新型鴨呼腸孤病毒廣東分離株σC蛋白基因序列分析
2019-02-17 03:21:58張思遠盧秀嫻梁昭平潘俊斌林舉攀葉賀佳
中國獸藥雜志 2019年12期
張思遠,盧秀嫻*,云 驁,梁昭平,潘俊斌,林舉攀,葉賀佳
(1.廣州市華南農大生物藥品有限公司,廣州510300;2.華南農業大學,廣州510642)
近年來,一種新型鴨呼腸孤病毒(NDRV)疾病,稱為北京鴨“脾壞死”病,在我國多個地區流行,該病的主要特征病變為肝、脾臟不規則出血、壞死與心肌、腔上囊出血,發病沒有明顯季節[1],與傳統的番鴨呼腸孤病毒(MDRV)相比,具有更廣泛的宿主,不同品種鴨均易感,發病率與病死率有較大差異,被感染的病鴨日齡越小,發病、病死率越高[2]。
2018年下半年,廣東湛江某肉鴨養殖場櫻桃谷肉鴨發病,臨床主要表現為腳軟、關節腫大等臨床特征,解剖發現主要為肝臟腫大、脾臟有壞死灶等病變特征,該肉鴨群主要是1~2周齡的雛鴨發病,最早可見于3~5日齡,發病率60%以上,死亡率40%左右。根據臨床診斷初步懷疑該病是由鴨呼腸孤病毒引起的疾病,但該病所引起的死亡率卻不同于經典的北京鴨“脾壞死”病。本研究對采集的病死鴨病料進行了病原分離鑒定,對分離株的σC蛋白基因進行了克隆、測序和序列基因遺傳變異分析,以期了解該病毒的變異情況,為該病的防控和診斷提供參考依據。
1 材料和方法
1.1 材料 病料為廣東湛江某肉鴨養殖場臨床發生“肝脾壞死癥”死亡雛鴨的肝臟和脾臟;50枚9日齡健康櫻桃谷鴨胚購自廣東某健康種鴨養殖場,40只1日齡健康櫻桃谷鴨購自廣東某健康種鴨養殖場。
