細胞分裂素和赤霉素對3種山茶科種子萌發特性的影響
2019-03-21 07:18:26,,,
種子 2019年2期
,,,
(廣西壯族自治區中國科學院廣西植物研究所, 廣西 桂林 541006)
山茶科(Theaceae)植物種類繁多,全科約36屬700種,廣泛分布于熱帶和亞熱帶,主要集中于亞洲,其中我國分布有15屬480余種[1]。山茶科植物具有重要的經濟價值,如茶組中多種茶的嫩葉可加工成茶葉,是風靡世界的飲品;石筆木屬和金花茶組內多種植物花色艷麗、樹形優美,觀賞價值極高;油茶組和紅山茶組內60多種植物茶籽含有豐富的油脂,是廣泛種植的重要油料植物,具有較高的食用和藥用價值[1-8]。此外,山茶科植物還包含多種具有經濟用途和觀賞價值的種類尚未被開發利用[9-12]。由于環境變化和過度采集,山茶科內大部分野外資源日益減少,已有不少種類被列為稀有瀕危種[13-15]。山茶科種子具有堅實質密的種皮,透水和透氣性差,屬于頑拗性種子且具有休眠特性,野外種子萌發率和成苗率低[16-17]。因此,打破山茶科種子休眠,提高發芽率和成苗率是山茶科植物擴大開發利用的重要手段之一。
博白大果油茶(Camelliagigantocarpa)、多齒紅山茶(Camelliapolyodonta)、石筆木(Tutcheriachampioni)均為山茶科常綠喬木樹種,花碩大,樹形優美,茶籽油脂含量高,是極具價值的園林樹種和油料植物[18-23]。目前,3種山茶科植物只有零星栽培或處于野生狀態,產業化程度不高,其繁殖手段主要采用扦插方式[20-22,25],而相關種子育苗技術及外源激素對種子萌發影響的研究報道偏少。因此,本研究采用不同濃度的赤霉素(GA3)、細胞分裂素(6-BA)對博白大果油茶、多齒紅山茶、石筆木的種子進行處理,研究其對3種山茶科種子發芽的影響,以期為外源激素用于山茶科種子育苗提供實驗依據。
1 材料與方法
1.1 材 料
3種山茶科種子采集于桂林植物園內,采收時間為2017年10月中下旬,果實采收后置于陰涼處,待果皮開裂后,取出種子,并將種子冷藏于4 ℃冰箱。
