廣西會仙濕地不同植物根際細菌群落結構及多樣性研究
2019-04-09 05:57:26凃月李海翔姜磊董堃王敦球
生態環境學報 2019年2期
凃月,李海翔,姜磊,董堃,王敦球
桂林理工大學環境科學與工程學院,廣西 桂林 541006
巖溶濕地指主要分布在巖溶地區(包括地表、地下),或以巖溶水為主要補給水源,具有巖溶地區特有的富鈣偏堿性水土特征和典型巖溶水土循環演化機制,以喜鈣耐堿的濕地生物群落為主或與喜鈣耐堿的生物群落相互依存為特征的內陸濕地,包括巖溶濕地的地表或地下的湖泊、沼澤、河流或其他巖溶地下水文系統,但其規模小、數量少、穩定性差、破壞后難以恢復(蔡德所,2012)13。廣西會仙濕地作為低中海拔中規模最大的巖溶濕地,其自然資源豐富,是巖溶地區少見的物種庫,被稱作漓江流域的“腎臟”,對涵養水源、氣候和環境改善、水質凈化、生物多樣性保護以及漓江蓄洪補水等方面發揮重要的生態作用(Ghermandi et al.,2014)。由于周邊污水排放、河道占用,會仙濕地逐漸萎縮,近40年來濕地面積減少了27 km2,生態功能嚴重退化,使生態系統受到嚴重威脅(林棟材等,2008)。
一般認為,濕地系統的凈化功能是利用植物-土壤-微生物的綜合作用實現的,水生植物通過與根際微生物的相互作用完成污水凈化,其中微生物是對污染物進行吸附和降解的主要生物群體和承擔者(Liang et al.,2003;Liang et al.,2017)。研究(Münch et al.,2007)發現,植物根際微生物在污水凈化過程中比濕地植物發揮的作用更大。越來越多的研究表明,在濕地系統中微生物種群的多樣性、穩定性對維護濕地生態系統的平衡具有關鍵作用(De-Bashan et al.,2004;武鈺坤等,2012)。不同植物根際泌氧能力和根際分泌物性質不同,在一定程度上影響微生物的種類和多樣性,而微生物的活性、多樣性與污水處理過程緊密相關,表現為根際微生物群落結構多樣性越高,濕地系統的凈化效果和穩定性越好(魏成等,2008;Zhou et al.,2015)。細菌是微生物的重要組成部分,對不同根系分泌物的利用率和敏感度遠高于其他微生物(真菌、放線菌),能夠與植物根系并存(李潞濱等,2008),在根際微生物中占主導地位。高通量測序技術從整體水平上研究微生物種群結構,真實、準確地反映環境中細菌群落的多樣性和復雜性,具有低成本、高通量、流程自動化等優勢,被廣泛運用于植物根際細菌群落結構特征研究中(李慶崗等,2012;Ligi et al.,2014;Cao et al.,2017;吾爾恩·阿合別爾迪等,2017)。
目前,大部分研究主要集中在沉積物和土壤細菌群落結構等方面,而對于不同植物根際細菌在低溫下多樣性和豐富度研究較少。低溫條件下,植物地上部分枯萎甚至死亡,使得微生物發揮重要降解作用,而微生物的數量、多樣性和群落結構在很大程度上影響污染物的去除。本研究以會仙濕地中分布較廣的單一優勢植物群落為研究對象,采用Illumina HiSeq 2500高通量測序技術,研究南方低溫條件下根際土壤細菌群落組成和變化,以豐富喀斯特濕地植物根際細菌多樣性,為低溫環境下濕地生態系統的結構和功能研究提供依據。
1 材料與方法
1.1 樣品采集和前處理
