酶法合成環(huán)肽研究進(jìn)展
2019-01-29 10:26:08程孝中周志昉吳志猛
食品與發(fā)酵工業(yè) 2019年1期
程孝中,周志昉,吳志猛
(江南大學(xué),糖化學(xué)與生物技術(shù)教育部重點(diǎn)實(shí)驗(yàn)室,江蘇 無(wú)錫,214122)
環(huán)肽化合物在自然界中普遍存在,與線性肽相比,它可以形成剛性構(gòu)象,提高了對(duì)蛋白酶降解的抵抗能力;某些環(huán)肽還具有獨(dú)特的細(xì)胞膜穿透能力;另外,小分子環(huán)肽對(duì)某些蛋白受體具有較高的親和力和選擇性[1]。環(huán)肽因其優(yōu)良的生物活性及構(gòu)象特征,在生物醫(yī)藥領(lǐng)域得到廣泛關(guān)注。自從1944年蘇聯(lián)科學(xué)家發(fā)現(xiàn)了第1個(gè)環(huán)肽短桿菌肽(gramicidin S)[2],這種治療傷口感染的環(huán)肽挽救了無(wú)數(shù)生命。至今為止,研究者從動(dòng)植物、微生物中均發(fā)現(xiàn)了環(huán)肽,例如,抗生素類的短桿菌酪肽A、乳酸鏈球菌肽;免疫抑制劑類的環(huán)孢菌素A;抗癌抗病毒因子類的萬(wàn)古霉素等[3]。
目前,環(huán)肽的合成方法主要有:(1)化學(xué)合成法。使用該法合成環(huán)肽需要側(cè)鏈全保護(hù)的線性多肽,當(dāng)環(huán)肽氨基酸數(shù)量增多,由于熵的壁壘增加,導(dǎo)致合成效率降低。近年來(lái)又發(fā)展了很多位點(diǎn)特異性的反應(yīng)用于環(huán)肽合成,如化學(xué)自動(dòng)連接反應(yīng)(native chemical ligation, NCL)[4],點(diǎn)擊化學(xué)(click chemistry)[5],酮酸-羥胺偶聯(lián)法(ketoacid-hydroxylamine ligation)[6],氮丙啶醛法(aziridine aldehydes-mediated macrocyclization)[7]等。(2)生物合成法。基于生物體內(nèi)轉(zhuǎn)肽機(jī)制發(fā)展起來(lái)的一些方法,例如利用內(nèi)含肽自我剪接的表達(dá)蛋白連接(expressed protein ligation,EPL)[8]、蛋白反式剪切(protein trans-splicing,PTS)[9],以及依賴體外的翻譯系統(tǒng)的遺傳密碼重編程(genetic code reprogramming),這類方法主要針對(duì)大分子蛋白的環(huán)化,一般都要進(jìn)行基因表達(dá)獲得線性底物。
酶催化的環(huán)肽合成是近年來(lái)興起的一種新型環(huán)肽合成方法,該法具有反應(yīng)條件溫和、特異性強(qiáng)、合成效率高等優(yōu)點(diǎn),另外酶催化的環(huán)肽合成還可以在肽鏈中引入非天然氨基酸或非氨基酸結(jié)構(gòu),進(jìn)而大大豐富了環(huán)肽結(jié)構(gòu)。本文就近幾年來(lái)發(fā)展的酶法合成環(huán)肽進(jìn)行綜述,本文涉及到的環(huán)肽包括均環(huán)肽和雜環(huán)肽,這些環(huán)肽的連接方式主要有首尾連接、側(cè)鏈與首或尾連接以及側(cè)鏈連接。