1.2 實驗方法
1.2.1 種子重量及形態觀察
試驗在廣西植物研究所園林園藝研究中心進行。對采集后的3種山茶科種子形狀、大小、顏色等形態特征進行觀察。使用精密電子天平、游標卡尺分別測量50顆種子形態特征:長、寬、厚,測百粒重及單粒種子重量,百粒重重復測量10次,計算各指標的變異系數。
1.2.2 種子萌發抑制試驗
參照陳香波等[26]的方法,將3種山茶科種子材料的種皮和胚分離,并稱取博白大果油茶、多齒紅山茶的外種皮、內種皮和胚各0.5 g;由于石筆木的內外種皮難分離,因此,稱取石筆木的種皮和胚各0.5 g,分別用研缽研磨,研磨后置于錐形瓶中,加入100 mL 80%甲醇,放入4 ℃冰箱,浸提60 h,浸提過程中振蕩數次,過濾后得到種子不同物質的醇提取物。將3種山茶科種子不同部位醇提取物為原液稀釋到5%,同時以80%甲醇為原液稀釋到5%,作為甲醇對照。用5%的醇提取物和甲醇對照浸泡白菜種子3 h,設置3個重復,每個重復100粒種子,另外設置無菌水處理組。將浸泡后的種子放入2層濾紙的培養皿中,并加入適量對應的浸泡液潤濕濾紙,于25 ℃種子培養箱避光培養,48 h后測定發芽率。
1.2.3 種子破休眠處理
選擇無病蟲害的3種山茶科種子各30粒,用0.5%的高錳酸鉀浸泡15 min后,蒸餾水沖洗干凈。在各激素浸泡24 h,其中6-BA和GA3分別設計400,200,100 mg/L不同濃度梯度。對照組(Control group)為蒸餾水浸泡24 h,3種山茶科種子和激素濃度梯度各設置3個重復。將處理完成后的種子放入鋪有2層發芽紙的萌發盒內,種子萌發期間保持濕潤,種子培養箱內進行種子萌發實驗,培養箱溫度設置為25 ℃,濕度為90%,黑暗培養。每3天觀察1次種子發芽情況,以胚根伸長2 mm為種子發芽標準,持續觀測60 d。 測試指標如下:
發芽啟動時間為從發芽試驗開始到第1粒種子開始萌發所需天數(d)
發芽率(%)=(種子發芽數/試驗種子總數)×100%;