會 仙 巖 溶 濕 地 ( 25°01′30″- 25°11′15″N ,110°08′15″-110°18′00″E,海拔 118 m)氣候屬于中亞熱帶季風氣候,全年月平均溫度為8.6-28.9 ℃,年平均降雨量為 1890.4 mm,主要集中在 4月、5月,植物群落類型以挺水植物群落(60.45%)和沉水植物群落(9.7%)為主,植物種類較多,且生長茂盛,蓋度常可達 80%-95%,以單一優勢種群較多,主要群落有蘆葦、華克拉莎、長苞香蒲、苦草、五刺金魚藻等(蔡德所,2012)21,24,133,139-143。研究表明(Gopal,1999),選擇當地優勢植物,突出生物多樣性,是提高濕地凈化能力和保護的關鍵措施。故以會仙濕地中典型的單一優勢蘆葦、苦草、華克拉莎和美人蕉植物群落為研究對象,于 2017年 10月10日(秋,水溫18-23 ℃)和2017年12月27日(冬,水溫7-11 ℃)進行采樣(圖1),采樣點選擇在該河段植物生長優勢且種群單一、覆蓋度高的位置。每次采樣共15個,為了減少誤差,有3組重復樣,取水樣后于同一位置取植物根際土樣。采集水樣時,先潤洗1遍采樣瓶,然后連同瓶蓋一同放在水面10 cm下,打開瓶蓋避免水中懸浮物進入瓶中,采集完后確保瓶中無氣泡,送回實驗室盡快測定水質指標。植物根際土壤取樣時,采用“抖根法”,先將整個植物根際部分挖出,抖掉與植物根際松散結合的土壤,放入無菌封口袋中,一部分冷凍保存在冰盒里,帶回實驗室于無菌操作臺上將根系0-4 mm 范圍的土壤放入無菌離心管中,-80 ℃保存,用于測定植物根際土壤細菌種群結構(劉波等,2010);另一部分土樣自然風干,磨細過60目和100目篩,用于土樣理化指標的測定;底泥取樣時,使用底泥采樣器采集0-10 cm的土壤作為底泥樣。
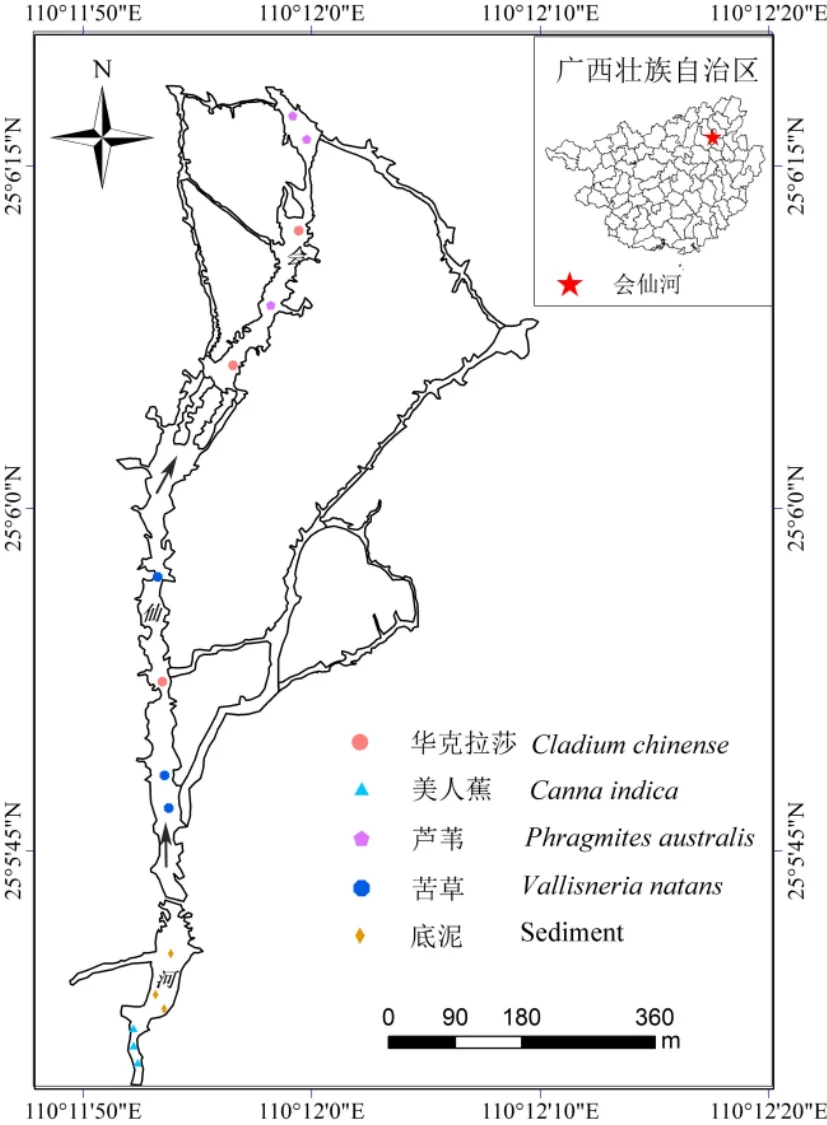
圖1 采樣點位置Fig. 1 Sampling location
測定指標與方法:水質總氮測定方法為堿性過硫酸鉀消解紫外分光光度法,總磷測定方法為鉬酸銨分光光度法。根際土壤總磷測定方法為堿熔-鉬銻抗分光光度法,全氮測定方法為凱氏法,有機碳測定方法為重鉻酸鉀氧化-分光光度法,pH測定方法為電位法。
1.2 植物根際土壤DNA的提取和PCR擴增
采用CTAB法對樣本的基因組DNA進行提取,然后通過1%的瓊脂糖凝膠電泳檢測DNA的質量和濃度,用無菌水稀釋樣品濃度至1 ng·μL-1。以稀釋后的基因組DNA為模板,采用細菌16S rDNA基因 V4+V5 擴增區域,引物 515F(5′-GTGCCAGCMG CCGCGGTAA-3′)/907R(5′-CCGTCAATTCCTTTG AGTTT-3′)。反應條件是:98 ℃預變性 1 min;98 ℃變性10 s,50 ℃退火0.5 min,72 ℃延伸0.5 min,循環30次;然后72 ℃延伸5 min。使用2%的瓊脂糖凝膠電泳檢測擴增產物。擴增過程使用高效、高保真酶以保證擴增過程的準確性(DNA提取、PCR擴增、測序由北京諾禾致源科技股份有限公司完成)。
1.3 文庫及系統發育
PCR擴增產物經過純化回收后,使用 TruSeq?DNA PCR-Free Sample Preparation Kit試劑盒進行文庫構建,然后采用Illumina HiSeq 2500進行高通量測序。所得序列去除 Barcode和引物序列后,使用FLASH(Magoc et al.,2011)和 MUSCLE3.8.31(Edgar,2004)對樣品進行拼接、過濾、多序列對比。
1.4 數據分析