1 介導(dǎo)環(huán)肽合成的酶
1.1 分選酶Sortase A
分選酶A(Sortase A)是從金黃色葡萄球菌(Staphylococcusaureus)分離出來(lái)的1種轉(zhuǎn)肽酶,能特異地識(shí)別肽C端的保守序列LPXTG(Leu-Pro-X-Thr-Gly;X代表任1種氨基酸),并從蘇氨酸和甘氨酸之間使肽鍵斷裂,形成酰基酶-底物復(fù)合物,N端帶有多聚甘氨酸的親核基團(tuán)進(jìn)攻復(fù)合物,最終形成連接產(chǎn)物(圖1-a)[10]。該酶基因工程表達(dá)產(chǎn)率較高(232 mg/L),易于獲取,已廣泛地用于蛋白連接或蛋白標(biāo)記[11]、細(xì)胞表面的蛋白標(biāo)記[12]和蛋白環(huán)化[13]。
WU等系統(tǒng)性地探索了分選酶A催化多肽底物環(huán)化時(shí)肽鏈的最適長(zhǎng)度,發(fā)現(xiàn)線性底物肽最短為19個(gè)氨基酸(包括信號(hào)序列LPXTG和GG)時(shí),反應(yīng)產(chǎn)物主要是單環(huán)產(chǎn)物,化合產(chǎn)率達(dá)到80%;當(dāng)線性肽核心序列小于17個(gè)氨基酸(包括信號(hào)序列LPXTG和GG)時(shí),環(huán)化單環(huán)產(chǎn)物僅為6%,主要形成二聚體和三聚體環(huán)肽,而且底物濃度增大,二聚體環(huán)肽和三聚體環(huán)肽增多,單環(huán)產(chǎn)物相應(yīng)減少[14]。這種規(guī)律還和底物肽序列有關(guān),在利用分選酶A合成RGD環(huán)肽過(guò)程中,當(dāng)?shù)孜镫臑?6個(gè)氨基酸時(shí)(包括信號(hào)序列LPXTG和GG),主要產(chǎn)物仍是單環(huán)肽[15]。分選酶A通常在大環(huán)肽合成方面產(chǎn)率更好,在合成唾液肽組胺素(環(huán)肽為38個(gè)氨基酸)產(chǎn)率能夠達(dá)到90%[16],可能是因?yàn)榈孜镫逆溤介L(zhǎng),N端多聚甘氨酸在空間上更容易接近LPXTG。另外,分選酶A強(qiáng)大的環(huán)化能力還可以用來(lái)合成復(fù)雜的二硫鍵環(huán)肽(胰蛋白酶抑制劑SFT-1、芋螺毒素Vc1.1和抗病毒肽kB1),即使產(chǎn)物中引入了信號(hào)序列(LPXTG),并不影響其生物活性[17],例如利用該酶對(duì)人干擾素和生長(zhǎng)激素的環(huán)化[18-19]。ZHANG等拓展了分選酶A的應(yīng)用范圍,在環(huán)肽SFT-1非保守區(qū)域嫁接不同功能的小肽基序,賦予SFT-1更多的生物功能,形成特異的配體嫁接環(huán)肽可以調(diào)節(jié)蛋白質(zhì)之間的相互作用[20]。

圖1 分選酶A介導(dǎo)的環(huán)肽合成(a);N端不同的甘氨酸模擬物抑制分選酶A環(huán)肽合成(b)中的可逆反應(yīng)
