4種濕地植物-微生物燃料電池產電性能比較
2019-03-07 06:21:16劉海林陳金峰鄒春萍
廣東農業科學 2019年12期
關鍵詞:植物
劉海林,陳金峰,鄒春萍
(廣東省農業科學院環境園藝研究所/廣東省園林花卉種質創新綜合利用重點實驗室/農業農村部華南都市農業重點實驗室,廣東 廣州 510640)
【研究意義】植物微生物燃料電池(Plant-Microbial Fuel Cell,P-MFC)以植物光合作用產生的根系分泌物作為土壤產電微生物的電子供體,將太陽能持續不斷地轉化為電能,是一種能耗少、無污染、持續性好、具有自我修復能力、可再生的綠色能源。它將植物的光合作用與微生物燃料電池技術相結合,在可再生能源開發、水污染治理、生態環境修復等方面顯現出廣闊的應用前景,并作為一種有效緩解能源危機和環境壓力的新技術受到越來越多國家的關注[1-3]。【前人研究進展】P-MFC研究中采用的植物以濕地植物居多[4],目前已有關于水稻(Oryza sativa)、大米草(Spartina anglica)、野古草(Arundinella anomala)、蘆竹(Arundo donax)、短葉茳芏(Cyperus malaccensisvar.brevifolius)、美人蕉(Canna indica)、風車草(Cyperus alternifolius)、菖蒲(Acorus calamus)等P-MFC的研究報道[5-8]。然而,已報道的P-MFC產電性能仍相對較弱,其最大輸出功率密度僅為222 mW/m2[5]。為適應濕地厭氧環境,濕地植物根系一般具有滲氧的生理特性。根系滲氧能在P-MFC陽極形成微氧環境,氧氣可與電極爭奪電子,從而影響P-MFC的產電性能[9]。自然狀態下,P-MFC陰極會自發形成菌藻膜[10],菌藻膜進行光合作用,吸收水體中的HCO3-釋放OH-和O2[11],改變陰極pH和氧化還原條件,也可能會影響陰極的電極反應。另外,環境溫度變化影響植物的生長和微生物的代謝活性,從而也會對P-MFC的產電性能產生較大影響[4]。目前,對于以上因素如何影響P-MFC產電性能尚缺乏進一步的了解。【本研究切入點】研究不同植物及環境因素對P-MFC產電性能的影響,對提高P-MFC產電能力、開發利用綠色能源具有重要意義。