利用Uparse軟件(Edgar,2013),在97%相似性下,對樣品有效數據進行 OTU序列聚類。采用Mothur法(閾值0.8-1.0)和SILVA(Wang et al.,2007)的SSUrRNA數據庫(Quast et al.,2013)對OTU代表序列進行物種注釋分析,得到樣品在各分類水平上的群落組成。使用Qiime 1.9.1計算測序深度 ACE指數、豐富度指數(Chao1)、多樣性指數(Shannon)。不同分類水平上物種組成柱狀圖用Origin.Pro 9.0繪制,可以直觀了解群落組成情況。數據分析于SPSS 19.0中進行,采樣圖用Arcgis 10.2繪制。
2 結果與分析
2.1 采樣點水質及土壤的理化性質
濕地中污染物來源主要是生活廢水的排放,導致水中N、P含量偏高。由表1可知,土壤pH為6.95-7.86,水質 DO 為 4.13-12.90 mg·L-1。不同植物間的理化指標差異顯著(P<0.05),同種植物根際土壤的理化指標呈現相同的變化趨勢,例如,冬季各植物根際土壤中TN、TP、TOC大多高于秋季。水質TN平均質量濃度表現為冬季(2.27 mg·L-1)高于秋季(2.09mg·L-1),超出地表水Ⅴ類標準(GB 3838—2002);而水質 TP 表現為秋季(0.19 mg·L-1)高于冬季(0.16 mg·L-1),超過Ⅱ類標準。沿著水流方向,水中N、P有逐漸降低的趨勢,說明濕地系統對N、P有凈化作用。
2.2 群落結構分析
2.2.1 在門分類水平上
采用 97%相似水平作為標準界定 OTU(張健等,2010),對OTU代表序列進行聚類和物種注釋,共檢測出細菌有65門、160綱、219目、391科、677屬和 246種,細菌群落具有較高的多樣性。由圖 2可知,所檢出的細菌由10個主要門類組成,包括變形菌門(Proteobacteria)、綠彎菌門(Chloroflexi)、酸桿菌門(Acidobacteria)、浮霉菌門(Planctomycetes)、擬桿菌門(Bacteroidetes)、硝化螺旋菌門(Nitrospirae)、藍藻門(Cyanobacteria)、放線菌門(Actinobacteria)、厚壁菌門(Firmicutes)和迷蹤菌門(Nitrospinae)等。在秋、冬兩季不同植物根際土壤中,變形菌均為第一優勢類群,所占范圍是18.6%-49.5%,主要包括4個亞類:γ-變形菌( Gammaproteobacteria ) 、 β- 變 形 菌(Betaproteobacteria)、δ-變形菌(Deltaproteobacteria)、α-變形菌(Alphaproteobacteria),其所占的比例范圍分別為5.4%-12.9%、1.9%-19.2%、1.8%-18.1%、1.9%-8.8%,其中,以β-變形菌和δ-變形菌為優勢菌綱。綠彎菌門占 2.8%-21.6%,為群落中第二大優勢類群,且大部分屬于厭氧繩菌綱(Anaerolineae)。酸桿菌門占 1.8%-12.4%,為群落中第三大優勢類群,其主要是由全噬菌綱(Holophagae)和Subgroup_6組成。相比而言,秋季美人蕉中主要的細菌群落有較大區別,主要是由厚壁菌門(29.0%)、放線菌門(26.1%)、變形菌門(18.6%)以及藍藻菌門(15.0%)組成,且大部分屬于桿菌綱。
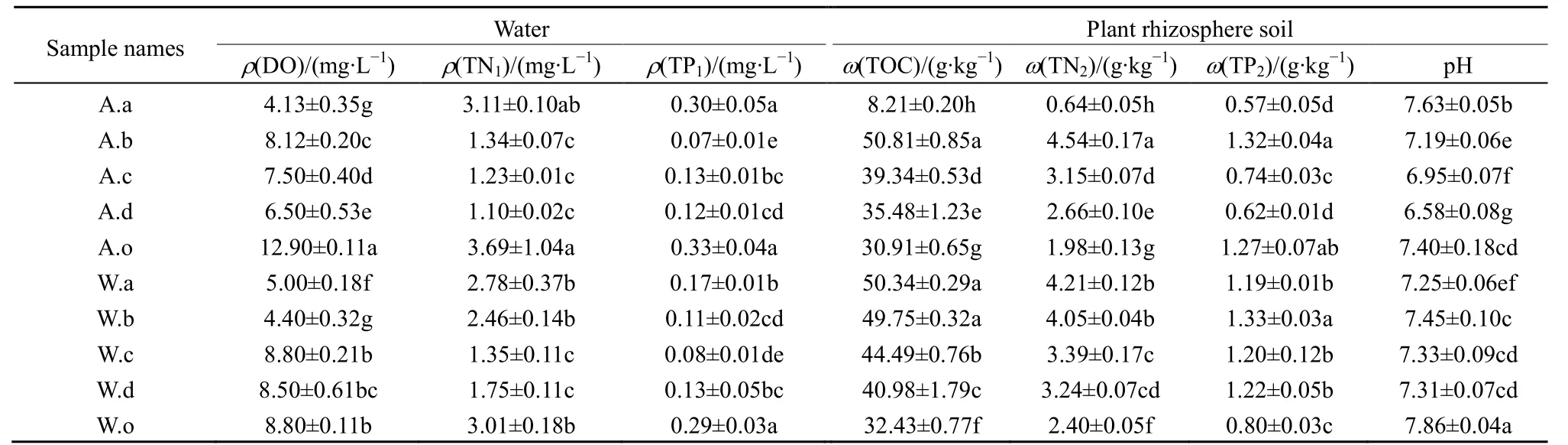
表1 采樣點水質及植物根際土壤理化性質Table 1 Physicochemical parameters in water and plant rhizosphere soil