Fig.1 Sortase A-mediated cyclization of peptides(a),glycine mimics at N terminus inhibit reversible reactions of sortase A -mediated cyclization of peptides(b)
分選酶A催化效率在同類酶中較低(與底物的摩爾比例為0.1~1.0),因此反應(yīng)需要更長(zhǎng)的時(shí)間(>20 h)和更高的酶量。另外,分選酶A催化反應(yīng)是一個(gè)可逆反應(yīng)。為了使反應(yīng)平衡朝目的產(chǎn)物方向進(jìn)行,可以加大反應(yīng)底物的量或者及時(shí)清除反應(yīng)中的副產(chǎn)物[21]。PLOEGH課題組將分選序列(LPXTG)C端的甘氨酸用甲酯替代,轉(zhuǎn)肽反應(yīng)形成連接產(chǎn)物后,釋放出甲醇而不是甘氨酸親核基團(tuán),阻止逆反應(yīng)進(jìn)行[22],但這種反應(yīng)的產(chǎn)率并不高,而用羥基乙酸替代甘氨酸后的反應(yīng)速率更高[23]。LIU對(duì)甘氨酸親核試劑進(jìn)行改進(jìn),利用酰肼代替甘氨酸作為親核試劑進(jìn)攻酶-底物復(fù)合物形成一種非天然的環(huán)肽產(chǎn)物[24],本課題組利用這種策略合成了非天然的環(huán)肽類似物抗菌肽P-113,產(chǎn)率達(dá)到76%~93%[25](圖1-b)。
1.2 肽連接酶Butelase-1
蝶豆蘭花肽連接酶-1(Bunga telang plus ligase,Butelase-1)是一種天冬酰胺內(nèi)切蛋白酶(asparaginylendoproteases,AEP),參與熱帶藥用植物蝶豆中大環(huán)肽合成[26]。2014年由TAM課題組首次從碟豆中分離出來(lái),能識(shí)別肽鏈C端三肽序列D/N-HV,并從組氨酸前面進(jìn)行肽鍵的斷裂,釋放HV二肽序列并形成酰基酶底物復(fù)合物,隨后的N端氨基酸作為親核基團(tuán)進(jìn)攻復(fù)合物重新形成肽鍵(圖2),產(chǎn)物中連接位點(diǎn)只留下D/N印跡(分選酶A為L(zhǎng)PXTG)[27]。蝶豆蘭花肽連接酶-1對(duì)底物N端第1,2位氨基酸有一定要求,第1位氨基酸可以為除Pro、Asp和Glu以外的任何天然氨基酸,第2位為疏水性氨基酸Ile、Leu、Val以及Cys[28]。

圖2 蝶豆蘭花肽連接酶-1介導(dǎo)環(huán)肽合成
Fig.2 Butelase-1-mediated cyclization of peptides
蝶豆蘭花肽連接酶-1催化效率很高(0.005摩爾當(dāng)量),可以在很短時(shí)間內(nèi)環(huán)化10個(gè)氨基酸或者更長(zhǎng)的環(huán)肽(70個(gè)氨基酸)生成[29-30]。這種酶的催化反應(yīng)水解產(chǎn)物較少,更傾向環(huán)化產(chǎn)物形成,例如,合成29個(gè)氨基酸的環(huán)肽kalata B1產(chǎn)率達(dá)到95%以上,底物的耐受性較好,除C端D/N氨基酸以外,其余的氨基酸都可以為D型氨基酸[28]。