1.2 主要試劑 核酸提取試劑盒(9766)、pMD19-T載體(6013)購自 TaKaRa公司、DL2000 DNA Marker(BM101-01)購自北京全式金公司;2×Gotap Master Mix(M711)購自Promega公司;核酸染料(RT210)和反轉錄試劑盒(KP103)均購自TIANGEN公司。
1.3 病料處理 將病料剪碎研磨,與PBS按1∶3的比例制成組織勻漿液,將勻漿液置于冰箱-20℃冷凍并反復凍融3次,離心后取上清液過濾除菌、分裝,分裝量為1 mL/管,于-70℃及以下保存。
1.4 病原傳代培養 病料濾液經尿囊腔接種9日齡鴨胚,0.2 mL/胚,收集接種后24~108 h死亡鴨胚,收獲胚液,繼續傳代至鴨胚規律性死亡止。檢測胚液對1%雞紅細胞懸液的血凝性。
1.5 RT-PCR檢測 分別用以下4對引物對病料濾液及收獲的胚液體進行RT-PCR檢測。根據GenBank中已發表的番鴨細小病毒(MPV)、新型鴨呼腸孤病毒(NDRV)、番鴨呼腸孤病毒(MDRV)、鴨瘟(DPV)的基因序列,用Oligo7.0軟件設計特異性檢測引物,由上海生工生物工程有限公司合成,引物序列見表1。

表1 RT-PCR檢測引物Tab 1 RT-PCR detection primers
1.6 組織切片病理觀察 患鴨的脾臟和肝臟固定于10%的多聚甲醛溶液48 h,經脫水、透明、石蠟包埋、切片、H.E.染色,觀察病理變化。
1.7 對鴨胚的半數致死量 胚液的 10-2、10-3、10-4和10-5稀釋度分別經尿囊腔接種9日齡鴨胚各6枚,0.2 mL/胚,用蠟封孔,于37℃靜置孵育,記錄各稀釋度24~108 h死亡鴨胚數,按Reed-Muench法計算ELD50。
1.8 動物回歸實驗 40只1日齡健康櫻桃谷鴨,隨機分為2組,攻毒組30只經皮下接種病毒液,接種劑量0.5 mL/只;對照組10只以同樣的方式和劑量接種PBS。每日觀察雛鴨的臨床癥狀和死亡情況,觀察10日后撲殺解剖2組雛鴨。