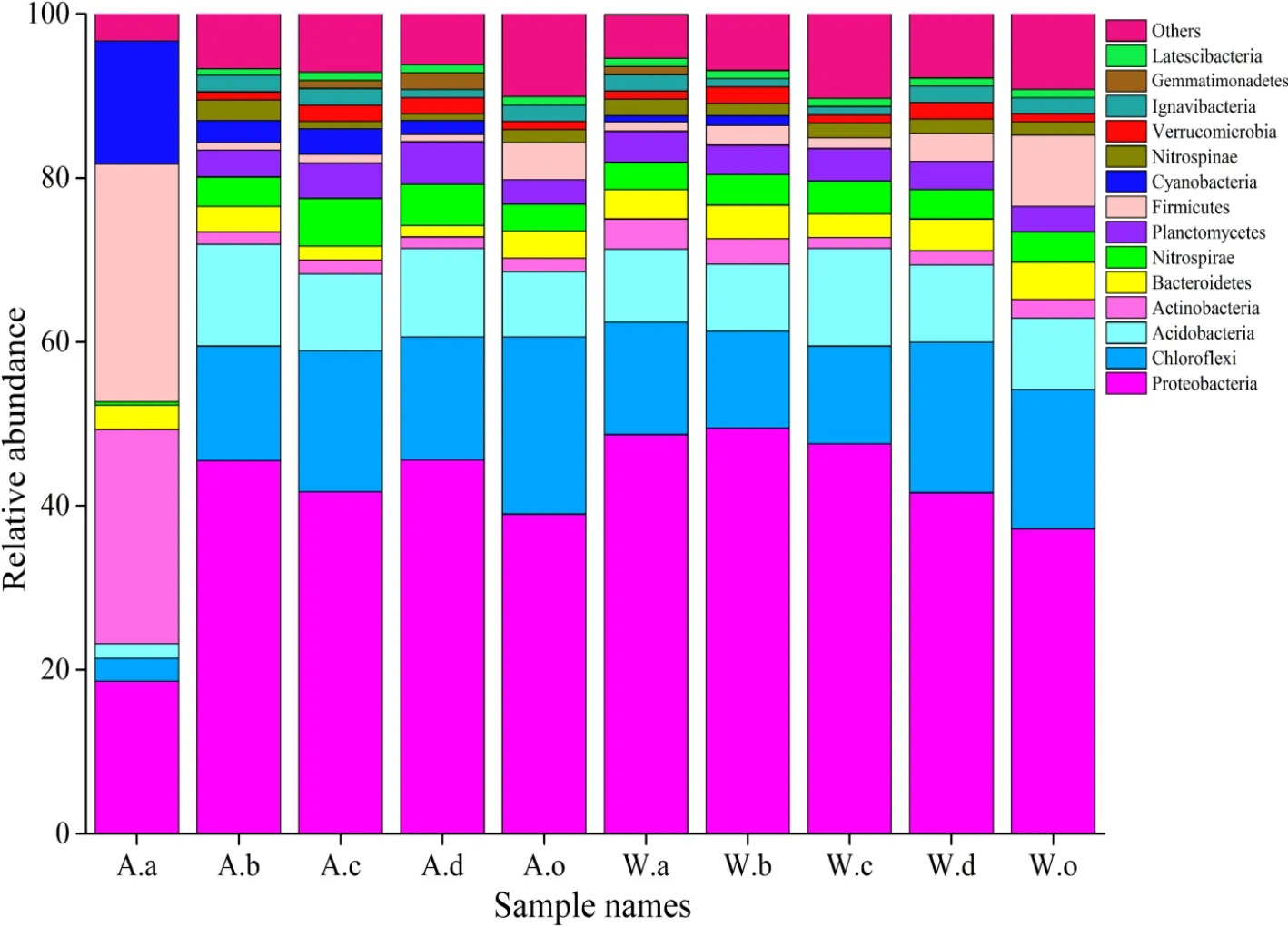
圖2 門分類水平上細菌群落組成Fig. 2 Classification of bacterial sequencing at level of phylum
從季節變化上看,占優勢的群落無較大區別,還是以變形菌門、綠彎菌門為主,但在組成數量上有所區別。變形菌在冬季所占的平均比例(45.1%)高于秋季(38.0%),其中美人蕉中變形菌組成差異最大,秋季為18.6%,冬季為48.7%;而綠彎菌門在華克拉莎中區別較大,秋冬季分別為 17.2%、11.9%。同一植物不同季節有新微生物出現,但其所占比例很小,如,冬季美人蕉中出現的深古菌門(Bathyarchaeota、0.3%)、Parcubacteria門(0.03%)等在秋季美人蕉中均未檢出。柔膜菌門(Tenericutes)僅不存在于美人蕉根際環境,而在其他樣中均可檢測到。與植物根際土壤相比,秋、冬底泥中主要門類細菌群落組成差異最小,說明植物根際對環境中微生物群落分布有影響。總之,不同植物根際土壤細菌中占主導的種群無較大差別,但在數量上有區別。
2.2.2 在屬分類水平上
為了解屬水平物種進化關系,將豐度前100屬的代表序列構建系統發育樹(圖3)。結果表明,組成前100屬的菌有15門,主要是變形菌(51.0%)、放線菌(12.0%)、厚壁菌(7.0%)。其中,豐度較高且存在于大多數樣品中的有硫桿菌屬(Thiobacillus)、厭氧粘菌屬(Anaeromyxobacter)、H16屬、脫硫酸鹽橡菌屬(Desulfatiglans)、地桿菌屬(Geobacter)、Candididatus_competibacter屬(聚磷微生物)、Syntrophorhabdus屬、芽孢桿菌屬(Bacillus)等。從圖可知,芽孢桿菌屬(占所有屬的 28.0%)、紅球菌屬(Rhodococcus,18.0%)、unidentified_Chloroplast屬(15.0%)在秋季美人蕉中的豐度顯著高于其他植物,冬季美人蕉中占優勢菌沒有秋季明顯,屬水平上分布較均勻,占總屬的0.01%-1.2%。不同植物(除秋季美人蕉)共有的優勢菌屬為芽孢桿菌屬(0.3%-28.0%),其次為厭氧粘菌屬(0.8%-2.2%)。總之,除秋季美人蕉中優勢菌群明顯外,其他9個樣各菌在屬水平豐度較低,分布較均勻,秋季美人蕉中Unidentified bacteria占20.2%,說明該植物根系土壤中還存在大量未知細菌,有待進一步研究。
2.3 細菌群落多樣性和環境因子相關性
運用 ACE、Chao1、Shannon指數對不同植物根際土壤細菌多樣性進行分析(表2)。結果表明,各樣本中物種覆蓋率在95.4%-97.8%之間,能夠反映植物根際細菌群落的真實情況。通過分析發現,不同植物、不同季節根際土壤細菌多樣性存在顯著差異(P<0.05),綜合而言,冬季苦草指數最高、秋季美人蕉最低,表明冬季苦草細菌豐富度和多樣性最高,秋季美人蕉的最低。從季節上看,冬季ACE、Chao1、Shannon指數分別為5892、6614.3、10.1;秋季各指數為4711.8、4637、8.68,冬季平均指數高于秋季,說明冬季土壤細菌的多樣性和豐富度高于秋季。從植物種類上看,冬季Shannon指數以苦草最大,為 10.33,其次為美人蕉和蘆葦,華克拉莎最低,為9.83;秋季則不同,華克拉莎、蘆葦指數較高,分別為10.04和10.17,美人蕉最低為5.13。總之,不同植物根際土壤細菌群落分布存在差異,根際細菌多樣性隨季節而變化,且群落變化規律有差異。