雖然蝶豆蘭花肽連接酶-1催化效率高,但是和分選酶A催化的反應(yīng)一樣,該反應(yīng)也是可逆反應(yīng),反應(yīng)過(guò)程中需要大量底物促使反應(yīng)進(jìn)行。另外,蝶豆蘭花肽連接酶-1目前沒(méi)能實(shí)現(xiàn)基因工程表達(dá),即使添加GST或MBP促溶標(biāo)簽也無(wú)法進(jìn)行原核表達(dá),這限制了蝶豆蘭花肽連接酶-1應(yīng)用前景。HARIIS課題組通過(guò)突變獲得了1種AEP突變體OaAEP1,在進(jìn)化地位上與蝶豆蘭花肽連接酶-1相關(guān),功能類似,可以實(shí)現(xiàn)在大腸桿菌中進(jìn)行表達(dá),但產(chǎn)率較低(<2 mg/L),催化效率也遠(yuǎn)遠(yuǎn)低于蝶豆蘭花肽連接酶-1[31-32]。因此,實(shí)現(xiàn)蝶豆蘭花肽連接酶-1基因工程表達(dá),提高其產(chǎn)量仍是一個(gè)重要課題。
1.3 枯草桿菌蛋白酶(subtillisin)突變體
早在1997年,WELLS課題組利用基因工程技術(shù)從解淀粉芽孢桿菌得到了一種枯草桿菌蛋白酶突變體(S221C,P225A),命名為subtiligase,并初次實(shí)現(xiàn)了該酶的環(huán)肽合成,產(chǎn)率為36%~85%,當(dāng)環(huán)肽大于14個(gè)氨基酸時(shí),主要是環(huán)肽產(chǎn)物,小于14個(gè)氨基酸的產(chǎn)物主要以水解產(chǎn)物和二聚體為主[33]。但該酶的穩(wěn)定性較差,反應(yīng)過(guò)程易生成水解產(chǎn)物。TOPLAK課題組隨后通過(guò)基因工程方法表達(dá)了另1種來(lái)源于枯草芽孢桿菌的枯草桿菌蛋白酶突變體肽連接酶(peptiligase, S221C,P216A),這種蛋白的表達(dá)產(chǎn)率達(dá)到500 mg/L以上,而且酶穩(wěn)定性和催化能力都得到了極大改善[34]。這種突變體連接酶并不依賴于鈣離子,酶的巰基首先與C末端的甲酰胺甲基酯(carboxamidomethyl-ester,oCam)反應(yīng)形成復(fù)合物,N端氨基作為親核試劑進(jìn)攻酶復(fù)合物并形成產(chǎn)物(圖3),這和分選酶A的催化機(jī)理類似[34]。催化形成的環(huán)肽產(chǎn)物與水解產(chǎn)物之比(S/H)與底物肽長(zhǎng)度有關(guān),形成13個(gè)氨基酸產(chǎn)物時(shí)的環(huán)肽產(chǎn)物與水解產(chǎn)物之比(S/H)大于100,線性肽底物1 h內(nèi)的轉(zhuǎn)化率>99%,而轉(zhuǎn)化為水解產(chǎn)物不足1%。并且該反應(yīng)可以允許有機(jī)溶劑(50% DMSO和DMF)和變性劑(2 mol/L尿素或者鹽酸胍)存在,這可以使那些水溶性不好的多肽實(shí)現(xiàn)環(huán)化[34],擴(kuò)大了酶的使用范圍。

圖3 枯草桿菌蛋白酶突變體肽連接酶介導(dǎo)環(huán)肽合成
Fig.3 Peptiligase-mediated cyclization of peptides