相對發芽率(%)=(處理組發芽數/對照組發芽數)×100%;
發芽勢(%)=(規定時間內發芽種子總數/供試種子總數)×100%;
發芽指數=∑在不同時間的發芽數/相應的發芽天數。
1.2.4 數據統計與分析
采用SPSS 16.0軟件對數據進行統計分析。差異顯著性分析應用Duncan法進行多重比較檢驗,檢驗顯著性水平為0.05,計算數據采用平均值±標準誤表達。
2 結果與分析
2.1 3種山茶科種子形態特征和種子重量
圖1所示,博白大果油茶和多齒紅山茶種子外種皮均呈茶褐色,石筆木種子種皮呈現黑褐色或茶褐色。3種山茶科種子種皮角質堅硬,且光滑無毛。測定結果發現(見表1):3種山茶科種子體積大小依次為:博白大果油茶>多齒紅山茶>石筆木。變異指數顯示,3種山茶科種子長度平均變異系數值相對較小,而種子寬度和厚度平均變異系數相對較大。此外,不同山茶科種子重量有顯著差異,博白大果油茶種子顆粒最大,質量最大,為3.779 g/顆,多齒紅山茶次之,最低為石筆木種子,平均為1.899 g/顆。

注:A為博白大果油茶;B為多齒紅山茶;C為石筆木。 圖1 3種山茶科種子形態特征
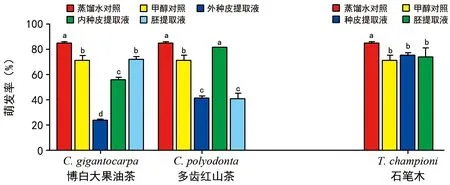
圖2 3種山茶科種子各部位浸提液對白菜種子萌發的抑制作用
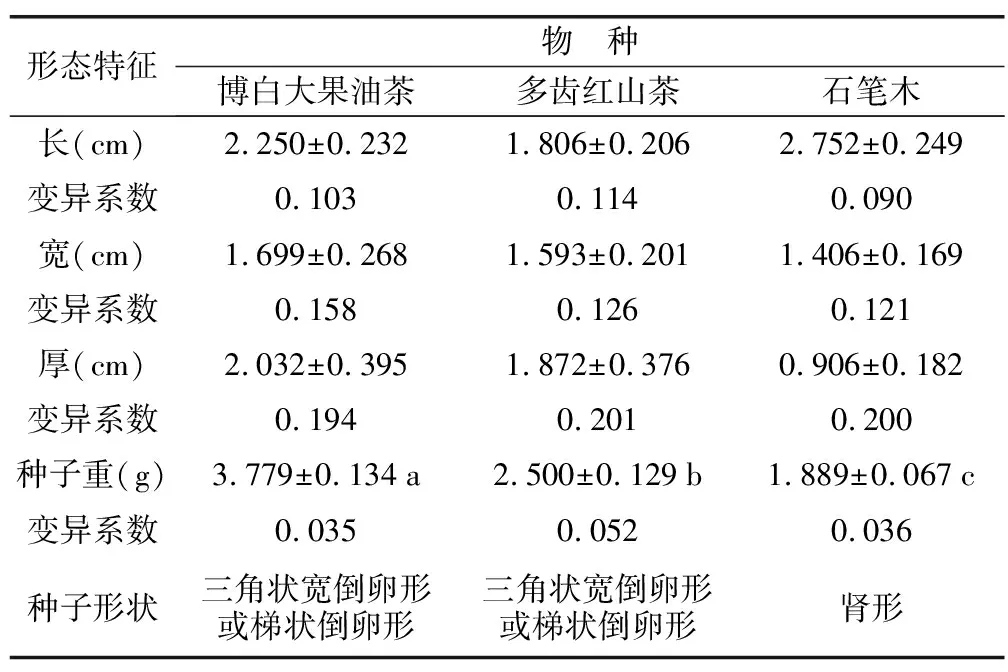
表1 3種山茶科種子形態特征
注:表中同行不同小寫字母表示在0.05水平的差異顯著性。下同。
2.2 種子萌發抑制作用
3種山茶科種子各部分提取液對白菜種子萌發的抑制結果見圖2,其中博白大果油茶種子的萌發抑制物質主要存在于內外種皮中,與甲醇對照間存在顯著差異,且外種皮提取液對白菜種子萌發抑制強于內種皮提取液,具有顯著性差異。多齒紅山茶的萌發抑制物質主要存在于外種皮和胚中,與甲醇對照間存在顯著差異,而內種皮提取液處理白菜種子的萌發率顯著高于外種皮提取液,且具有顯著性差異。石筆木種子的5%種皮和胚提取液對白菜種子萌發無明顯抑制作用,與甲醇對照組間無顯著差異。
