CRISPR/Cas9基因編輯技術在真菌中的應用
2019-04-04 01:06:36蘇文英楊和川譚一羅付永平
江西農業學報 2019年3期
蘇文英,楊和川,譚一羅,李 曉,付永平*,李 玉*
(1.連云港市農業科學院,江蘇 連云港 222006;2.吉林農業大學 食藥用菌教育部工程中心,吉林 長春 130118)
隨著分子生物學技術的快速發展,基因編輯技術已經成為基因組改造和基因功能研究的熱點。早期僅基于同源重組的基因打靶技術效率比較低,限制了基因打靶技術的應用。后來,研究者們用改造過的人工核酸內切酶(Engineered Endonuclease,EEN)在基因組的特定位置使DNA雙鏈斷裂,并在修復過程中達到基因定點編輯的作用,人工核酸內切酶的出現,徹底改變了基因編輯效率低的這一現狀[1]。早期的EEN介導的基因編輯技術主要包括鋅指核酸內切酶技術(Zinc-finger Nucleases,ZFN)、類轉錄激活因子效應物核酸酶技術(Transcription Activator-like Effector Nucleases,TALEN),這2種技術的作用原理都是通過DNA結合蛋白與核酸內切酶Fok Ⅰ組成蛋白復合物對靶點進行特異性的識別和切割[2]。然而,這2項技術在操作過程中不僅費時費力,而且對操作技術要求很高,因而在普通實驗室很難被有效利用。2013年初,新型基因編輯技術CRISPR/Cas系統進入人們的視野[3],因其制作成本低廉、操作過程簡單、快捷且易于掌握的特點,迅速在作物品種改良[4-8]、動物模型構建[9-10]、疾病治療研究[11-13]、發酵菌株代謝途徑改造[14-15]等領域得到了廣泛的應用。但在真菌中,由于部分真菌同源重組率極低、高效遺傳轉化體系的缺乏、啟動子和有效篩選標記的缺乏等原因,阻礙了該技術在真菌中的發展。本文綜述了近幾年CRISPR/Cas9技術在真菌中的開發和應用,以期為進一步加快真菌基因編輯研究提供參考。
1 CRISPR/Cas9基因編輯技術及其作用原理
1.1 CRISPR/Cas9基因編輯技術概述
1987年,日本學者首次在大腸桿菌(Escherichiacoli)堿性磷酸酶基因(Alkaline Phosphatase,IAP)的側翼序列中發現了成簇的有規律間隔的短回文重復序列[17];Jansen等[18]將其正式命名為成簇規律間隔短回文重復序列(Clustered Regularly Interspaced Short Palindromic Repeats,CRISPR)。之后,研究者發現在40%的細菌和90%的古細菌的基因組中都存在CRISPR位點[19]。Barrangou等[20]對嗜熱鏈球菌(Streptococcusthermophilus)進行遺傳改造時,發現嗜熱鏈球菌的CRISPR/Cas Ⅱ型系統能夠抵抗外源噬菌體的入侵。隨著研究的進一步深入,Marraffini等[21]發現細菌中的CRISPR系統能抵御外源質粒的轉移,并通過實驗驗證了CRISPR系統的功能。Feng Zhang團隊的Cong L等[22]和George M. Church團隊的Mali P等[23]同時在《Science》雜志上在線報道了應用sgRNA介導的Cas9核酸酶對哺乳動物細胞基因組的特異性打靶。此后,利用該技術在不同生物中進行基因編輯的工作陸續展開。
1.2 CRISPR/Cas9基因編輯技術的作用原理