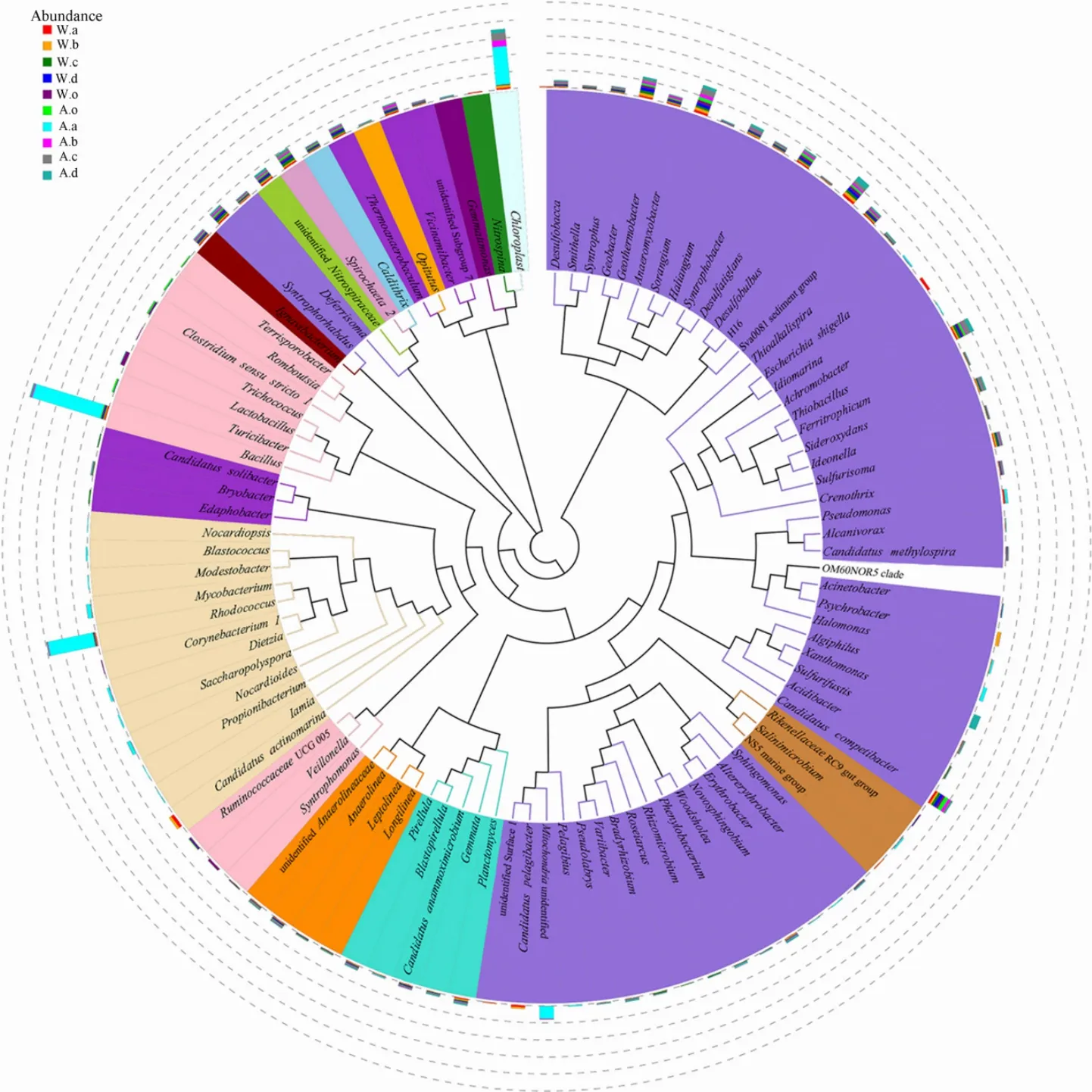
圖3 前100屬代表序列系統發育樹狀圖Fig. 3 Phylogenetic tree of top 100 genus represent sequence
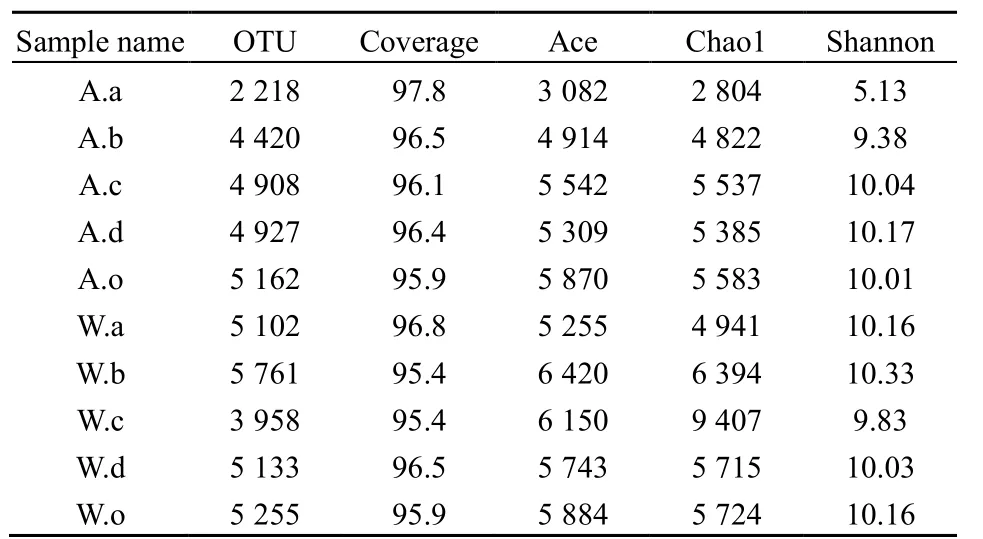
表2 秋、冬植物根際土壤中細菌的豐富度和多樣性指數Table 2 Diversity index of bacteria in plants rhizosphere soil of autumn and winter
為了解環境因子對細菌群落結構的影響,對環境因子和多樣性指數進行 Spearman相關分析(表3),結果表明,根際土壤中TP、TOC、TN與覆蓋率呈負相關,與Ace、Chao1、Shannon指數呈正相關,水質TP與多樣性指數關系則表現相反。運用典范對應分析(圖4)和Spearman分析(圖5),進一步篩選影響細菌群落的驅動因子,由圖可知,前兩軸共解釋了 95.25%的變異,第一主軸上的主要影響因子是土壤TOC、TN、TP,其相關系數分別是0.84、0.74、0.54,第二主軸上的影響因子是水質TP、TN、DO,其相關系數分別是0.71、0.61、0.66。其中水質 TP、TN、DO等對底泥中細菌群落有較大影響,土壤 TOC、TN、TN對植物根際土壤細菌群落有較大影響;而秋季美人蕉不同,主要受水質TP、TN、土壤pH的影響。土壤TOC、TN與變形菌門、迷蹤菌門豐度呈顯著正相關,與厚壁菌門呈顯著負相關,與藍藻菌門、放線菌門相關性不明顯;水質TP、TN、土壤pH與厚壁菌門呈極顯著正相關,與硝化螺旋菌門、酸桿菌門、浮游菌門呈顯著負相關,與綠彎菌門、擬桿菌門的相關性不大。
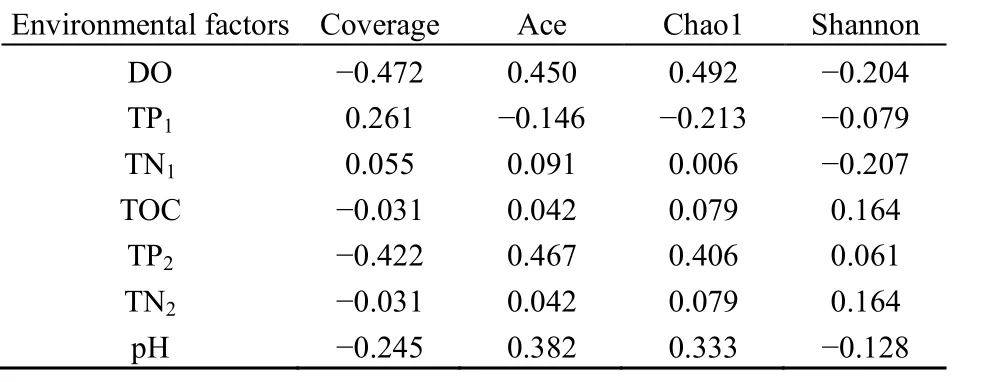
表3 環境因子與多樣性指數的Spearman相關分析Table3 Spearman correlation analysis of environmental factors and diversity index
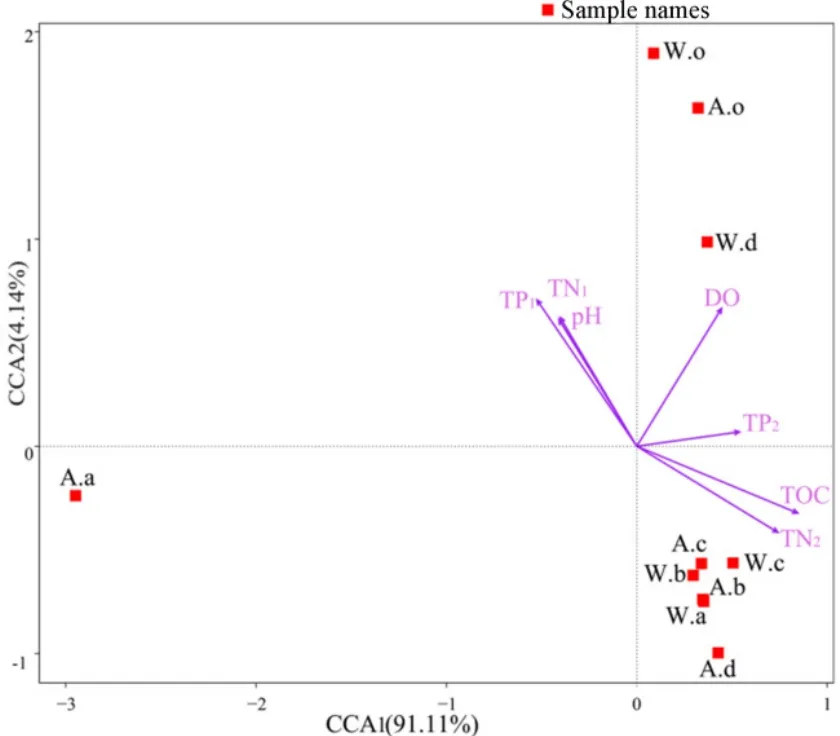
圖4 樣品與環境因子的CCA分析Fig. 4 CCA analysis between samples and environmental factors
2.4 細菌群落相似性和差異性分析
根據OTU聚類結果對樣品進行均一化處理后,繪制Venn圖(圖6),比較不同季節、不同植物細菌群落的相似性,10個樣品共有的OTU數為935,占總OTU數(46844)的2.0%。從季節上看,秋季共有OTU為1042(4.8%),而冬季為1182個(4.7%),這表明季節對植物根際土壤細菌的種類和數量有影響,但占主要比重的細菌種類幾乎無變化,還是以變形菌、綠彎菌為主。從種類上看,秋、冬季華克拉莎相似性最高,為35.5%,說明華克拉莎細菌群落季節變化最小;蘆葦次之,相似性為34.4%;然后是苦草為31.9%;美人蕉細菌群落季節變化最大,相似性最低為21.3%,對環境的響應更敏感。兩季底泥相似性最高(36.7%),說明底泥中細菌群落受溫度變化較小以及植物種類對細菌群落存在一定影響。聚類結果(圖 7)顯示,不同植物根際細菌群落之間有差異,分為4個分支,以同一個季節聚類在一起,說明溫度對植物根際土壤細菌群落的分布有一定影響。其中秋季美人蕉獨立成一個分支,主要原因是其土壤中營養物質含量明顯低于其他土樣,導致群落分布不同。