枯草桿菌蛋白酶突變體肽連接酶(peptiligase)同樣需要識(shí)別序列,這個(gè)序列包括C末端的4個(gè)氨基酸(P1~P4)以及N末端的2個(gè)氨基酸(P1’和P2’),C末端需要形成活化酯結(jié)構(gòu),P1’氨基酸主要為Ser、Gly和Ala,P1’和P2’不能為Pro,另外,P4位適合疏水性氨基酸(圖3)。而另1種枯草桿菌蛋白酶突變體肽連接酶(omniligase-1)底物寬泛性更好,該酶識(shí)別序列的P1’可以為除了Pro以外的任何氨基酸[35]。而且能夠?qū)崿F(xiàn)最小環(huán)化的氨基酸數(shù)量為11個(gè),當(dāng)環(huán)肽氨基酸數(shù)量大于12個(gè)氨基酸,可以獲得90%產(chǎn)率,并實(shí)現(xiàn)了克級(jí)別環(huán)肽的合成。另外該酶還可以合成復(fù)雜環(huán)肽,比如34個(gè)氨基酸的二硫鍵環(huán)肽MCoTⅠ-Ⅱ,在產(chǎn)物中引入活性位點(diǎn)序列、D型氨基酸以及側(cè)鏈異肽鍵形成都能高效的實(shí)現(xiàn)。
和分選酶A,蝶豆蘭花肽連接酶-1相比,枯草桿菌蛋白酶突變體肽連接酶催化效率更高(<0.0003摩爾當(dāng)量),由于該連接酶并不是轉(zhuǎn)肽酶,是個(gè)非可逆反應(yīng),這樣反應(yīng)水解產(chǎn)物量很低,但需要在底物肽C末端引入活化酯結(jié)構(gòu)。
1.4 硫酯酶(thioesterase)
自然界中生物合成的環(huán)肽可以分為核糖體環(huán)肽和非核糖體環(huán)肽,非核糖體環(huán)肽的合成是由非核糖體肽合成酶(nonribosomal peptide synthetases,NRPSS)完成,這種酶是目前發(fā)現(xiàn)的最大的酶復(fù)合物,由于沒(méi)有核糖體幫助,非核糖體環(huán)肽合成依賴于這種大的酶復(fù)合物完成氨基酸運(yùn)載、肽鍵形成、多肽轉(zhuǎn)移與釋放等多種功能[36]。酶復(fù)合物中最為關(guān)鍵的結(jié)構(gòu)域?yàn)榱蝓ッ附Y(jié)構(gòu)域(thioesterase domain,TE-domain),該結(jié)構(gòu)域具有環(huán)化、終止反應(yīng)和釋放產(chǎn)物功能,含有保守催化三肽序列(SHD),其中Ser側(cè)鏈羥基進(jìn)攻底物肽C端硫酯結(jié)構(gòu)(常用的有SNAC)中的羰基形成肽-酰中間產(chǎn)物,中間產(chǎn)物可以被水解成線性肽,也可以被分子內(nèi)的親核基團(tuán)(如氨基或者羥基)攻擊形成環(huán)化產(chǎn)物(圖4),親核基團(tuán)可以來(lái)源于N端(如氨基)或者側(cè)鏈[37]。
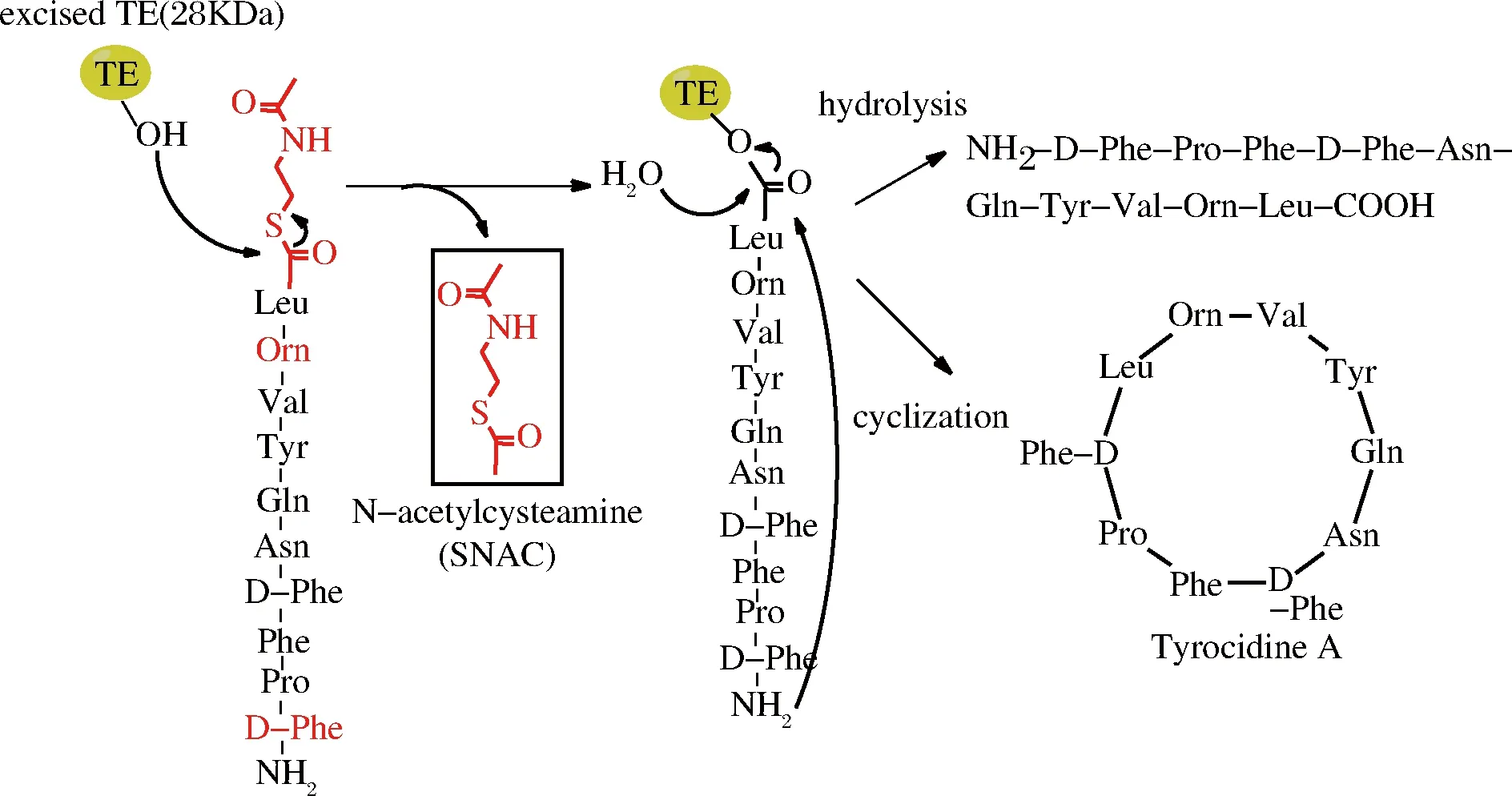
圖4 TE結(jié)構(gòu)域介導(dǎo)的環(huán)肽合成(以合成短桿菌酪肽為例)
Fig.4 TE-mediated cyclization of peptides