1.9 分離毒σC蛋白基因序列測定 根據σC基因兩端外的保守序列設計1對擴增σC全長基因的引物,引物序列:P1(上游):5'-TTGAAAACTGAACAAAAGA - 3',P2(下游):5'- CCATAACTAA-CATAAGGGCA -3',預期擴增片段大小為 986 bp。經RT-PCR擴增σC基因全序列,回收RT-PCR產物,與pMD19-T載體相連轉化至DH5α,經PCR鑒定后挑取陽性菌送華大基因公司廣州測序部進行基因全長序列的測定。
1.1 0病毒σC蛋白基因序列分析 利用DNAStar等軟件將病毒株基因序列與GenBank中登錄的禽源呼腸孤病毒參考毒株進行比較。
2 結果與分析
2.1 病毒分離 將病料接種鴨胚,培養96~108 h后胚體死亡,胚胎體表充血、出血;死亡胚尿囊液清亮,胚胎蜷縮,見圖1。該分離毒不能凝集1%雞的紅細胞。
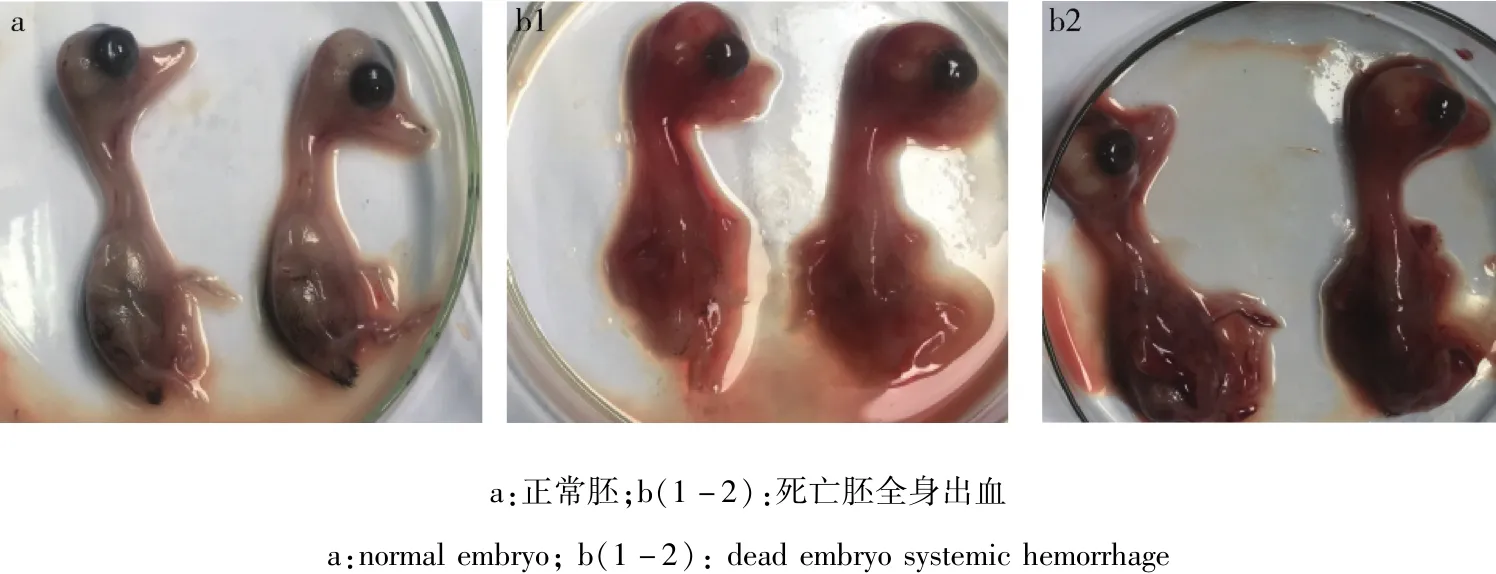
圖1 死亡鴨胚胚體變化Fig 1 Death embryo body changes
2.2 RT-PCR檢測結果 RT-PCR鑒定應用NDRV特異性引物能從病料濾液和分離的胚液種擴增到500 bp目的片段(圖2),應用其他3種鴨的病毒特異引物不能從病料中擴增到目的片段,均為陰性。分離到的毒株命名為GD693。

圖2 分離毒株RT-PCR檢測結果Fig 2 RT-PCR results of isolated strains
2.3 組織切片病理觀察 脾紅白髓界限不清,組織結構崩解,被膜下形成炎癥反應帶;肝臟組織間出現大量的紅細胞、出現局灶性壞死,空泡變性(圖3)。