2.3 不同激素濃度處理對3種山茶科種子萌發的影響
2.3.1 不同激素濃度對博白大果油茶種子萌發的影響
表2所示,博白大果油茶種子經激素浸泡24 h后的博白大果茶種子在6 d時可以啟動發芽,而對照組在12 d時才啟動發芽,表明6-BA和GA3對博白大果油茶種子萌動有一定的促進作用。在48 d時,博白大果油茶種子萌發基本結束,其統計結果見表2,其中濃度為6-BA 100 mg/L和GA3200 mg/L浸泡的博白大果油茶種子萌發率最高,分別達到99.07%和99.05%,不同激素濃度浸泡24 h的種子發芽率均高于對照組發芽率。表明6-BA和GA3對博白大果油茶發芽有促進作用,但與對照相比無顯著性差異。在發芽勢方面,除了GA3100 mg/L處理組發芽勢低于對照組外,其余處理組發芽勢均高于對照組,其中6-BA 100 mg/L處理下的發芽勢最高,達到87.69%,顯著高于對照組,表明博白大果油茶種子在激素處理后,促進種子生命力,萌發速度加快。此外,激素處理后的種子發芽指數顯著高于對照,且具有顯著性差異,表明激素處理后的博白大果油茶種子發芽速度和整齊度均高于對照組。
2.3.2 不同激素濃度對多齒紅山茶種子萌發的影響
通過分析發現,在第12天時,不同濃度GA3處理的多齒紅山茶種子均出現萌動,而不同濃度6-BA處理組和對照組均在15 d時啟動萌發,GA3處理的多齒紅山茶萌發結束時間為第42天,6-BA處理組和對照組萌發結束時間為第54天,表明GA3處理多齒紅山茶種子24 h后,能促進其提早萌發,縮短種子萌發時間。表3結果顯示,GA3處理多齒紅山茶種子的發芽率、相對發芽率、發芽勢、發芽指數均顯著高于6-BA處理和對照組,且具有顯著性差異,其發芽率達95.9%~98.8%,而6-BA處理的發芽率為72.8%~85.5%,對照組發芽率為86.37%,表明GA3處理能提高多齒紅山茶種子發芽率和種子生命力。
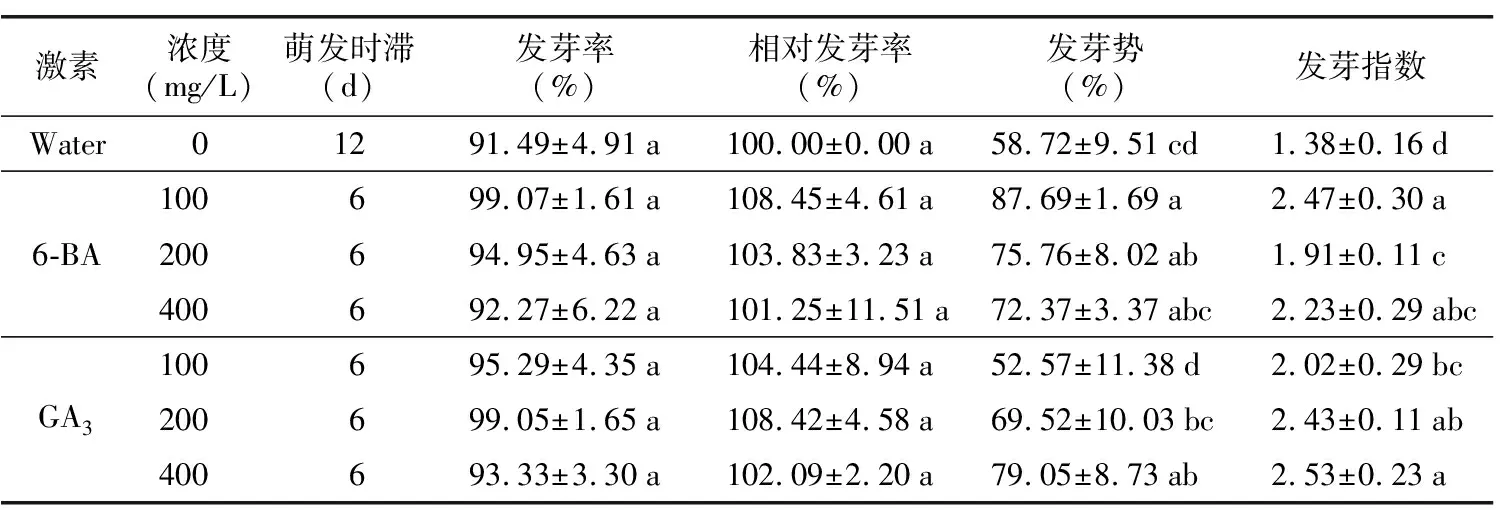
