下,往往具有較高的養分利用效率和再吸收率,是對貧瘠環境狀態的一種生存策略[54-55],本研究中側柏在低溫生長季節中更易發生養分元素限制和短缺的情況,因此,無論是地上部分的枝葉、還是地下部分的根系均可能通過提高N、P利用效率的方式來滿足自身養分的需求[56]。器官生態化學計量學特征隨季節更替而不斷變化,研究認為一次性采樣,階段性的采樣不能夠將植物的生態化學計量學特征真實的反應出來,全面的、跨時空的采樣將更加科學、合理[50,57],這一方面是由于植物自身的生物學特征造成;另一方面是由于植物不同生長階段所處溫度、降雨等環境因素不同,例如:雨水沖刷造成植物葉片N、P元素的流失,土壤水分影響土壤有效養分的釋放與轉移等[52]。
植物在漫長的進化過程中,為適應環境的變化形成了多樣的生活史對策,當外界環境或者種內關系發生變化時,植物為了適應養分限制情況,從而調節不同器官中的養分含量變化及化學計量比值,其生存策略和方式也將發生相應變化。針對幼齡側柏人工林C、N、P生態化學計量特征的季節變化規律,建議在幼齡側柏經營管理過程中,理想條件下可以在N、P元素需求量達到峰值之前適當添加N肥和P肥,尤其是限制性元素N素的添加,以促進林分快速健康生長。本研究對側柏不同器官的C、N、P生態化學計量生長季節變化進行了較為深入的探討,但實際上“植物體-凋落物土壤”連續體構成了一個密不可分的系統,后期研究中需要在此基礎上進一步分析凋落物和土壤的化學計量才能夠更全面的揭示養分元素的傳遞與調控機理。此外,全球變化導致陸地植被元素平衡和計量指標發生重新匹配,有序開展側柏生態化學計量對氣候變化(氮沉降、干旱、溫度升高等)的響應和適應機制以及持續的定點觀測是今后的研究重點。
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
華人時刊(2019年13期)2019-11-17 14:59:54
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
科技知識動漫(2017年5期)2017-05-11 21:34:16
科技知識動漫(2017年4期)2017-04-15 22:24:55
爆笑show(2016年7期)2017-02-09 09:36:13
科技知識動漫(2017年2期)2017-02-06 20:59:46
科技知識動漫(2016年10期)2016-10-18 20:35:00
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