2.4 病毒半數鴨胚致死量(ELD50) 測定結果為:ELD50=10-4/0.2 mL。
2.5 動物回歸實驗 攻毒組鴨在接毒的第5天開始出現精神萎靡,排白色稀糞,食欲減退,觀察期滿10 d后剖檢發現鴨出現與臨床自然發病櫻桃谷鴨一致的病理變化(圖4);第2組對照無異常。
2.6 σC蛋白基因片段RT-PCR擴增結果 結果顯示:NDRVσC蛋白基因擴增的片段長度約986 bp,與預期的結果一致(圖5)。
2.7 σC蛋白基因序列測定與遺傳進化分析 σC蛋白基因開放閱讀框(ORF)為966 bp,編碼321個氨基酸,是新型鴨呼腸孤病毒S1基因組第3個ORF編碼的。GD693株與參考毒株σC基因核苷酸序列及推導氨基酸序列同源性比較結果表明,GD693株與 MDRV毒株D1546株的同源性為60% ~61%之間;與 NDRV代表毒株 091、NP03、TH11株的同源性為96.7%~98.6%之間。
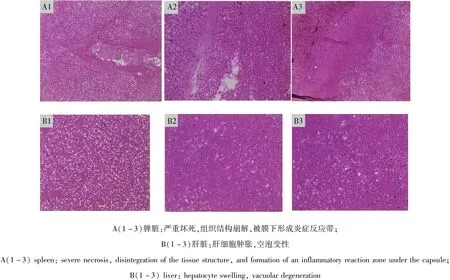
圖3 病死鴨脾臟、肝臟組織病理切片圖Fig 3 Histopathological section of the spleen and liver of dead duck
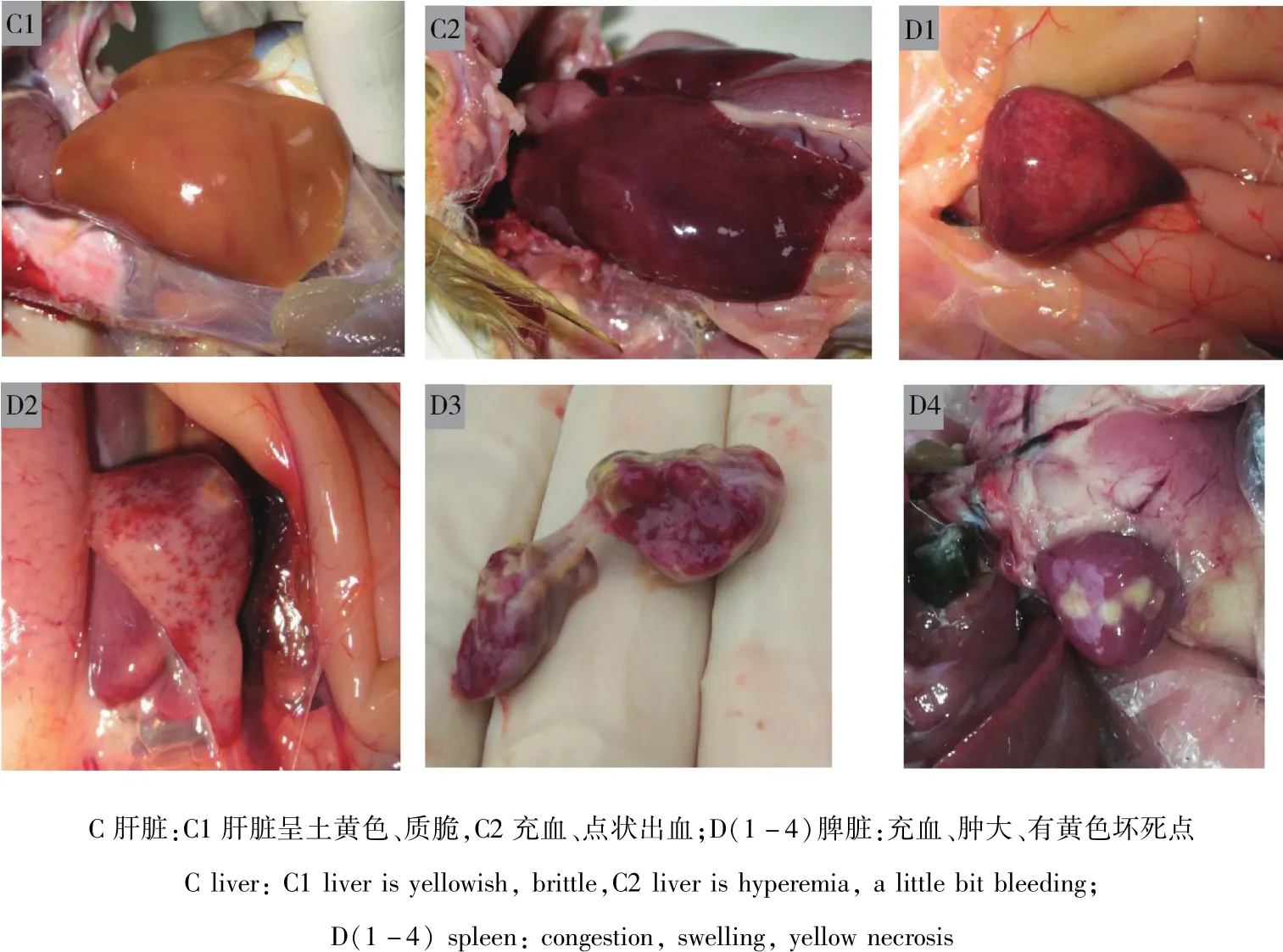
圖4 動物回歸實驗Fig 4 Animal regression experiment
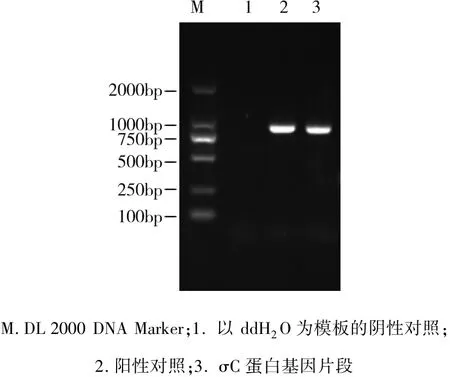
圖5 σC蛋白基因片段RT-PCR擴增結果Fig 5 RT-PCR results ofσC protein gene segment