表2 不同激素濃度處理下博白大果油茶發芽情況
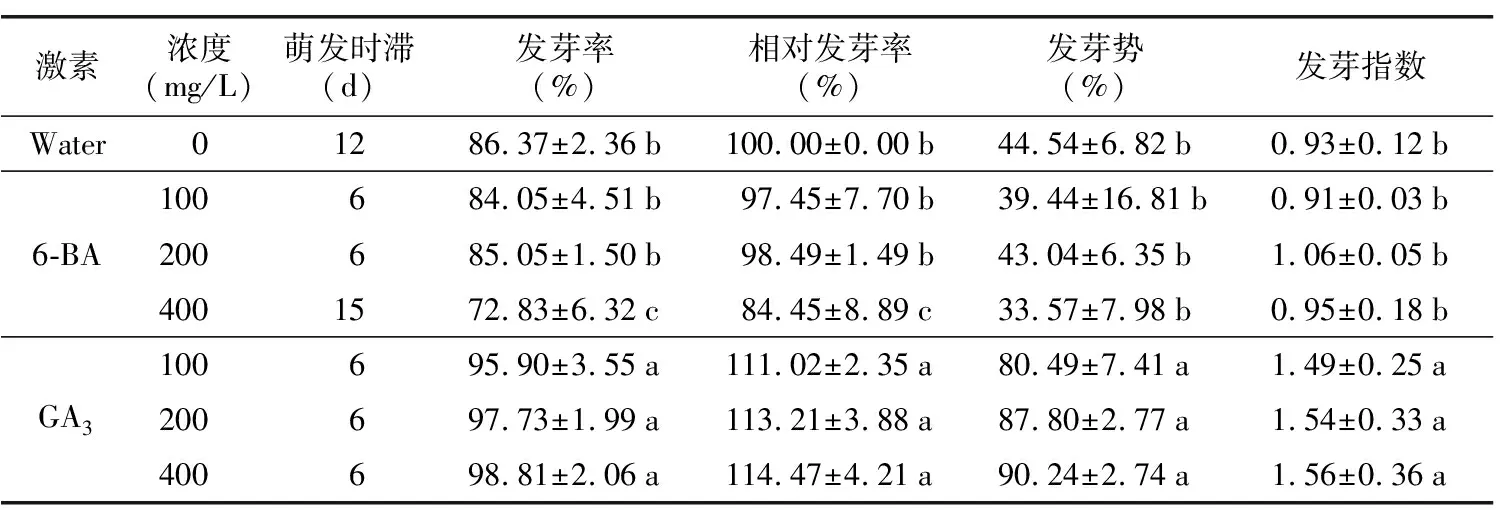
表3 不同激素濃度處理下多齒紅山茶發芽情況
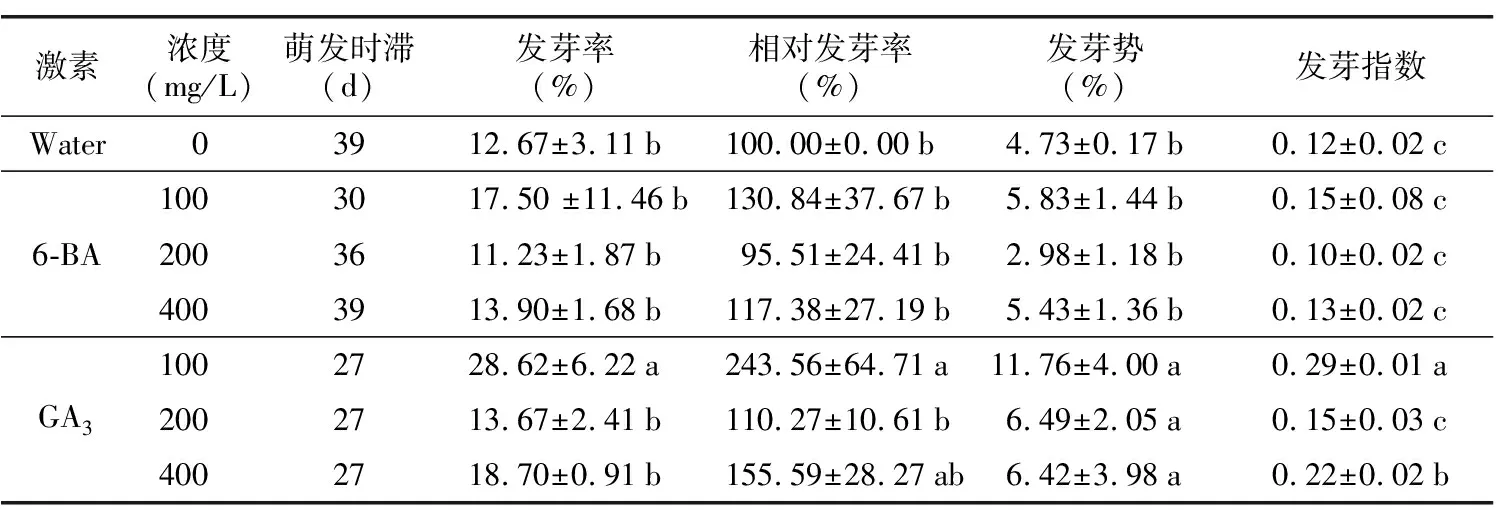
表4 不同激素濃度處理下石筆木發芽情況
2.3.3 不同激素濃度對石筆木種子萌發的影響
通過分析可知,石筆木種子發芽啟動所需時間較長,對照組在第40天左右種子才啟動萌發,而GA3處理石筆木種子24 h后,在27 d發芽啟動,6-BA處理的種子在第30~39天啟動發芽,表明GA3處理石筆木種子能促進其發芽,縮短萌發啟動時間。表4結果顯示,低濃度的GA3和6-BA有利于石筆木種子萌發,高濃度反而抑制種子萌發。與對照組相比,濃度為100 mg/L的GA3處理后的石筆木發芽率、相對發芽率、發芽勢、發芽指數顯著升高,其發芽率達到28.62%,表明GA3處理石筆木種子能提高其發芽率,但萌發率依然偏低,不能有效打破石筆木種子休眠。
3 結論與討論
山茶科種子屬于厚實種子,種皮堅硬。本研究中3種供試山茶科種子均具有堅實的外種皮,體積大而重,3種山茶科種子長度平均變異系數值相對較小,而種子寬度和厚度平均變異系數相對較大,與浙江紅山茶類似[27],博白大果油茶與多齒紅山茶種子形狀接近,均與石筆木種子形狀差異較大,可能是不同屬間差異所致。博白大果油茶種子質量最大,平均單粒種子重可達3.8 g左右(CV:0.035)。有研究發現,種子質量越大,其貯存內含物質越多,種子生命力越強[28],呂曉梅等對5種山茶屬種子萌發特性進行比較發現:長柱紅山茶重量最大,其發芽率、發芽指數、發芽勢均顯著高于其他4種山茶屬種子[16]。本試驗中,博白大果油茶種子啟動萌發時間早于多齒紅山茶和石筆木,且發芽率、發芽指數均高于后兩者,可能與博白大果油茶種子所含內含物質較多,為種子萌發過程提供營養物質和能量。而石筆木種子質量最小,所含內含物質最少,導致其萌動所需時間較長,發芽率偏低,萌發整齊度較差。
