嫁接體植株中核酸與蛋白質的砧穗交流
2019-01-25 01:33:04謝露露尚慶茂
西北農業學報 2019年1期
關鍵詞:長距離
謝露露,尚慶茂
(中國農業科學院 蔬菜花卉研究所,北京 100081)
嫁接指將一個植株從莖或下胚軸處切斷獲得的芽或枝,與另一個植株帶有根系的部分于切口處貼接,使其愈合并生長成新的嫁接體植株。芽或枝的部分稱為接穗(scion),帶有根系的部分稱為砧木(stock),接穗和砧木愈合的部位稱為嫁接結合部(graft union)。嫁接結合部具有連通砧木與接穗的維管組織,保證嫁接體植株砧穗兩部分的共同生長和發育。或許受到自然界中不同植物個體之間融合生長現象的啟迪,在公元2000多年前,我們民眾就已經能夠使用嫁接技術和表型篩選,獲得具有更優良性狀的作物[1]。現如今,嫁接已被作為一種常規手段廣泛應用于蔬菜的繁育。較之蔬菜品種單獨種植,優良的嫁接體植株具有產量和品質提高、生物或非生物脅迫抗性增強、株型優化、開花或塊莖化習性優化等增益效果[2-4]。然而并非任意品種之間的嫁接均能獲得理想的性狀改良。在生產應用中,經常會發生一個性狀獲得改良的同時另一性狀退化,或是改良效果不明顯等現象。對優良砧穗組合的篩選還存在很大的盲目性。為了探討表型變化的規律,砧穗間物質交流的類型和方式被廣泛研究。之前的觀點認為,只有糖類、無機鹽離子、小分子代謝物和內源性植物激素可經由韌皮部在植物體內被系統性地轉運,而近10余年的研究證實,大分子物質如蛋白質和RNA也可在細胞間轉運和韌皮部運輸[5-6]。隨后又發現甚至DNA也可以在嫁接面局部發生轉移[7]。DNA、RNA、蛋白質等大分子物質在砧穗間的交流,很可能對性狀改變起到更為關鍵的作用。本文結合最新進展,按照物質類型和砧穗間交流方式進行歸納和梳理(表1)。
1 DNA在嫁接面局部的水平轉移
近期研究表明,砧穗之間存在基因水平轉移(Horizontal gene transfer,HGT)現象。Stegemann及其合作者將核基因組和葉綠體基因組分別標記抗性基因和熒光標識的2個煙草轉基因株系以斜切45°的方式嫁接,待嫁接體形成后在嫁接結合部連續橫切,再將由此獲得的莖組織薄片在含有2種抗生素選擇壓力的培養皿上培養。培養皿上分化出來的植株兼具2種抗性基因和2種熒光標識,而遠離嫁接部位的莖組織薄片則不能分化出植株,這個現象說明了砧穗嫁接面附近區域發生的基因水平轉移事件[7]。隨后他們又將標記葉綠體基因組的煙草(Nicotianatabacum)作為砧木,標記核基因組的煙草屬另外2個種N.glauca和N.benthamiana作為接穗,同樣是利用嫁接結合部切片和雙抗選擇壓下的組織培養獲得抗性植株。結果顯示,該植株的表型在幼苗期和成體期均與接穗物種的表型幾乎相同。DNA分子標記和特征片段測序結果表明,雙抗培養基篩選出來的植株捕獲了來自砧木的完整葉綠體基因組[8]。為了繼續驗證核基因組是否也有轉移,將N.tabacum(核型2n=48)2個株系的核基因組分別標記,獲得了包含2套核基因組標記的核基因轉移(Nuclear gene transfer,NGT)株系,該NGT株系可以自交形成種子,自交后代染色體數目多為96,且表型變異很大。