將GD396株與NDRV代表毒株091、TH11株的σC蛋白基因序列進行比較,GD396株核苷酸及氨基酸序列存在差異位點,氨基酸差異位點分別位于第25(T→K)、64(E→D)、65(L→M)、91(L→L)、132(T→ S)、138(R→Q)、150(P→ S)位氨基酸。核苷酸和氨基酸的序列改變,有可能會導致毒力相關位點發生改變,從而使毒株的毒力發生變化。
各毒株σC蛋白基因遺傳進化樹分析結果(圖6)顯示,可將水禽源呼腸孤病毒分為兩個基因型,經典的MDRV和GRV屬于基因1型,而NDRV及N-GRV屬于基因2型。GD693毒株與NDRV代表毒株處于進化樹的同一大分支,但卻處于一個單獨的分支,同屬于基因2型,具有相近的遺傳演化關系;與其他毒株處于進化樹的不同分支,親緣關系相對較遠。
3 討論與結論
本研究從臨床表現為腳軟、關節腫大的病死鴨中分離到NDRV,該病以脾臟壞死,肝臟點狀或斑塊狀出血為主要特征的。經動物回歸試驗,可以復制出與臨床自然病死鴨相似的病變。NDRV屬于呼腸孤病毒科正呼腸孤病毒屬,沒有囊膜,呈二十四面體的雙層衣殼結構,核酸是線性雙鏈RNA,基因組由10個雙鏈RNA基因片段組成,大片段(L1-L3)、中片段(M1 -M3)和小片段(S1 -S4)[3-4]。呼腸孤病毒在抗原致病性、基因編碼蛋白的順序及病毒基因間的同源性方面上完全不同于之前的MDRV 和雞呼腸孤病毒(ARV)[5-6],宿主更加廣泛,對各種品種鴨均有致病性[7],給我國水禽養殖業造成了一定程度的經濟損失。
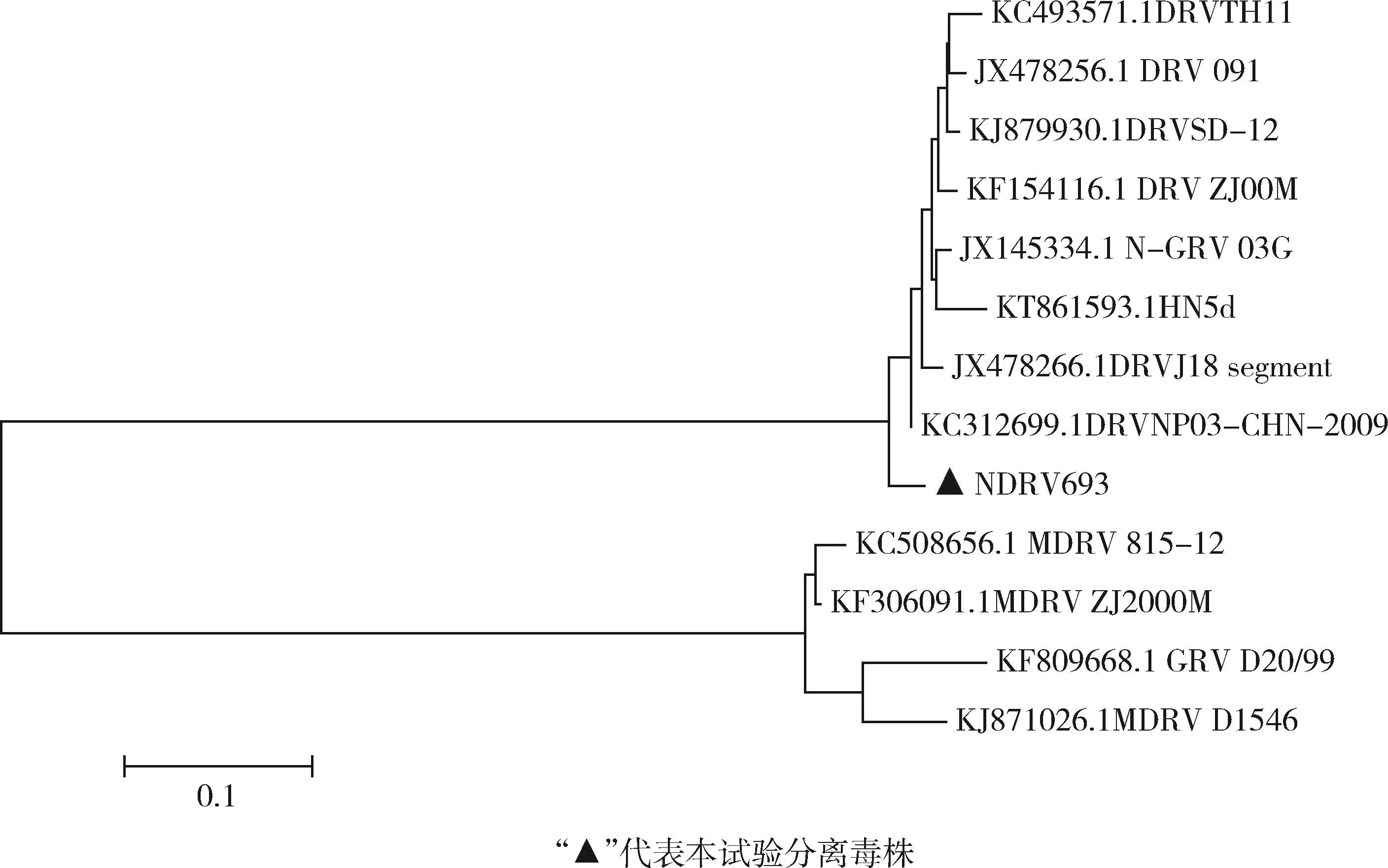
圖6 NDRV GD693株與參考毒株σC蛋白基因序列的遺傳進化樹Fig 6 Inheritance of NDRV GD693 strain and reference strainσC protein gene sequence tree