【擬解決的關鍵問題】本研究以巴拉草(Brachiaria mutica,禾本科)、短葉茳芏(Cyperus malaccensisvar. brevifolius,莎草科)、多枝扁莎(Pycreus polystachyus,莎草科)、雙穗雀稗(Paspalum paspaloides,禾本科)4種野外常見且耐水淹能力比較強的濕地植物為材料構建P-MFC,研究了不同P-MFC產電性能的差異及其產電性能與環境因素和植物特性之間的關系,為構建高效P-MFC、P-MFC產電性能機理的研究及相關產品的研發提供參考。
1 材料與方法
1.1 4種濕地植物P-MFC的構建
采用無離子交換膜的單室P-MFC為模型,以頂部直徑18 cm、底部直徑14.5 cm、高18.5 cm、總容積4 L的黑色塑料桶作為反應器,直徑14 cm、厚5 mm石墨碳氈作為陰、陽電極,電極間距5 cm,鈦絲連接電路[12]。陽極位于反應器的底部,覆泥土5 cm,栽種植物后將陰極置于土層表面,注滿水(圖1)。反應器中分別單獨栽種巴拉草(B. mutica)、短葉茳芏(C. malaccensis)、多枝扁莎(P. polystachyus)、雙穗雀稗(P. paspaloides)4種植物,每種植物3次重復,每個重復中栽種數量相等、健康無病害且生長狀況基本一致的植株,以無植物MFC作空白對照。啟動所有電池,并置于室外自然條件下預培養30 d,待開路電壓穩定后,外接1 kΩ電阻,開始監測數據,每隔10 min檢測和記錄1次電壓值和溫度值,連續監測211 d(2016年10月24日至2017年5月28日)。
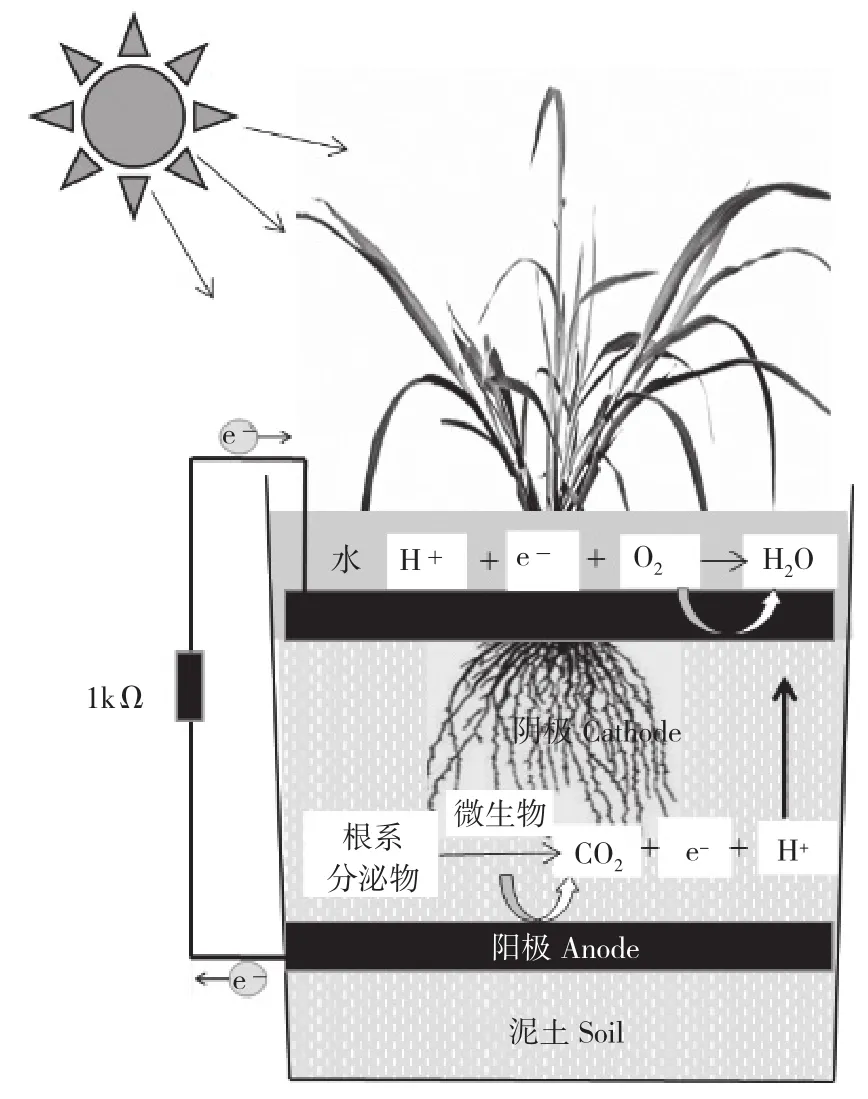
圖1 植物-微生物燃料電池結構示意圖Fig. 1 Schematic diagram of the P-MFC system
1.2 產電性能相關參數測定
1.2.1 電壓和電極測定 采用Keithley 2700萬用表(美國Keithley儀器公司)實時監測P-MFC輸出電壓,設置每隔10 min記錄一次數據。6 mm銀/氯化銀參比電極(上海楚兮實業有限公司)測定電池兩極電位,SG98多參數測試儀〔梅特勒-托利多(上海)有限公司〕測定陰極附近水體pH和溶解氧濃度。GSP-6型溫濕度記錄儀(杭州精創電子有限公司)實時記錄外界環境溫度值。輸出功率密度(Output Power Density,mW/m2)按以下公式計算:
輸出功率密度=U2/RS