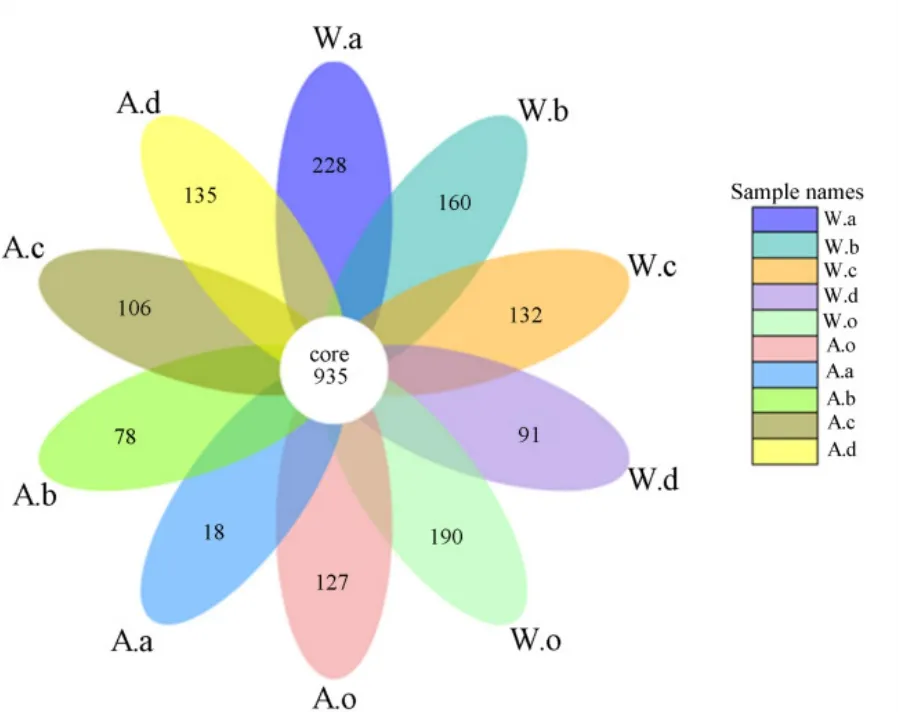
圖6 OTU venn分析Fig. 6 Analysis of OTU venn
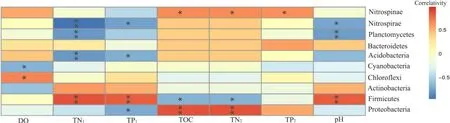
圖5 前10門細菌與環境因子相關分析Fig. 5 Correlation analysis between the top 10 bacteria and environmental factors
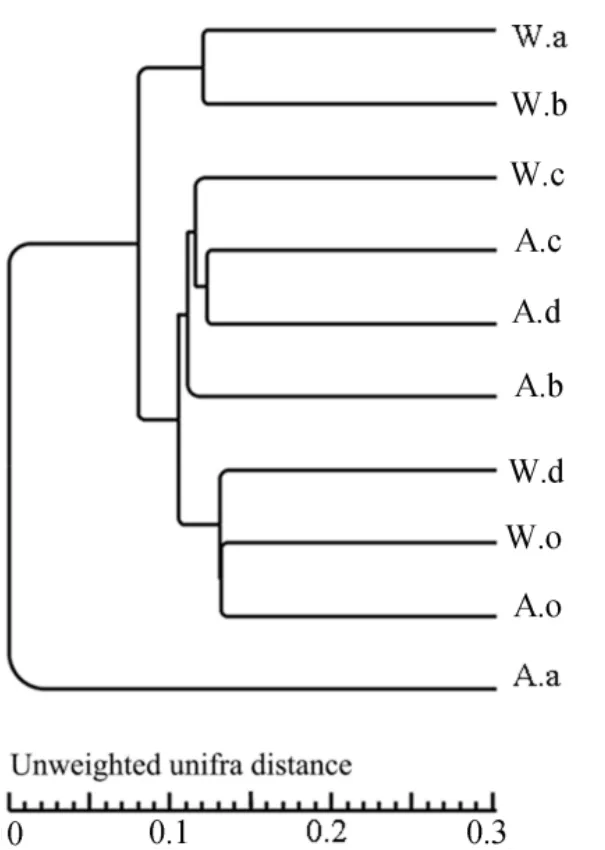
圖7 10門主要細菌聚類分析Fig. 7 Cluster analysis of 10 dominant bacteria
3 討論
3.1 細菌群落結構
本研究區植物根際土壤中細菌群落具有較高的多樣性,發現 10門類主要細菌:變形菌門、綠彎菌門、酸桿菌門、浮霉菌門、擬桿菌門、硝化螺旋菌門等,說明這些菌類分布范圍廣,并以變形菌門、綠彎菌門、酸桿菌門為主,三者約占總測序序列的57.0%-83.5%。而阮楚晉等(2018)對會仙濕地底泥可培養原核微生物多樣性的研究中,僅分離得到厚壁菌門、變形菌門、放線菌門、擬桿菌門這4門細菌,這說明采用現代分子學技術更能靈敏和準確地揭示細菌群落結構多樣性。
會仙濕地不同植物根際土壤中優勢菌均以變形菌、酸桿菌、綠彎菌為主,這一結果與國內外有關濕地土壤中微生物多樣性的研究結果一致(房昀昊等,2018)。劉興等(2015)對貴州喀斯特地區植被土壤群落結構進行研究,共發現 33門細菌,其優勢菌群與本文一致,但發現的細菌種類比會仙喀斯特濕地的低,說明會仙濕地中細菌種類豐富,對維持濕地系統的平衡有重要作用。研究發現,變形菌在會仙濕地秋、冬植物根際土壤中均為優勢菌,其中以 β-、δ-、γ-變形菌綱為優勢亞群,具有豐富的代謝多樣性,在 N、P、C等元素循環及其他污染物的降解過程中起關鍵作用(Hoefel et al.,2005;Ansola et al.,2014);通過分析發現秋季美人蕉中變形菌最低,水中N、P含量最高,而其他植物中變形菌較高,環境中N、P較低,同樣證明變形菌有較好的脫氮除磷作用。苦草中變形菌含量較高,表明苦草在脫氮除磷方面可能發揮重要作用。綠彎菌作為第二優勢菌,主要分布在有機物豐富的沉積物中,通常參與COD和萘的去除(Grégoire et al.,2011;楊小麗等,2013)。酸桿菌主要存在于濕地、活性淤泥中,能夠參與枯枝爛葉分解的碳循環(Chen et al.,2015),在蘆葦和華克拉莎中豐富度較高,說明蘆葦和華克拉莎的根際環境有利于碳的循環轉化。與其他植物相比,秋季美人蕉根際細菌群落結構明顯不同,以放線菌、厚壁菌和藍藻菌為主,而柔膜菌卻僅存在于其他植物根際土壤中,主要原因是土壤理化性質、植物群落類型對土壤細菌群落結構有較大影響(Zhao et al.,2010)。除此以外,秋季美人蕉根系土壤中酸桿菌、硝化螺旋菌明顯低于其他樣,可能原因是酸桿菌為嗜酸菌,而植物根際土壤pH值以秋季美人蕉最高(7.63),不利于其生長,同樣說明酸桿菌可以在非酸性條件下生存。硝化螺旋菌在好氧條件下可以將亞硝酸鹽氧化成硝酸鹽,在氮循環中起著重要作用,而美人蕉根際環境DO最低為4.13,抑制硝化螺旋菌的繁殖,導致硝化螺旋菌豐度最低;浮霉菌作為兼性厭氧菌和嚴格厭氧自養菌,在秋季美人蕉中最低,僅有0.9%,說明除DO外,還有其他因素影響植物根際土壤中細菌分布。