由于Cas基因的多樣性異常豐富,研究者根據Cas基因核心元件序列的不同,將CRISPR/Cas免疫系統分為Type Ⅰ、Type Ⅱ、Type Ⅲ共3種不同類型。TypeⅠ系統主要存在于細菌和古生菌中,結構最為復雜,特征蛋白為Cas3 蛋白,該蛋白具有解旋酶和核酸酶功能[24];Type Ⅲ系統的特征蛋白為Cas10 蛋白,該蛋白具有RNA 酶活性[25]。而在這3種不同類型中,由于Type Ⅱ型系統的結構簡單,除去crRNA和tracrRNA外,只有一個標志性Cas9蛋白來切割DNA雙鏈,并且它僅存在于細菌中[26]。因此到目前為止,被改造的最為成功的人工核酸內切酶是產膿鏈球菌(Streptococcuspyogenes,SF370)的Type Ⅱ型系統,并且已經在多種生物中成功實現了基因組定點修飾。
CRISPR/Cas的組織結構相對簡單。以產膿鏈球菌SF370的Type Ⅱ型結構為例,其結構可被分為3個部分:第一部分為5′端的轉錄激活RNA(tracrRNA)基因;第二部分為中間一系列與CRISPR相關的Cas(CRISPR-associated)蛋白,目前發現有6個核心Cas蛋白,包括Cas9、Cas1、Cas2、Csn2,其中Cas9蛋白包含2個獨特的功能結構域HNH和類RuvC核酸酶亞基,它們分別切割DNA雙鏈,從而產生基因定點突變;第三部分為3′端的CRISPR基因座,由前導區(Leader)、間隔區(Spacers)以及多個重復序列(Direct repeats)順序連接而成[27]。
CRISPR/Cas9的作用機理大體可以分為3個步驟:(1)外源質粒上與CRISPR基因座上對應的序列被稱為protospacer,它的5′端或3′端的3~5個保護堿基,被稱為PAM(Protospacer Adjacent motif)序列。當外源質粒入侵時,通過識別PAM序列來將其附近的序列定為protospacer,然后該序列被整合到CRISPR基因座的兩個重復序列間,即獲得間隔序列;(2)當外源質粒再次入侵時,CRISPR基因座與插入的新間隔序列首先被轉錄成長的前體RNA(pre-crRNA),然后在Cas蛋白和RNase Ⅲ核酸酶的作用下被加工成含有間隔序列的crRNA;(3)然后,crRNA與特異的Cas蛋白以及轉錄激活RNA tracrRNA形成復合物crRNA- tracrRNA,再通過PAM序列掃描外源入侵DNA上的靶序列,crRNA上的間隔序列與靶序列進行互補配對,最后Cas9蛋白發揮切割酶活性,外源噬菌體或是質粒DNA在配對的特定位置被切割,從而達到防御的目的[28]。具體的作用機理如圖1所示。
2 CRISPR/Cas9基因編輯技術在真菌中的應用
2.1 在酵母菌中的應用
2.1.1 釀酒酵母 早在2013年,DiCarlo等[30]就將CRISPR/Cas9技術應用到了釀酒酵母(Saccharomycescerevisiae)中,研究者將攜帶 sgRNA的質粒與供體DNA同時轉至含有組成型表達的Cas9細胞中,使供體DNA的同源重組率接近100%,而且相較于傳統的同詢重組技術,該技術使單鏈及雙鏈靶基因斷裂效率分別提高5倍和130倍,該研究為后續酵母基因定點突變和等位基因替換研究提供了強有力的支撐。隨后,Bao等[31]構建了一步實現多基因敲除的CRISPR-Cas系統——HI-CRISPR(Homology-Integrated CRISPR-Cas),在對釀酒酵母進行基因編輯時,其中3種基因CAN1、ADE2、LYP1在4 d內同時被破壞,效率為27%~87%;另外3個基因ATF2、GCY1、YPR1在6 d內同時被破壞,效率為100%,該研究實現了在釀酒酵母中進行多基因編輯,大大提高了基因編輯效率。