式中,U為外接電阻兩端的電壓,R為外界電阻1 kΩ,S為陰極表面積(m2)。
每天8:30、13:30、18:00分別測定陰極區pH、溶解氧濃度,連續測定7 d。
1.2.2 根系滲氧量和滲氧率測定 采用檸檬酸鈦比色法[13]測定根系滲氧量。選用株高30 cm左右、根長約10 cm的4種植物單株分別置于營養液(0.1%瓊脂+10%Hoagland溶液)中,每組設3次重復,培養30 d。取50 mL潔凈試管,將連接氮氣瓶的導管插入試管底部持續吹氮排空氣,保證無氧環境;然后注入30 mL無氧營養液,再將用去離子水洗凈的植物根系轉入試管中,注入5 mL無氧檸檬酸鈦(Ⅲ)溶液,整個操作在持續通氮氣條件下進行;最后注入適量石蠟油密封液面,停止通氮氣。以無植物組作對照。將所有試樣及對照在光照培養箱中培養6 h,UV Power PC型紫外分光光度計(北京萊伯泰科有限公司)測定其在527 nm處的吸光度,同時以濃度1~20 mmol/L的無氧檸檬酸鈦(Ⅲ)溶液繪制標準曲線。計算根系滲氧量(ROL)和滲氧率(ROL rate):
ROL(μmol/d·株)=4C×(Y-Z)
ROL rate(μmol/d·g)=4C×(Y-Z)/m
式中,C表示每個試管加入檸檬酸鈦(Ⅲ)后的溶液總體積(L),Y表示對照組反應6 h后檸檬酸鈦(Ⅲ)濃度(μmol/L),Z表示放入植物的試管反應6 h后檸檬酸鈦(Ⅲ)濃度(μmol/L),m表示植物根系干重(g)。
1.2.3 根系孔隙度測定 采用25 mL比重瓶法測定根系孔隙度,計算公式如下:
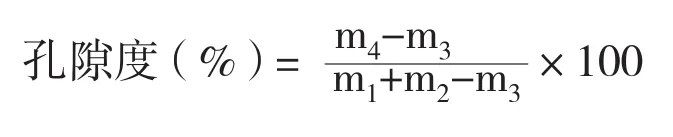
式中,m1為比重瓶+水的質量(mg),m2為根樣鮮質量(mg),m3為研磨前比重瓶+水+根樣的質量(mg),m4為研磨后比重瓶+水+根樣的質量(mg)[13]。
1.2.4 石蠟切片觀測 手工制作根系石蠟切片,利用HM340E型半自動石蠟切片系統(德國Microm公司)切片后用DM2500型正置顯微鏡(德國Leica儀器有限公司)進行觀察和拍照。
1.3 數據處理與分析
試驗數據用Microsoft Excel 2007進行統計分析,并結合Origin Pro 8.0軟件進行數據處理和繪圖,SPSS Statistics 19.0軟件對數據進行方差分析,采用LSD法多重比較進行差異顯著性測驗。
2 結果與分析