地桿菌屬、厭氧粘菌屬、硫桿菌屬、H16屬等在大多數植物根際土壤中皆存在,屬于革蘭氏陰性菌,可能是濕地植物根際環境有利于革蘭氏陰性菌生長,亦有利于環境中污染物的降解。研究表明(周文杰等,2015),假單胞菌屬(Pseudomonas)是根際有益微生物,對植物的生長有顯著的促進作用,但其對環境的適應能力較差,相比于其他根際微生物競爭較弱,營養物濃度是其主要影響因素。本研究得到了類似結果,相比于其他植物而言,美人蕉生長環境中的營養物濃度有明顯差異,從而導致其假單胞菌屬豐度最高。海源菌屬(Idiomarina)、嗜冷桿菌屬(Psychrobacter)只在冬季出現,可能原因是不同細菌對溫度等條件要求不同,有些細菌卻適宜在冬季環境下生存。韋榮球菌屬(Veillonella)只存在底泥中,說明植物根際對細菌群落分布有影響(Chaudhary et al.,2012)。此外,在植物根際環境中還發現了有益微生物(王建寧等,2012;林瑩瑩等,2016),如紅球菌屬和芽孢桿菌屬,它對碳源的利用率強,在促進植物生長和污染物降解中起重要作用,在秋季美人蕉中含量最高,可能是環境中碳源豐富促進其繁殖和秋季溫度更適應紅球菌的生長。
綜上所述,不同植物根際土壤中細菌群落結構有一定差別,但差別不大,占主導的細菌群落相同,相同植物根際土壤中細菌群落結構相似度較高(商迎迎,2017),營養物濃度對細菌分布有較大影響。
3.2 細菌群落結構多樣性和環境因子
秋、冬植物根際土壤細菌多樣性指數均較大,說明供微生物生長的營養物質較豐富(雷旭等,2015)。總體而言,冬季各植物根際多樣性高于秋季,主要是因為冬季土壤中營養物濃度高于秋季,除此以外,會仙濕地植物種類豐富、面積大,植物根際細菌對秋、冬季溫度變化具有較好的適應能力。綜合各指數可知,植物根際土壤細菌多樣性表現為蘆葦>華克拉莎>苦草>美人蕉,與趙慶節(2011)研究相似。本研究發現,蘆葦根際土壤細菌多樣性和豐富度最高,原因可能在低溫情況下,蘆葦根際的好氧菌仍保持較高比例以及蘆葦根系較其他植株更發達,有利于細菌的繁殖。值得注意的是,由于美人蕉是人工引入到濕地中的,在短時間內植物根際吸收作用不明顯,使得秋季美人蕉根際土壤中各指標(表 1)遠低于其他植物根際環境中的,故其細菌多樣性、豐富度遠低于其他樣。與植物相比,底泥中各多樣性指數最大,說明植物根際對某些細菌產生抑制作用,Zhou et al.(2009)研究表明,濕地表層中細菌的數量顯著高于其下層的數量,并且出現明顯的分層效應,形成好氧和厭氧層交替現象,種植不同植物對細菌群落分布的影響主要發生在0-10 cm處。
已有研究證明(Córdova-Kreylos et al.,2006;金笑等,2017),土壤TN、TOC是細菌群落結構影響的顯著因子,與微生物群落數量、生物量等呈顯著正相關。通過對環境因子與樣品的 CCA分析發現,影響植物根際土壤細菌群落結構的主要環境因子同樣是土壤 TOC、TN,其與變形菌門、迷蹤菌門呈顯著正相關,與厚壁菌門呈顯著負相關,而水質環境因子主要對底泥中細菌群落分布有影響。所有樣品分成3個相對獨立的群,其中秋季美人蕉獨立成一個群,其細菌群落結構與其他樣本有明顯差異,影響其細菌群落的主要環境因子也不同,可能原因是在土壤營養物較低的情況下,水中含量較高的營養物對其產生了主要影響。