有趣的是,如果將N.tabacum與染色體數目不同的N.glauca(核型2n=24)嫁接獲得NGT株系后,則自交后代的染色體數目穩定在72條,且群體內表型一致,抗性基因呈現孟德爾分離比。嫁接部位由疑似細胞融合(Cytomixis)機制產生的異源多倍體具有無繁殖障礙和穩定遺傳等特點,表明經由基因水平轉移介導的非有性(Asexual)途徑能夠形成嫁接雜合體(Graft hybrid)的事實[9]。
由以上試驗可知,砧穗間的DNA交流形式既包括完整葉綠體基因組,又包括核基因組。然而交流僅發生在嫁接面部位,即使有從嫁接結合部小概率長成的具有嫁接雜合體性質的芽或側枝,嫁接體植株上絕大部分組織仍然不具備從嫁接搭檔那里捕獲的DNA。盡管如此,卻不能完全排除DNA轉移對表型的影響。研究表明,基因水平轉移普遍存在于陸地植物的表型演化歷程,植物基因組中保留的異源基因在一些基本功能中發揮作用,如木質部形成、植物防御、氮循環以及淀粉、多胺、植物激素和谷胱甘肽等的生物合成等[10]。而嫁接結合部作為砧木和接穗在形態和生理上的緩沖區,DNA所承載遺傳信息的局部轉移會對砧穗雙方的表型產生怎樣的影響,還需要進一步證實。
2 蛋白質在砧穗間的長距離運輸
遍布植物體的韌皮部篩管是由首尾相接的篩分子(Sieve elements,SE)-伴胞(Companion cell,CC)復合體構成。篩管中的韌皮部運輸流(Phloem translocation stream),是物質在植物體內各個組織間長距離系統性運輸(Long-distance systemic transportation)的唯一途徑。韌皮部的SE和CC連接處,有特化的孔間單元(Pore plasmodesmal units,PPUs)結構。由于這種結構的存在,胞間連絲的尺寸排阻極限(Size exclusion limit,SEL)可達到遠超普通細胞間SEL的程度,為1 ku以上,最大為67 ku[11]。這樣的SEL足夠蛋白質和分子質量較小的RNA通過[5]。可以在韌皮部運輸的物質,當砧穗正常愈合后,均可以在砧穗間穿梭。利用嫁接系統完成的長距離運輸試驗,為大分子物質的砧穗間穿梭提供了大量證據。
從韌皮部汁液(Phloem sap)中鑒定出來的蛋白質數量眾多[11]。綠色熒光蛋白(Green fluorescent protein,GFP)和所有分子質量低于SEL閾值的GFP融合蛋白均能夠通過CC-SE邊界[5,12]。說明蛋白質在進入韌皮部時的方式是非選擇性的。另外有證據表明,較之內質網錨定核糖體上翻譯出的蛋白質,胞質游離核糖體上翻譯出來的蛋白質更容易進入SE。而編碼亞細胞定位信號的多肽鏈不足以指導蛋白質的韌皮部運輸。這說明,韌皮部蛋白質中很大一部分是由于非特異性流失而造成的[13]。