早在2000年,WALSH課題組從短桿菌Bacillusbrevis中克隆了合成短桿菌酪肽TE結(jié)構(gòu)域TycC TE(tyrocidine TE,28 kDa),在體外成功合成了環(huán)肽短桿菌肽S和短桿菌酪肽[36],隨后他們又分別從Bacillusbrevis,Bacillussubtilis中克隆了合成短桿菌肽和脂肽的TE結(jié)構(gòu)域GrsB TE和SrfA TE,都能實(shí)現(xiàn)體外合成相應(yīng)環(huán)肽[38]。TRAUGER等探索了TycC TE對(duì)底物的專一性,N-端的D-Phe以及序列中9位稀有氨基酸Orn對(duì)環(huán)化影響較大,鳥(niǎo)氨酸能夠使底物肽形成β結(jié)構(gòu),這樣易于與酶發(fā)生相互作用,因此,線性肽N端和C端氨基酸種類以及底物的二級(jí)結(jié)構(gòu)對(duì)環(huán)肽合成具有重要作用[36]。
為了拓展TycC TE在合成環(huán)肽方面的應(yīng)用,WALSH課題組將固相合成與酶法催化結(jié)合起來(lái),利用PEGA樹(shù)脂作為載體,在96孔裝置里一步法進(jìn)行多肽固相合成與酶法環(huán)化,合成了短桿菌酪肽庫(kù)用于篩選活性更好的抗菌肽,環(huán)化產(chǎn)物與線性水解肽比例約為0~48.1,由于水解產(chǎn)物,最終環(huán)化產(chǎn)率較低[39]。WU等利用同樣策略合成短桿菌肽S,固相載體改為TentaGel樹(shù)脂,TE結(jié)構(gòu)域?yàn)镚rsB TE,但樹(shù)脂上只有28%的線性肽進(jìn)行了環(huán)化,即使增加反應(yīng)溫度以及酶量,都無(wú)法提高轉(zhuǎn)化率[40],這可能與酶沒(méi)法進(jìn)入固相載體與底物肽進(jìn)行充分的接觸有關(guān),需要對(duì)酶的大小尺寸進(jìn)行基因工程改進(jìn),或者改變固相載體。
硫酯酶突變體在體外合成環(huán)肽主要模擬生物體內(nèi)合成抗菌肽的機(jī)理,特定的TE結(jié)構(gòu)域往往只能合成特定的環(huán)肽及其類似物(如合成短桿菌肽、短桿菌酪肽和脂環(huán)肽及其類似物),對(duì)其他序列的適應(yīng)性并不清楚,序列中需要含有大量的非天然氨基酸和稀有氨基酸以形成酶的最適底物構(gòu)象,難以推廣至其他環(huán)肽合成。目前為止,大量的水解產(chǎn)物問(wèn)題并沒(méi)有得到很好的解決。
1.5 PatG相關(guān)結(jié)構(gòu)域
海洋藍(lán)藻環(huán)肽(cyanobactins)是由海洋生物體內(nèi)合成的核糖體環(huán)肽,具有抗腫瘤活性。合成此類環(huán)肽也是由一系列的酶協(xié)調(diào)完成,2009年,SCHMIDT等從珊瑚礁中克隆了合成該海洋藍(lán)藻環(huán)肽的環(huán)化功能的結(jié)構(gòu)域(PatG),并在體外合成了該環(huán)肽及其類似物[41]。這種酶催化環(huán)肽合成機(jī)理類似于硫酯酶,也會(huì)形成水解線性肽副產(chǎn)物,區(qū)別在于它并不需要引入活化酯結(jié)構(gòu),因此,底物線性肽可以直接利用固相合成技術(shù)制備[42]。NAISMITH課題組隨后從Prochloronsp. 