不同水禽源呼腸孤病毒之間存在較高的序列變異性,外衣殼蛋白(μB、σB和σC)編碼基因節段表現出更高的序列變異性,適合于不同禽源毒株的鑒別,其中變異性最大的是σC蛋白編碼基因,而它在病毒的致病過程中發揮重要作用[8]。σC蛋白是一種結構蛋白,是由NDRV S1基因編碼的,在呼腸孤病毒基因組編碼的所有σ類蛋白中是最小的,同時是病毒的主要免疫蛋白,負責宿主細胞的附著,通過受體介導的方式對靶細胞進行識別與結合來啟動病毒感染過程,可誘導機體產生特異性保護性中和抗體[9]。NDRV的σC基因在分子大小和基因同源性方面與雞呼腸孤病毒(雞病毒性關節炎)存在很大的差異[10]。
將GD693株的σC蛋白基因序列與參考毒株進行了比對分析,結果顯示,所分離的GD693株與MDRV毒株的同源性比較低(90% ~92%),而與NDRV代表毒株同源性則較高(95% ~97%)。遺傳進化樹顯示,GD693株與MDRV則處于不同的分支,而與NDRV代表毒株處于同一進化大分支,但又處于不同的小分支,表明可能它們來源于共同的祖先,但在病毒流行過程中形成了不同分支。將GD396株與NDRV代表毒株091、TH11株的σC蛋白基因序列進行比較,GD396株及氨基酸序列存在6個差異位點,可能會使病毒對鴨的致病性增強,據此推測GD693株可能是對鴨具有較高致病性的變異株,關于毒力的増強機制,仍需進一步研究。
本研究通過對NDRV的毒力、致病性及主要抗原σC蛋白的變異分析,為今后新型鴨呼腸孤病毒分子流行病學研究、疾病防控和診斷提供參考依據。疫苗免疫是預防本病的主要防控措施,但目前還沒有商品化的疫苗,因此研發新疫苗對該病的防控非常必要。