2.1.2 裂殖酵母 裂殖酵母(Schizosaccharomycespombe)與真核生物基因組的相似性高于釀酒酵母,但由于分子工具的缺乏,關于裂殖酵母的研究不如釀酒酵母深入。特別是由于缺乏用于表達sgRNA的啟動子而阻礙了CRISPR/Cas9基因組編輯體系在裂殖酵母中的實施。2014 年,Jacobs 等[32]開發了應用于裂殖酵母中的CRISPR/Cas9系統,在該研究中研究者利用RNA(rrk1)的啟動子構建靶向指導RNA的表達載體,與組成型Cas9相結合,在裂殖酵母中實現無篩選標記的特異性誘變,誘變效率接近100%,實現了在裂殖酵母中進行特定、精確和高效的基因組編輯。
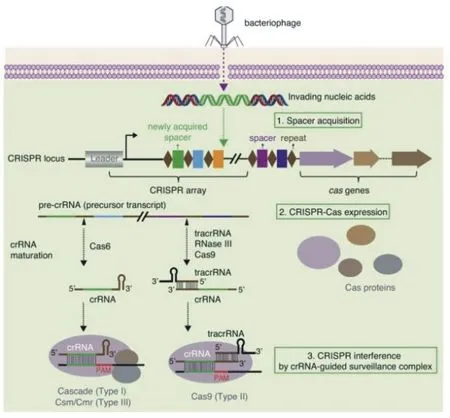
圖1 CRISPR/Cas系統的作用機理[29]
2.1.3 白色假絲酵母 Shapiro等[33]建立了一種基于CRISPR/Cas9的基因驅動平臺(Gene Drive Array,GDA),能夠在二倍體病原體中進行基因操作及快速形成突變體,該研究被用于對白色假絲酵母(Candidaalbicans)高效基因編輯中,在此研究中,一種DNA切割的Cas9酶靶向2個區域,位于單倍體白色念珠菌真菌中一個基因的兩側,由兩個所謂的“向導 RNA”(gRNAs) 基因組成。在靶向基因序列被切斷后,一種基因驅動的盒式表達,將所有的 Cas9 和 gRNA 組件插入其位置。當2個單倍體菌類交配形成二倍體后代時,基因驅動也將替代基因在另一個染色體上的對應基因,得到目標基因有效地從機體完全刪除的原始版本。這項研究為了解白色念珠菌致病機制和耐藥性提供了新的途徑。
2.2 在霉菌中的應用
2.2.1 里氏木霉 2015年,Liu等[34]通過特定的密碼子優化和體外RNA轉錄,融合增強的綠色熒光蛋白(eGFP)對其進行檢測,建立了適用于絲狀真菌里氏木霉(Trichodermareesei)的CRISPR/Cas9系統,不僅可以在同源臂較短的條件下實現靶基因高效率的同源重組,也可對多個靶基因進行同時編輯。CRISPR/Cas9系統在里氏木霉中的成功應用,為進一步加速絲狀真菌的功能基因組學和菌株改良研究奠定了基礎。
2.2.2 米曲霉 Katayama等[35]利用CRISPR/Cas9系統對工業菌株米曲霉(Aspergillusoryzae)基因組進行了編輯,在該研究中研究者利用amyB作為Cas9的啟動子,來自米曲霉的U6 啟動子作為gRNA 啟動子,SV40作為核定位信號,FLAG-tag作為蛋白表達與否的檢驗標記,構建了靶向于wA、yA、pyrG基因的載體,突變率在10%~20%之間,結果表明:單堿基的缺失或插入突變較多,且不會對米曲霉的生長造成影響。該技術在米曲霉中的應用有利于后續米曲霉定向誘變研究。
2.2.3 煙曲霉 Zhang等[36]在煙曲霉(Aspergillusfumigatus)中建立了以 CRISPR/Cas9 技術為基礎的微生物學介導的末端連接(Microhomology-Mediated End Joining,MMEJ)靶基因突變系統,不僅實現了精確高效的靶基因編輯,并且通過35 bp的同源臂就可使基因的編輯效率高達95%~100%。
2.2.4 水稻稻瘟病菌 在水稻稻瘟病菌(Pyriculariaoryzae)中,Arazoe等[37]分別利用內源性的 RNA 聚合酶Ⅲ(RNA polymerase, RNAP)所識別的U6啟動子和 RNA 聚合酶Ⅱ控制的TrpC啟動子表達 sgRNA,同時發現利用RNA聚合酶Ⅲ識別的U6啟動子比利用 RNA 聚合酶Ⅱ識別的TrpC啟動子進行基因組編輯的效率更高,這表明驅動sgRNA的啟動子也會影響敲除效率。