中克隆了合成小盤酰氨(patellamide)的環(huán)化結(jié)構(gòu)域(PatGmac)[43],這類酶首先識(shí)別底物肽C端信號(hào)序列AYG(E),酶上的羥基進(jìn)攻A前面的肽鍵并形成酰基酶復(fù)合物,同時(shí)釋放AYG(E)短肽,N端的氨基作為親電基團(tuán)進(jìn)攻酯結(jié)構(gòu)中的羰基完成環(huán)化[44]。AYG(E)中丙氨酸前面(P1位)的氨基酸對(duì)環(huán)肽合成至關(guān)重要,一般都是脯氨酸或者噻唑啉結(jié)構(gòu)(如圖5中Tzl),這種結(jié)構(gòu)能夠使酶與底物形成最佳結(jié)合構(gòu)象[45]。另一種從海洋藍(lán)藻中發(fā)現(xiàn)的OscGmac對(duì)P1位的寬泛性更好,利用Ser和Cys模擬的脯氨酸結(jié)構(gòu)都可以作為環(huán)化底物,包括底物中引入D型氨基酸,對(duì)6~11個(gè)氨基酸長(zhǎng)度的底物環(huán)化產(chǎn)率較好[46]。這種酶和硫酯酶一樣,都是從生物體內(nèi)合成特定環(huán)肽的酶復(fù)合體中分離得到的環(huán)化結(jié)構(gòu)域,對(duì)底物序列是有一定要求,大范圍的改變序列會(huì)導(dǎo)致環(huán)化失敗,而且正常環(huán)化產(chǎn)率并不高(32%~58%)[47]。

圖5 PatG介導(dǎo)環(huán)肽合成(以合成patellamide C為例)
Fig.5 PatG-mediated cyclization of peptides
1.6 谷胱甘肽S-轉(zhuǎn)移酶(glutathione S-transferase,GST)
PENTELUTE課題組首次報(bào)道了利用GST偶聯(lián)原理來(lái)合成大環(huán)肽,這種環(huán)化方式主要是側(cè)鏈之間的連接。他們將多肽N端接入GST識(shí)別標(biāo)簽ECG三肽序列,C端通過(guò)化學(xué)合成引入氟苯親電試劑,在GST作用下氟苯與ECG中的半胱氨酸巰基進(jìn)行共價(jià)連接(圖6)[48]。對(duì)比不同氟苯親電試劑發(fā)現(xiàn)十氟聯(lián)苯效果最好,反應(yīng)速度更快,5 min能夠完成90%以上的環(huán)化。環(huán)肽鏈在一定長(zhǎng)度范圍內(nèi)(14~24個(gè)氨基酸),隨著肽鏈增長(zhǎng),環(huán)化產(chǎn)率也相應(yīng)增大(93%~96%),肽鏈為40個(gè)氨基酸時(shí),可以獲得70%產(chǎn)率。對(duì)于其他的轉(zhuǎn)肽酶,底物肽濃度增加會(huì)導(dǎo)致多聚體副產(chǎn)物生成,而這種環(huán)化當(dāng)?shù)孜镫臐舛冗_(dá)到10 mmol/L,而酶濃度只需要摩爾分?jǐn)?shù)為0.1時(shí),就檢測(cè)不到多聚體副產(chǎn)物。另外,GST適應(yīng)的溫度范圍比較廣(4~60 ℃),水相中允許添加有機(jī)溶劑可以達(dá)到20%。但和NCL方法類似,這種環(huán)化方法仍需要引入巰基以及活性較好的親電基團(tuán)[48]。
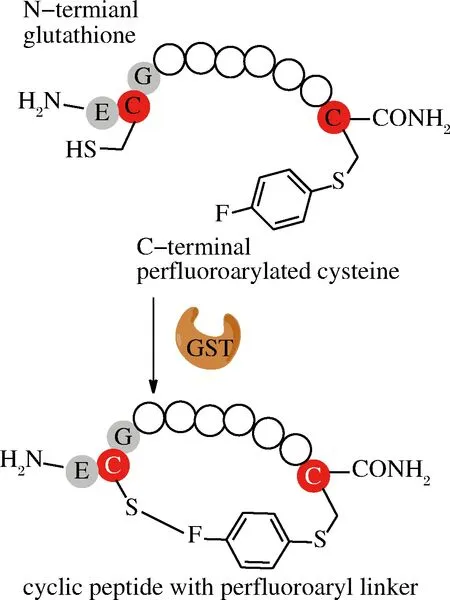
圖6 GST介導(dǎo)環(huán)肽合成