大多數不同植物形成一個群(圖4),對環境因子的響應趨勢相似,可能是不同植物對細菌群落結構有影響,但是沒有營養物濃度對細菌群落影響大。國內外相關研究表明(Buée et al.,2010),在土壤細菌群落研究中,pH作為指示因子之一,對細菌群落的分布有較大影響。本次研究同樣證明,pH對細菌群落有顯著影響,不同細菌對pH的響應不同,厚壁菌門豐度與pH呈極顯著正相關,硝化螺旋菌門、酸桿菌門、浮游菌門豐度與pH呈負相關,研究結果與劉興等(2015)相似。
3.3 細菌群落結構相似性和差異性
秋、冬植物根際土壤細菌群落相似性分析表明,不同植物根際細菌群落具有一定的相似性,但相似性較低,說明濕地土壤環境中形成了某些穩定的細菌群落,這些細菌適應濕地環境的變化,對底泥和植物根際環境中細菌群落結構有重要作用。秋季樣的整體相似性略高于冬季,說明冬季各植物細菌群落隨季節變化更大,主要原因可能是冬季植被生長、耐寒能力不同,各植物根際活動差異較大。美人蕉根際細菌群落結構在秋、冬兩季相似性最低,說明其受季節變化影響最大,其次是苦草和蘆葦,華克拉莎變化最小。原因可能是美人蕉相對于其他植物根際范圍小,沒有生長優勢,根際在秋、冬兩季活動差異相對更高,而蘆葦、華克拉莎在秋、冬兩季都為優勢植物,所以更為穩定。底泥細菌群落的相似性最高,為36.7%,說明濕地土壤細菌群落結構受到植物根際活動的影響,但并不能改變組成土壤細菌群落的主要種類,因為這是由溫度、植物種類、污水和土壤性質共同決定的。
4 結論
會仙濕地中植物根際土壤細菌有較高的多樣性,發現10門主要細菌,變形菌為第一優勢菌群,所占比例為42.4%;優勢菌綱占比最高的菌群為β-變形菌(13.5%);優勢菌屬占比最高的菌群為芽孢桿菌屬(3.8%)。其中,秋季美人蕉根際中細菌種類與其他樣本明顯不同,主要是由于其生長土壤環境中營養物濃度低導致。從季節上看,秋、冬植物根際土壤細菌多樣性和豐富度具有差異,冬季高于秋季;從植物種類上看,蘆葦多樣性和豐富度最高,其次為華克拉莎和苦草,美人蕉最低。相似性分析表明,同一植物不同季節其細菌群落結構相似性較高,美人蕉細菌群落結構隨季節變化最大,其次為苦草和蘆葦,華克拉莎變化最小。植物根際土壤細菌群落結構和多樣性受環境因子的影響較大,其中土壤TOC、TN對細菌群落影響最為顯著。從植物根際土壤微生物多樣性、豐富度、優勢菌種來看,在會仙濕地中增加優勢植物蘆葦和華克拉莎的種植,維持濕地系統中微生物多樣性,利于濕地的修復和保護。
猜你喜歡
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中華詩詞(2020年1期)2020-09-21 09:24:52
中國生殖健康(2020年6期)2020-02-01 06:28:50
中國生殖健康(2019年11期)2019-01-07 01:28:02
小學生作文(中高年級適用)(2018年5期)2018-06-11 01:22:56
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
中學生數理化·七年級數學人教版(2017年11期)2017-04-23 07:18:00
爆笑show(2016年7期)2017-02-09 09:36:13
數學大王·中高年級(2016年12期)2016-12-26 21:37:36