2.2.5 玉米黑粉菌 在玉米黑粉菌(Ustilagomaydis)中,Schuster等[38]使用密碼子優化的Cas9蛋白、組成型強啟動子Potef、gRNA以及來源于自身的U6 啟動子構建CRISPR/Cas9基因編輯體系,通過單步轉化法導入,對基因bE2和bW1進行編輯,結果表明:突變效率高達70%,為后續玉米黑粉菌致病基因功能的研究提供了很大的幫助。
2.3 在蕈菌中的應用
2.3.1 雙孢蘑菇 2016年賓夕法尼亞大學的楊亦農教授利用該技術對雙孢蘑菇(Agaricusbisporus)中控制褐變的多酚氧化酶(PPO)的基因進行了基因編輯,并將該酶的活性降低了30%,使其抗褐變。此研究雖然沒有公開具體操作過程,但這是基因編輯技術在大型真菌中的首次應用[39]。
2.3.2 灰蓋鬼傘 2017年,Sugano 等[40]構建了應用于大型真菌模式生物灰蓋鬼傘(Coprinopsiscinerea)中的高效的基因組編輯體系,在此研究中,作者利用高通量轉化方法篩選獲得的一個活性高出常規啟動子GPD活性7倍的新啟動子——CcDED1pro,然后將該啟動子作用于Cas9蛋白,來自灰蓋鬼傘的U6-snRNA啟動子表達gRNA,構建載體,利用PEG介導法轉化冷凍保存的原生質體。最后,利用上述體系在穩定的 GFP 表達體系中進行 CRISPR/Cas9 介導的 GFP 誘變,并在菌絲和子實體中成功檢測到了GFP 功能的喪失。該研究加速了大型真菌遺傳及分子育種研究。
2.3.3 金針菇 劉建雨等[41]將SpCas9根據金針菇(Flammulinavelutipes)密碼子偏好性進行了密碼子優化并合成FvCas9全長序列,構建了FvCas9雙元表達載體,通過農桿菌介導法轉化金針菇單核體Dan3,并成功獲得了轉化子。
2.3.4 靈芝 Qin等[42]在靈芝(Ganodermalucidum)中成功應用了CRISPR/Cas9系統,在此研究中,作者應用T7啟動子表達gRNAs,來源于靈芝自身的Pgpd啟動子和來自里氏木霉的Tpdc終止子作用于Cas9,然后利用PEG介導法轉化靈芝原生質體,靶向破壞靈芝中阻礙靈芝酸合成的基因——URA3,并成功獲得了轉化子,轉化效率高達66.6%,從而使靈芝產生更多具有抗腫瘤活性和抗轉移活性的靈芝酸。該研究為未來更高級真菌的深入研究和應用提供了有效的平臺。
2.3.5 蛹蟲草 孫丹[43]將植物敲除載體pFGC-pco Cas9 通過PmeⅠ和NcoⅠ兩種限制性內切酶將用于真菌的GPD啟動子成功地構建到pFGC-pco Cas9中,從而成功改造了適用于真菌敲除的載體,利用農桿菌介導法轉化蛹蟲草(Cordycepsmilitaris)原生質體,得到關于URA3基因(gRNA1+gRNA2)的轉化子共 70+52個,其中gRNA1的突變率為2.8%,gRNA2突變率為2.0%。隨后,Chen等[44]通過密碼子偏好性對Cas9酶進行優化,并將其與新報道的啟動子Pcmlsm3和終止子Tcmura3一起表達,構建了應用于蛹蟲草的CRISPR/Cas9系統,并通過熒光GFP標簽和蛋白質印跡分析證明該系統可以穩定表達。利用農桿菌介導法將體外合成的sgRNA和供體單鏈DNA(ssDNA)轉化蛹蟲草原生質體,利用5-FOA作為篩選標記,并成功獲得轉化子。該研究為進一步提高蟲草素的產量,促進相關產業的發展提供了技術支持。
隨著被成功測序的蕈菌數目的增加及實驗技術的進步,尋找子實體發育的關鍵基因,并探究不同基因的具體功能和參與的調控機制也變得越來越重要。目前CRISPR/Cas9 系統在蕈菌中的應用還比較少,且多集中在CRISPR/Cas9基因編輯體系的建立,并且突變體的篩選多依靠測序完成,工作量很大。相信隨著CRISPR/Cas9技術的不斷改進和完善,科學家們能夠建立一種高效簡便的CRISPR/Cas9基因編輯體系,為蕈菌功能基因挖掘、遺傳育種研究奠定基礎。