2.1 4種植物P-MFC電壓及電位變化情況
截取自2016年10月31日至2017年3月21日共142 d電池電壓較為穩定的時間段進行數據分析,其中2017年1月24—25日因停電數據缺失,結果(圖2,封二)顯示,P-MFC及空白對照的電壓整體上呈穩定的周期性波動,與溫度波動基本一致。10月至次年1月上旬平均氣溫高,電池平均電壓較高;1月中旬至2月上旬平均氣溫下降,電池平均電壓有所下降。雙穗雀稗的P-MFC平均電壓最高,為0.26(±0.02) V,其次是巴拉草、對照、短葉茳芏、多枝扁莎(表1)。從本研究結果來看,不同植物構建的P-MFC產電性能差異較大,禾本科的兩種植物優于莎草科的兩種植物,雙穗雀稗平均功率密度最大,為4.38(±0.56)mW/m2,多枝扁莎最小,為0.20(±0.08) mW/m2。
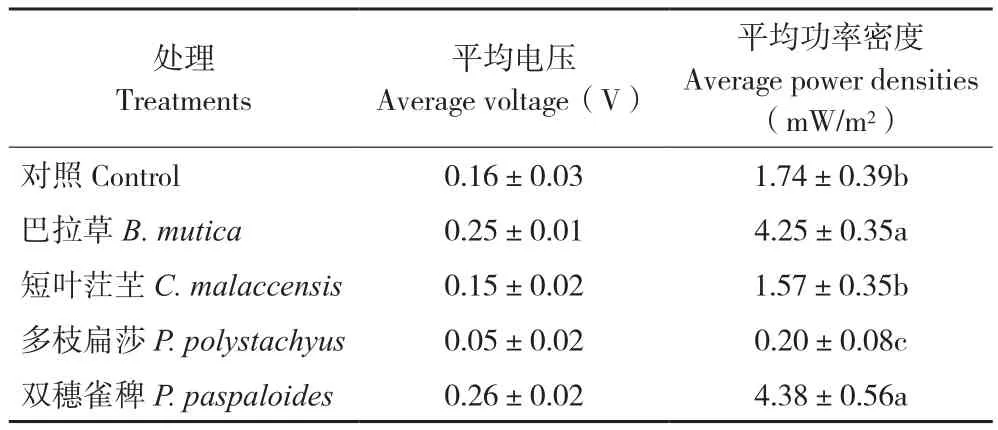
表1 4種植物P-MFC平均電壓和平均功率密度Table 1 Average voltage and average power densities of four P-MFCs
隨機選取3個晴朗日測定不同植物P-MFC的兩極電位和電壓,記錄時間為當天5: 00至次日4: 30,每隔30 min測定一次數據。結果(圖3、圖4)顯示,不同植物P-MFC的陰極平均電位差別不大,但陽極平均電位差別明顯。陰極日平均電位表現為雙穗雀稗(0.48V±0.08 V)>多枝扁莎(0.47V±0.01 V)>短葉茳芏(0.42V±0.02 V)>巴拉草(0.38V±0.01 V)>對照(0.02V±0.07 V),而陽極日平均電位表現為多枝扁莎(0.46V±0.01 V)>雙穗雀稗(0.32V±0.11 V)>短葉茳芏(0.30V±0.05 V)>巴拉草(0.12V±0.03 V)>對照(-0.12V±0.04 V)。圖5顯示,盡管P-MFC陰陽極平均電位均高于對照,但平均電壓并不都高于對照,表現為巴拉草(0.26V±0.02 V)>雙穗雀稗(0.17V±0.07 V)>對照(0.1V6±0.03 V)>短葉茳芏(0.12V±0.03 V)>多枝扁莎(0.02V±0.02 V)。
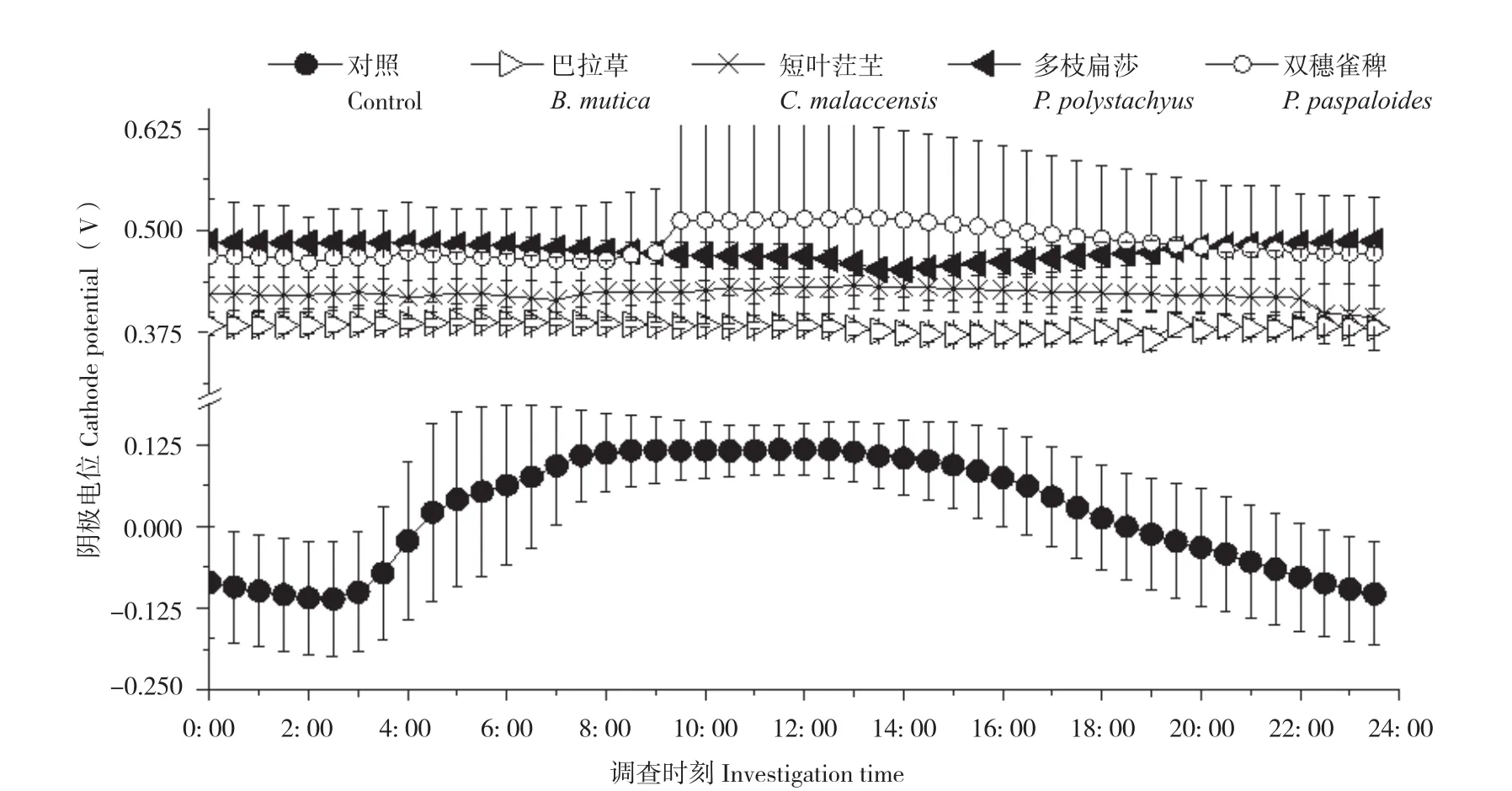
圖3 4種植物P-MFC陰極電位日變化Fig. 3 Daily variation of cathode potential of four P-MFCs