韌皮部運輸流中的蛋白質至少具有2種功能形式:信號傳遞和輔助運輸。起信號傳遞作用的蛋白在植物體內的長距離運輸,履行系統性防御或者發育信號傳遞的功能。例如成花素(Florigen)被認為是光信號接收部位和莖頂端開花效應部位之間的信號傳遞分子。在南瓜屬(Cucubita)2個種的異源嫁接試驗中,成花素的身份被確定為FT(Flowering locus T)蛋白。將長日照條件下生長的短日照植物白籽南瓜(C.moschata)與已經開花的印度南瓜(C.maxima)嫁接,則白籽南瓜接穗被誘導開花,伴隨接穗韌皮部汁液中大量存在的Cm-FTL1(Flowering locus T-like1)和Cm-FTL2蛋白,而非它們的mRNA[14]。FT蛋白不僅是誘導開花的信號蛋白,還能夠誘導馬鈴薯(Solanumtuberosum)地下莖的塊莖化(Tuberization)。如果將水稻的FT同源基因Hd3a在馬鈴薯中表達,并將轉基因植株作為接穗與野生型嫁接,則能夠誘導原本在長日照條件下生長不形成塊莖的野生型砧木形成塊莖[15]。另外,親環蛋白(Cyclophilin)也代表了一類可長距離運輸的蛋白質信號分子。在番茄(Solanumlycopesicum)中,將野生型接穗與親環蛋白SlCyp1突變的砧木嫁接后,可以恢復突變體的根系發育和對生長素的敏感性[16]。
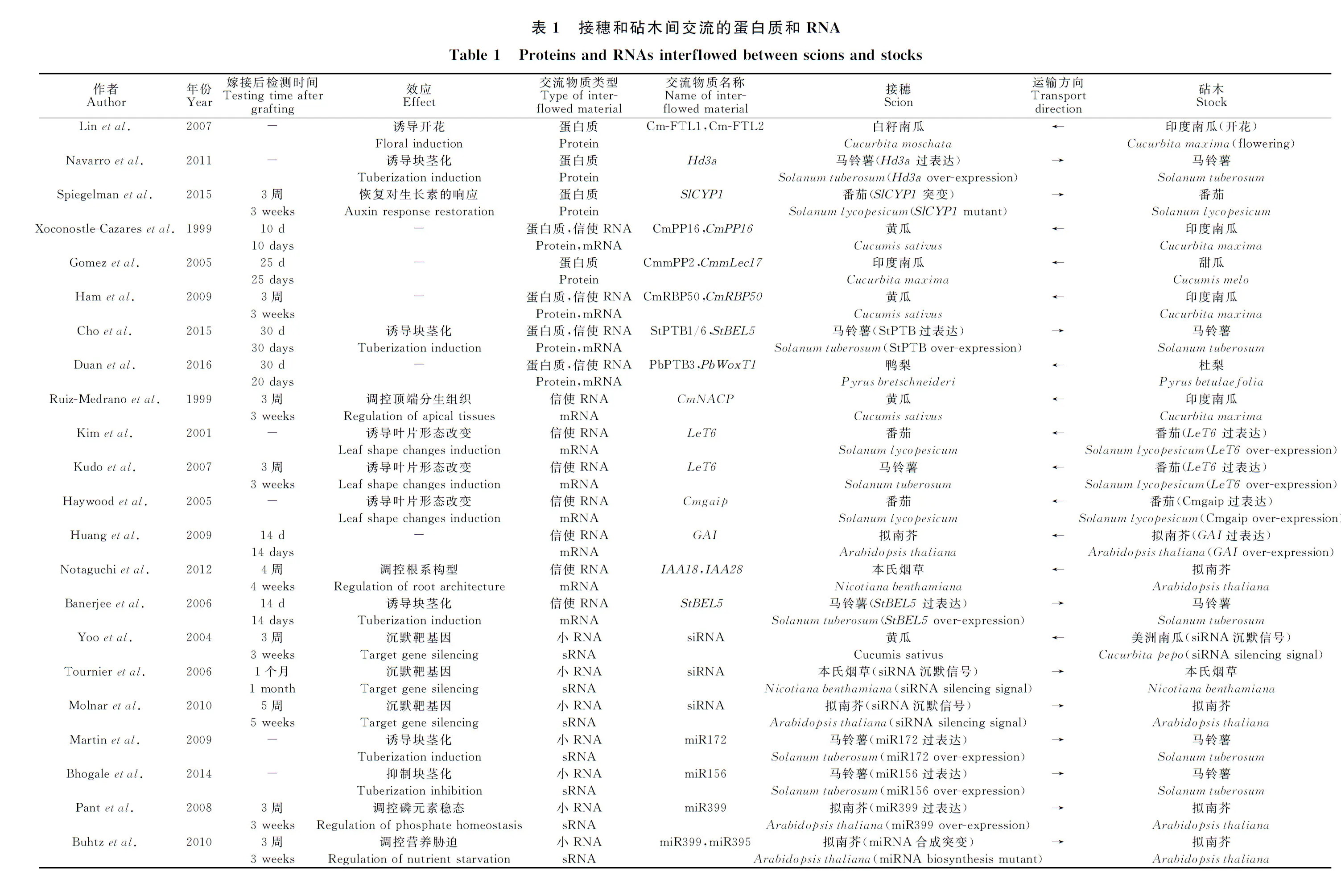
在韌皮部中還鑒定到一些行駛輔助運輸功能的蛋白質,被認為是大分子物質系統性長距離運輸機制的組成部分。這類蛋白結合RNA分子,同時能夠增大胞間連絲(Plasmodesmata,PD)的CEL,從而輔助RNA的選擇性運輸[5]。如印度南瓜中的CmPP16(Phloem protein 16),與病毒移動蛋白(Movement proteins,MPs)同源,存在于SE中,能結合包括自身mRNA在內的若干種RNA分子[17]。編碼細胞色素b5還原酶(Cytochrome b5 reductase,Cb5R)的CmPP36也結合RNA,但在韌皮部汁液中鑒定到的CmPP36蛋白,是其完整編碼序列去除N端一段跨膜序列后的形態,人為去除N端序列后,可以實現CmPP36蛋白的韌皮部轉運,從而推測蛋白質進入韌皮部時可能存在蛋白質水解過程[18]。此外,韌皮部凝集素(Lectin)類蛋白CmmPP2和CmmLec17在CC中合成,經PD轉運至SE,能夠結合韌皮部RNA和病毒RNA。甜瓜(Cucumismelo)砧木和印度南瓜接穗嫁接后,甜瓜CmmLec17蛋白在嫁接后25 d在印度南瓜接穗中被檢測到[19]。另一類RNA結合蛋白,PTB(Polypyrimidine tract-binding)類蛋白傾向于結合在RNA的尿嘧啶(U)和胞嘧啶(C)富集區域,介導RNA的運輸,如印度南瓜CmRBP50(RNA binding protein 50)蛋白和自身轉錄本的運輸[20]、馬鈴薯中的StPTB1/6 介導StBEL5(BEL1-like 5)轉錄本的運輸[21]、杜梨(Pyrusbetulaefolia)中的PbPTB3蛋白介導PbWoxT1(Wox transport 1)轉錄本的運輸[22]。
3 信使RNA在砧穗間的長距離運輸