3 影響CRISPR/Cas9系統編輯效率的因素
首先,該系統最大的問題就是脫靶率較高,CRISPR/Cas9 基因編輯系統的特異性取決于 sgRNA 上的識別序列,因此如何設計高效、特異性的sgRNA是降低脫靶效率的關鍵。研究表明,在設計sgRNA 時,高C/G含量百分比可以有效降低脫靶率[45]。在設計 gRNA 時,可通過特定算法預測潛在脫靶位點,盡可能選擇脫靶效應低的 gRNA,現在已使用各種算法的專業軟件來降低脫靶效率,如Cas-OFFinder、CRISPR Design Tool、CasFinder、CHOPCHOP、CRISPOR、E-CRISPR等軟件。此外,研究發現降低sgRNA的濃度也有助于降低脫靶率[46]。另一項研究也表明,較長的 PAM 序列能夠有效地減少脫靶效應[47]。
其次,同源臂的長短、質粒的濃度、啟動子及篩選標記的選擇以及是否建立了高效的遺傳轉化體系等因素都會影響CRISPR/Cas9系統的編輯效率。Paix等[48]研究發現由33~38個核苷酸組成的同源臂與由518個核苷酸組成的同源臂成功率相當,最優編輯條件下的成功率為10%~20%。Zhang等[36]在煙曲霉(Aspergillusfumigatus)中的研究也表明:通過35 bp的同源臂就可使基因的編輯效率高達95%~100%,并且使用gpdA或niiA等強啟動子驅動 Cas9的表達能有效提高敲除效率。Arazoe等[37]研究表明:采用U6啟動子表達 sgRNA進行基因編輯的效率高于tripC啟動子。這些研究都進一步表明不同來源的啟動子的靶標范圍、精準度和表達效率也不相同。
Matsuura等[49]發現,在一定的范圍內,Cas9和gRNA載體的濃度越高,轉化效率越高。Schuster 等[38]認為在制備原生質體時,如果采用混合的破壁酶將會導致脫靶率較高。此外,CRISPR/Cas9技術主要是通過基因槍和農桿菌介導的受體遺傳轉化法來實現將CRISPR/Cas9系統帶入到受體體內。所以,穩定高效的遺傳轉化體系也是CRISPR/Cas9成功高效運用的因素之一。因此,在今后的應用過程中,仍然要對這些影響基因編輯效率的因素進行進一步優化,加速推動該技術在真菌中的應用。
4 結語
綜上所述,利用 CRISPR/Cas9 介導的基因編輯技術可以有效地對真菌的靶基因進行編輯,利用該體系不僅可以對單個的目的基因進行編輯,也可對基因家族、整個基因座及代謝通路中的多個基因同時進行編輯。并且與ZFN和TALEN這兩種傳統的人工核酸酶相比,CRISPR/Cas9系統構建簡單、方便、快捷,且對細胞毒性小。但是運用過程中出現的脫靶、編輯效率低以及突變體篩選工作量大等問題也亟待解決。相信在不久的將來,隨著CRISPR/Cas9技術的改進和完善,真菌基因組的編輯將會變得越來越簡單,為進一步推進真菌功能基因組學及遺傳育種研究帶來突破性進展。
猜你喜歡
工業設計(2022年8期)2022-09-09 07:43:20
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
軍民兩用技術與產品(2021年10期)2021-03-16 06:05:30
北京測繪(2020年12期)2020-12-29 01:33:58
甘肅教育(2020年14期)2020-09-11 07:57:42
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
家庭影院技術(2017年9期)2017-09-26 03:41:45
時代英語·高二(2015年1期)2015-03-16 00:08:11