圖4 4種植物P-MFC陽極電位日變化Fig. 4 Daily variation of anodic potential of four P-MFCs
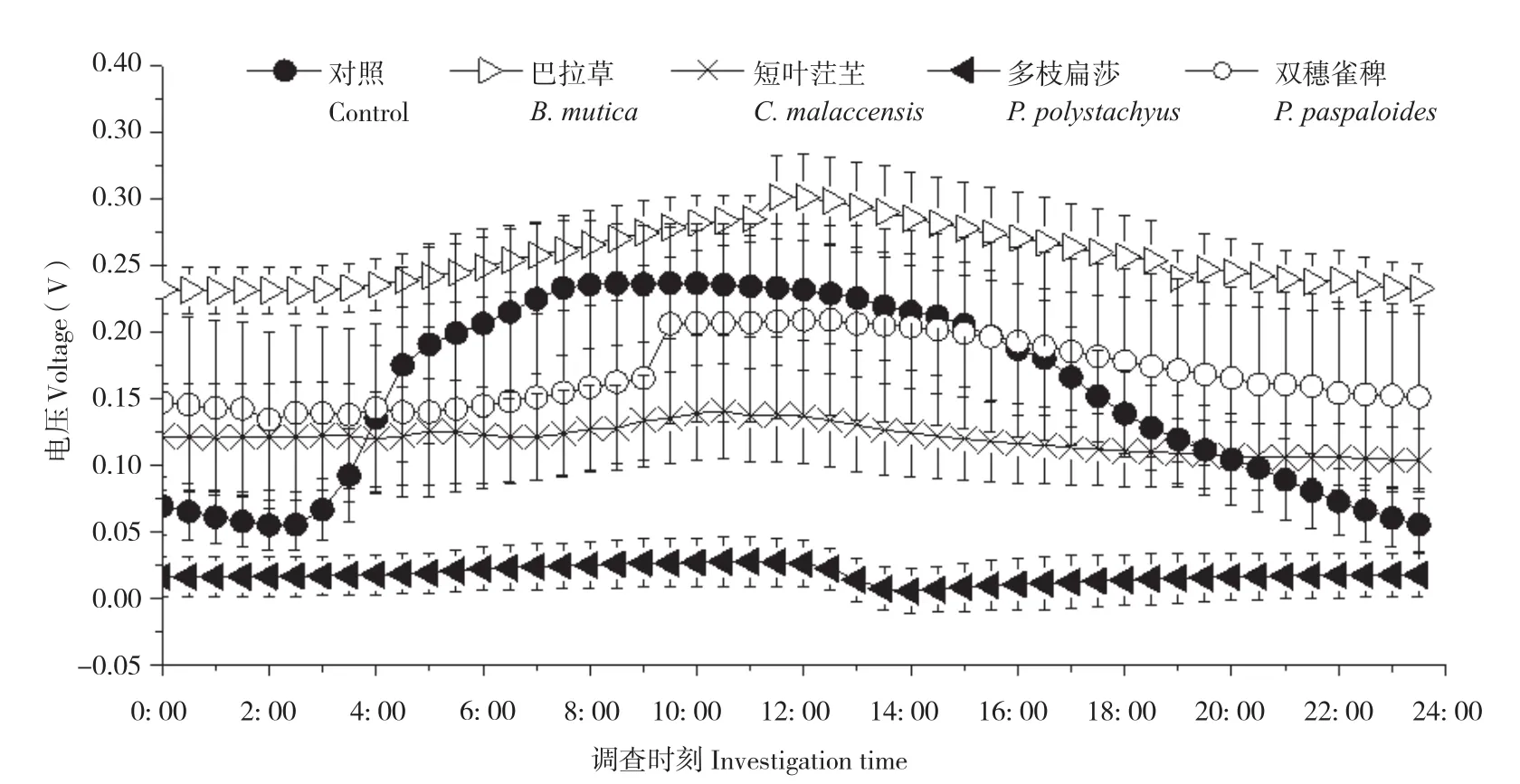
圖5 4種植物P-MFC電壓日變化Fig. 5 Daily variation of voltage of four P-MFCs
2.2 4種植物P-MFC陰極區pH值及溶氧濃度
從圖6、圖7可以看出,P-MFC及對照的陰極區pH值、溶氧濃度變化趨勢均呈單峰型,最高值出現在13: 00—14: 00之間。P-MFC陰極區pH(7.2~8.5)、溶氧濃度(4.83~10.81 mg/L)均低于對照(8.6~9.9、6.28~15.88 mg/L),其中pH值變化情況表現為巴拉草7.3-8.5-7.4、短葉茳芏7.3-7.5-7.2、多枝扁莎7.2-8.5-7.3、雙穗雀稗7.2-7.2-7.3、對照8.6-9.9-9.4,溶氧濃度變化情況表現為巴拉草5.73 mg/L-10.26 mg/L-6.92 mg/L、短葉茳芏5.21 mg/L-8.11 mg/L-5.65 mg/L、多枝扁莎7.55 mg/L-10.81 mg/L-5.64 mg/L、雙穗雀稗5.15 mg/L-5.66 mg/L-4.83 mg/L、對照6.28 mg/L-15.88 mg/L-10.56 mg/L。其中,多枝扁莎P-MFC陰極區pH值、溶氧濃度均值最大,而雙穗雀稗則最小。

圖6 4種植物P-MFC陰極區pH值變化Fig. 6 Variation of pH in cathode area of four P-MFCs
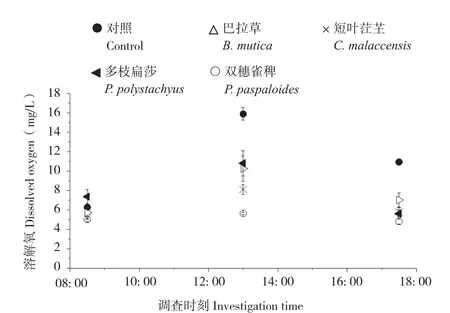
圖7 4種植物P-MFC陰極區溶解氧濃度Fig. 7 Variation of dissolved oxygen concentration in cathode area of four P-MFCs
2.3 4種植物根系結構及滲氧率
2.3.1 根系結構和孔隙度 通過根系石蠟切片結構觀察,發現不同植物根系內均含有氣腔,說明根系均具有一定的通氣能力(圖8),巴拉草、雙穗雀稗和多枝扁莎的根系上具有較多細小根毛,而短葉茳芏根系根毛較少。采用比重法測定得到的根系孔隙度由小到大依次為巴拉草、短葉茳芏、雙穗雀稗、多枝扁莎(表2)。