在韌皮部汁液中發現可遷移的mRNA(Transmissible mRNAs),曾讓人們感到出乎意料[6]。通過葫蘆科和茄科物種的異源嫁接試驗,發現多個可遷移mRNA,如CmNACP(NAC domain protein)從南瓜砧木遷移至黃瓜(Cucumissativus)接穗,參與黃瓜頂端分生組織的發育調控[6]。從番茄的Me(Mouseear)自然突變體中遷移出的過量LeT6(KNOTTED-1-like homeobox)基因轉錄本導致番茄和馬鈴薯接穗的葉片形態發生明顯改變[23-24]。擬南芥(Arabidopsisthaliana)gai-1(gibberellicacidinsensitive1)突變體中的GAI基因缺失了DELLA結構域序列,因而存在GA信號途徑的異常。將帶有ΔDELLA-gai基因的擬南芥或者遺傳轉化Cmgaip基因的番茄作為砧木,與各自的野生型植株進行嫁接,均導致野生型接穗呈現植株矮小的赤霉素缺陷表型,gai轉錄本的長距離運輸是其內在原因[25]。在擬南芥和本氏煙草(Nicotianabenthamiana)的遠緣嫁接試驗中,擬南芥生長素響應蛋白(Indoleacetic acid-inducible protein)IAA18/28轉錄本在本氏煙草接穗中被檢測到。由于此類蛋白定位于根系,被認為與根系形態的發育調控有關[26]。以上均是mRNA從砧木運輸至接穗的例證,關于塊莖化誘導的試驗表明,mRNA同樣可以由接穗運輸至砧木。例如在馬鈴薯中過表達StBEL5(BEL1-like 5)的接穗能夠誘導砧木的塊莖形成[27]。
除了已經被驗證功能的可遷移mRNA,最近幾項有關擬南芥、葡萄(Vitisvinifera)、葫蘆科異源嫁接系統的研究,利用高通量測序技術從組學角度闡釋了mRNA的遷移[28-30]。將擬南芥2種生態型互相嫁接,以單核苷酸位點多態性(Single nucleotide polymorphism,SNP)為標簽來區分不同來源的轉錄本,篩選出數以千計的可遷移mRNA。可遷移mRNA普遍具有明確的遷移方向和目標組織[28]。蛋白質組還顯示,可遷移mRNA是在運至目標組織之后才被翻譯為蛋白質[28]。使用黃瓜與西瓜(Citrulluslanatus)的異源嫁接體系,并加以營養脅迫處理后,可遷移mRNA在嫁接搭檔不同組織間的重復性很低,再次說明源組織中的mRNA被有針對性地運往特定的庫組織[30]。可遷移mRNA的類群和遷移方式與嫁接體所處的發育階段和外部環境密不可分。比如成體期中的可遷移mRNA數量比幼體期中的少得多[29]。同是營養缺乏脅迫,缺氮和缺磷時鑒定到的可遷移mRNA的種類和數量較之全營養條件明顯不同[28]。磷短期缺乏處理(24 h)后,可遷移mRNA中富集有已知直接參與磷信號轉導和同化過程的轉錄本,如VPT(Vacuolar phosphate transporter 1)、PHO2(Phosphate 2)、CAX3(Cation exchanger 3)等,以及脅迫反應通用的一些編碼各類激素受體和信號通路的轉錄本[30]。
mRNA的運輸除了需要起輔助運輸作用的蛋白質,還與mRNA自身的序列和結構密切相關。在擬南芥中,僅是GFP的轉錄本不能進入韌皮部,而當GFP與GAI的序列融合時,則能被長距離運輸。推測這與GAI上帶有的特征性基序(motif)和二級結構有關[31]。在馬鈴薯中,韌皮部遷移轉錄本StBEL5的3′UTR區域對維持mRNA的穩定性和向根運輸是必要的[32]。進一步研究表明,較高比例的可遷移mRNA在編碼區和UTR區具有類似tRNA的莖環結構(tRNA-like structure,TLS),這種TLS結構負責驅動mRNA的遷移[33]。另外,RNA序列上多聚UC(Uracil cytosine)元件的存在有利于PTB類RNA結合蛋白的結合[21]。