本研究發現,博白大果油茶的萌發抑制物質主要集中在種皮中,多齒紅山茶的萌發抑制物質主要集中在外種皮和胚中,且萌發抑制物活性較高,但在種子萌發實驗中發現,博白大果油茶和多齒紅山茶萌發率均較高,推測其可能與萌發過程中萌發抑制物質容易向外溶解有關,從而解除其對種子萌發的抑制作用。石筆木的種皮和胚提取液對白菜種子無明顯抑制作用,但其種子萌發率偏低,僅有12.67%,可能原因:一是抑制試驗所使用的萌發抑制物質濃度為5%,所含活性物質濃度較低;二是種子萌發過程與種皮厚度、致密度密切相關[29],有研究發現,堅硬的種皮是阻礙山茶科種子吸水的重要因素[16],去除種皮后的厚殼紅瘤果茶種子,發芽率明顯提高,萌發時間提前[30]。石筆木的種皮厚度和質密度較大,對石筆木種子吸水和萌動有一定的阻礙作用,從而導致石筆木發芽率偏低。
種子的休眠和萌發實際是促萌發因子與抑制萌發物質相互競爭的結果[26]。植物激素ABA(脫落酸)是促種子休眠的重要因子之一,其間接參與調控和維持種子休眠[31]。而GA3是萌發的主要促進因子,在種子萌發過程中與ABA互為拮抗關系,可解除種子休眠狀態,促進種子萌發[31-32]。此外,細胞分裂素能促進多種物種解除休眠,與GA協同促進種子萌發[31]。大量研究發現,外源添加GA3、6-BA可提高種子萌發效果,減少萌發時間[26,33-36]。本研究中,較蒸餾水浸種處理,GA3、6-BA浸泡博白大果油茶種子,發芽率、發芽整齊度有所上升,且發芽時間提前。同樣處理多齒紅山茶發現,GA3浸泡種子后,能促進種子萌發,發芽時間提前,而6-BA則對種子萌發作用不大。有研究發現,貴州石筆木種子存在生理后熟現象,用100 mg/L 6-BA和300 mg/L GA3共同浸泡24 h,種子活力上升,且發芽率達 96.7%[37-38],有效打破其休眠。本試驗中,低濃度的GA3能提高石筆木的發芽率、發芽整齊度,且發芽時間提前,但發芽率依然偏低,可能是石筆木同樣存在生理后熟現象,需進一步對其進行研究。