2.3.2 根系滲氧率 由表3可知,不同植物種類的根系滲氧率存在差異。巴拉草和短葉茳芏相比其他植物有著更低的滲氧率,而多枝扁莎的滲氧率較高,僅次于菖蒲。滲氧率大小表現為短葉茳芏<巴拉草<雙穗雀稗<多枝扁莎。

圖8 4種植物根系形態和解剖結構比較Fig. 8 Root morphological and anatomic structure of four different plants
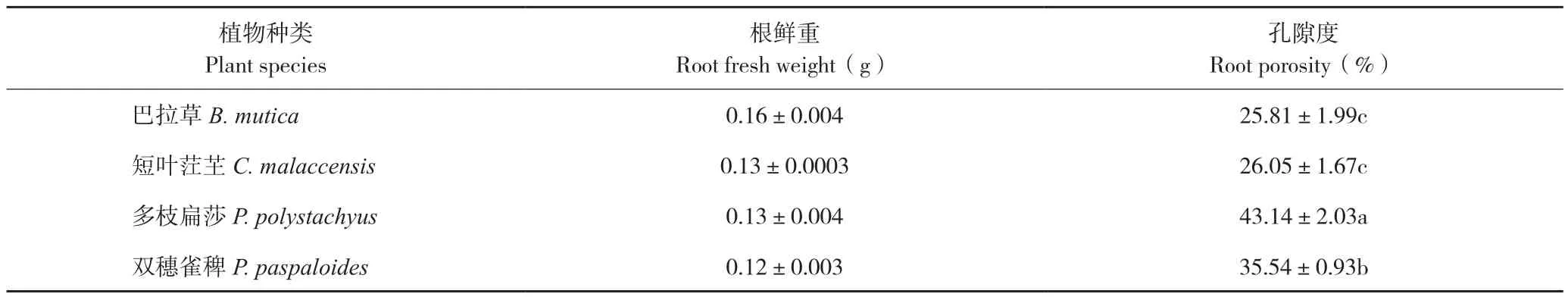
表2 4種植物根系孔隙度比較Table 2 Comparison of root porosity of four different plants
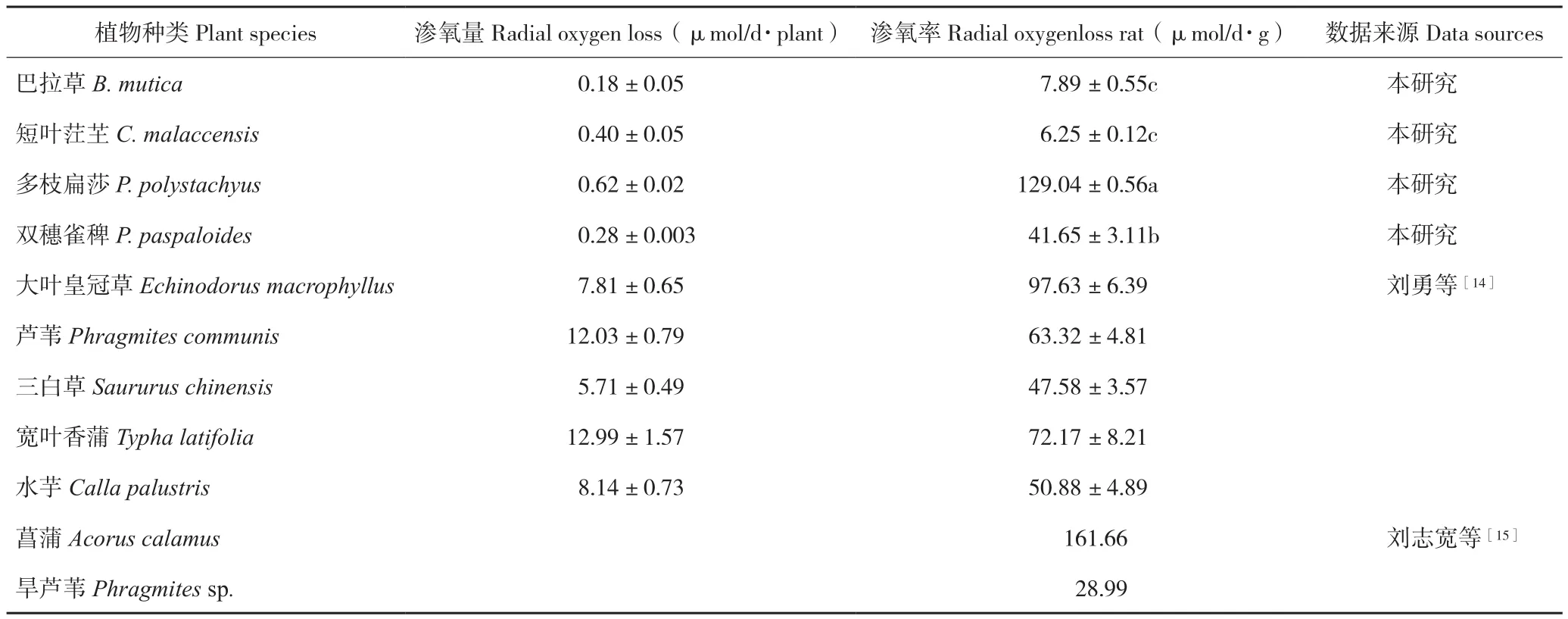
表3 4種植物根系滲氧率比較Table 3 Comparison of ROL rate of four different plants
3 討論
3.1 不同植物P-MFC產電特性
Helder等[5]以禾本科植物大米草(Spartina anglica)、野古草(Arundinella anomala)和蘆竹(Arundo donax)分別構建P-MFC,其產電性能差異明顯,大米草和野古草P-MFC的最大功率密度和平均功率密度分別為222 、10 mW/m2和22 、21 mW/m2。吳夏芫等[16]以粉黛萬年青(Dief fenbachia)、小白掌(Spathiphyllum kochiiEngl. & K. Krause)、金邊富貴竹(Dracaena sanderiana)構建了植物-沉積物微生物燃料電池(基質為土壤),最大功率密度分別為8.7、14.0、13.0 mW/m2。可見,在同樣條件下,采用不同植物構建的P-MFC產電性能是有差異的。Timmers等[17]報道以大米草構建的含離子交換膜的雙室P-MFC(陽極基質為霍格蘭溶液)最大平均功率密度為22(±14)mW/m2;而Helder等[5]同樣以大米草構建的P-MFC產電性能(222 mW/m2)要遠高于Timmers等的研究結果[17],這與二者P-MFC所用的電極材料及陽極體積不同有關[5]。