Fig.6 GST-mediated cyclization of peptides
1.7 谷氨酰胺轉(zhuǎn)氨酶(transglutaminase,TGase)
在自然界中,TGase主要催化蛋白質(zhì)或者多肽中谷氨酰胺上的酰胺與其他伯胺之間的酰基轉(zhuǎn)移反應(yīng),參與蛋白質(zhì)與蛋白質(zhì)或者小分子之間的交聯(lián)形成異肽鍵。在食品工業(yè)上,TGase經(jīng)常用來(lái)進(jìn)行肉質(zhì)酥松,也可以對(duì)大分子進(jìn)行小分子偶聯(lián)修飾,例如,將親和素、熒光基團(tuán)等偶聯(lián)至抗體上[49]。但是利用TGase進(jìn)化環(huán)肽合成報(bào)道較少。
TOUATI等利用來(lái)源于微生物鏈霉菌(Streptomycesmobaraensis)的谷氨酰胺轉(zhuǎn)氨酶(MTGase,38 kDa)嘗試了血漿激肽釋放酶抑制二環(huán)肽合成(圖7)[50]。
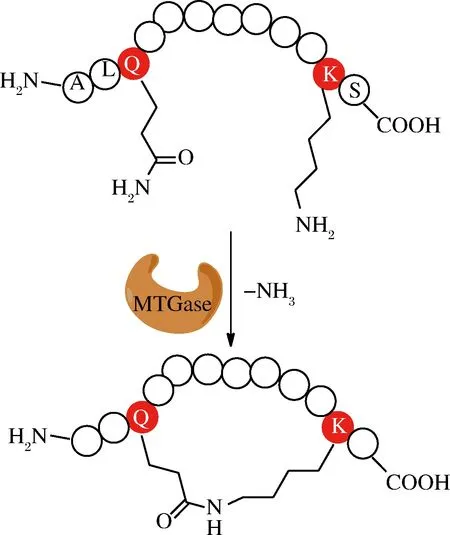
圖7 MTGase介導(dǎo)的環(huán)肽合成
Fig.7 MTGase -mediated cyclization of peptides
N端設(shè)計(jì)成酶的識(shí)別序列WALQRPH,其中Q作為酰基供體,C端引入了KS序列,K作為酰基受體,中間使用多聚甘氨酸鏈作為連接。這種識(shí)別序列只能置于N端,易于酰基供體與受體在空間上靠近。信號(hào)序列截短實(shí)驗(yàn)發(fā)現(xiàn)酰基供體Q前面的2個(gè)氨基酸對(duì)環(huán)化非常重要,在底物濃度較高情況下,同樣會(huì)出現(xiàn)二聚體副產(chǎn)物。
2 結(jié)論與展望
基于環(huán)肽復(fù)雜的拓?fù)浣Y(jié)構(gòu)和獨(dú)特的生物活性,環(huán)肽逐漸成為研究熱點(diǎn),有可能解決某些大蛋白無(wú)法成藥的困境。酶法和多肽固相合成技術(shù)相結(jié)合可以合成結(jié)構(gòu)復(fù)雜、種類多樣的環(huán)肽結(jié)構(gòu),為藥物篩選提供平臺(tái)。為了降低酶法合成成本,實(shí)現(xiàn)工業(yè)化級(jí)別合成,酶的獲取成為關(guān)鍵因素,目前,有些酶實(shí)現(xiàn)了商品化,例如,枯草桿菌蛋白酶突變體肽連接酶(omniligase-1)已經(jīng)開(kāi)發(fā)為試劑盒,可以進(jìn)行克級(jí)別的環(huán)肽制備,即使有些來(lái)源于植物的環(huán)肽合成酶暫時(shí)還沒(méi)法實(shí)現(xiàn)基因工程表達(dá),但相信這些問(wèn)題都會(huì)得到解決。另外,生物體內(nèi)非核糖體環(huán)肽合成酶可以合成多樣化的環(huán)肽,這為研究者提供了一個(gè)環(huán)化酶的篩選平臺(tái),利用分子生物學(xué)技術(shù)篩選定制酶也會(huì)成為一個(gè)新的研究方向。借助于計(jì)算機(jī)工具以及多肽發(fā)現(xiàn)平臺(tái),酶法合成可以作為化學(xué)合成策略很好的補(bǔ)充,加速未來(lái)環(huán)肽療法的發(fā)展。