可見,同一種植物采用不同的電極材料和構建P-MFC方法,其產電性能也是有差異的。朱娟平等[8]以風車草(Cyperus alternifolius)和短葉茳芏構建的河涌底泥P-MFC最大功率密度為10.74、10.05 mW/m2。而本研究中4種植物P-MFC平均功率密度均低于上述報道,但不同P-MFC間產電性能差異明顯,其中巴拉草和雙穗雀稗P-MFC產電性能優于對照,而短葉茳芏和多枝扁莎P-MFC則低于對照。總之,選用合適的植物和優化電池構造有利于提高P-MFC的產電性能。
3.2 植物根系結構和滲氧能力對P-MFC產電性能的影響
濕地植物為適應濕地厭氧環境一般都具有根系滲氧的生理特征[18-19]。濕地植物根系滲氧率與通氣組織的發達程度、根部細胞的排列緊密程度有關,一般通氣組織越發達,根部細胞排列疏松,根系孔隙度越大,根系滲氧率越大[20-22]。因此,可通過觀察根系的解剖結構和測定根系的孔隙度研究植物根系的滲氧能力。本研究中巴拉草和短葉茳芏的根系細胞排列相對緊密、孔隙度小,因而具有較低的滲氧率;而多枝扁莎和雙穗雀稗根系細胞排列疏松、孔隙度大,滲氧率高。當數據監測結束后,取出多枝扁莎P-MFC電極,發現植株根系已遍布整個反應器并穿透陽極,在陽極上表面和反應器內壁各形成一層較其他P-MFC更明顯的紅棕色氧化鐵膜,由此也反映出多枝扁莎根系具有更強的滲氧能力。與劉志寬等[15]和劉勇等[14]報道的其他濕地植物的滲氧率比較發現,多枝扁莎的滲氧率也屬于高水平。
已有報道表明,植物根際滲氧會為根際非產電微生物氧化降解根系分泌物提供電子受體[9],進而提高P-MFC陽極電位,降低其產電性能[23]。因而,我們推測不同濕地植物P-MFC產電性能的差異可能與根系滲氧能力有關:巴拉草根系滲氧率較低,從而能夠維持較低的陽極電位,而多枝扁莎根系滲氧率最高,維持了較高的陽極電位,在二者陰極電位差別不大的情況下,高的陽極電位減弱了輸出電壓,降低了電池的產電性能。可見,根系滲氧能力對P-MFC的產電性能具有明顯影響,可作為植物篩選時的一項重要參考依據。
3.3 陰極區pH和溶氧濃度對P-MFC產電性能的影響
本研究除雙穗雀稗陰極pH變化較為平穩外,其他P-MFC陰極pH在白天的變化呈單峰型,這可能主要是因為陰極表面菌藻膜的光合作用消耗了水體中大量HCO3-,釋放OH-,造成水體pH隨著光照升高[11]。雙穗雀稗P-MFC植物覆蓋茂密,陰極電極上藻類少,pH相對穩定。另外,陰極O2的還原過程也需要H+參與,白天P-MFC運行效率高,H+被大量消耗、濃度降低,pH增大。夜晚,藻類不再消耗水體中的HCO3-;同時陽極產電微生物代謝產生的H+擴散至陰極,補充了陰極上的損耗,使陰極區H+濃度逐漸升高,pH降低。吳瑾妤等[24]報道通過調節陰陽兩極pH可以優化電池的產電性能,陽極堿性(pH 8.5)和陰極酸性(pH5.5)時,沉積物微生物燃料電池(SMFC)輸出功率最高。本研究中雙穗雀稗P-MFC陰極pH穩定且相對較低,能為陰極O2的還原提供充足的H+,從而使其陰極電位較高,整體輸出功率較大。
除雙穗雀稗P-MFC外,其他P-MFC陰極以及對照表面均有一層菌藻膜,菌藻膜進行光合作用致使陰極溶解氧白天升高夜晚降低。陰極充足的氧氣濃度有利于提高MFC的產電性能[25]。巴拉草和多枝扁莎陰極的溶解氧濃度最高,但是巴拉草的陰極電位反而最低,這可能是因為陰極電位是氧氣、pH和微生物活性等共同作用的結果。
3.4 溫度對P-MFC產電性能的影響
本研究發現P-MFC的輸出電壓變化趨勢與溫度波動趨勢基本一致,溫度與P-MFC產電性能之間可能存在一定關系。10—11月平均氣溫高,電池平均輸出電壓較高,而此時期植物生長旺盛,株高增長明顯,根系和產電微生物代謝活躍;12月至次年3月,氣溫逐漸下降,電池平均輸出電壓下降,而此時期植物生長緩慢,根系和產電微生物代謝活動較弱,這與Zhou等[7]觀察到的現象類似。可見,溫度可能通過影響植物和微生物的生理代謝活性間接影響P-MFC的產電性能[4]。若將溫度控制在適宜范圍內,植物生長良好,根系分泌物豐富,陽極產電微生物代謝活躍,產生的質子和電子量多,將有利于陰極還原反應,從而使電池具備較高的產電性能。溫度對產電性能的影響及二者間的相關性,有待進一步研究。
4 結論
不同植物構建的P-MFC產電性能差異明顯,種植植物的P-MFC電位高于無植物的對照。巴拉草和雙穗雀稗P-MFC產電能力較強,可能與巴拉草根系滲氧速率較低、雙穗雀稗陰極pH較低且比較穩定有關。低的根系滲氧能力能夠維持低的陽極電位,陰極低pH環境有利于維持較高的陰極電位,從而使得電池整體輸出電壓較高。在利用濕地植物構建P-MFC時,植物的根系特點尤其是根系滲氧生理是需要重點考慮的因素。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