4 小RNA在砧穗間的長距離運輸
在mRNA被證實經韌皮部運輸之后,小RNA群體的長距離運輸也被證實。從韌皮部汁液中檢測到的小RNA(sRNA)包括小干擾RNA(Short interfering RNA,siRNA)和微小RNA(microRNA,miRNA)[34]。siRNA和miRNA是由雙鏈RNA經Dicer酶家族成員切割而形成的長度為21~26 nt單鏈RNA[35-36]。砧穗完全愈合后sRNA可經由韌皮部在嫁接結合部上下自由移動。可移動的sRNA以結合并裂解mRNA的形式使嫁接搭檔的基因被轉錄后沉默(Post-transcriptional gene silencing,PTGS),或是以RNA介導的DNA甲基化(RNA-directed DNA methylation,RDDM)途徑改變DNA的表觀遺傳修飾,使基因被轉錄沉默(Transcriptional gene silencing,TGS)[37]。
轉基因美洲南瓜(Cucubitapepo)作為砧木,野生型黃瓜作為接穗的嫁接中,砧木中的CP(coatprotein)基因特異性siRNA能夠通過長距離運輸沉默接穗中的CP轉錄本[34]。將分別具有GFP特異性siRNA沉默信號和非沉默的2個GFP轉基因煙草株系正反嫁接,結合GFP熒光和韌皮部同位素示蹤,顯示出siRNA介導的沉默信號跟隨韌皮部運輸流從源(Source)組織被穩定運往庫(Sink)組織[38]。利用擬南芥的嫁接試驗證實,24 nt siRNA可介導受體細胞中全基因組的DNA甲基化[39-40]。由于長距離運輸的siRNA同樣會出現在繁殖器官的頂端生長點或花粉細胞中,誘導DNA表觀遺傳層面的改變,因此有可能將嫁接體的性狀遺傳至后代個體[41]。
miRNA通常以負調控因子身份作用于轉錄因子,參與發育進程或環境響應的調控。例如,miR156和miR172的系統性運輸調控植物從幼體到成體的發育轉換。在馬鈴薯中,過表達miR172的植株表現出花期提前和塊根形成提前,miR172過表達接穗與野生型砧木雜交,能夠誘導野生型砧木的塊根形成量增多[42]。過表達miR156的植株表現為植株構型改變和塊根產量降低,miR156過表達接穗和野生型砧木嫁接后,砧木的葉片形態發生改變[43]。油菜(Brassicanapus)、印度南瓜中的研究結果顯示,當植株面臨磷缺乏脅迫時,韌皮部汁液中miR399大量積累,通過轉錄后沉默機制抑制磷吸收負調因子PHO2(Phosphate 2),提高植物對磷的吸收[44-45]。擬南芥微嫁接試驗顯示miR399過表達接穗能夠誘導野生型根系中產生大量成熟態miR399,如果砧穗互換,則在接穗中檢測不到或僅檢測出痕量的成熟態miR399[45]。在受到硫缺乏脅迫時的油菜韌皮部汁液中,miR395和miR171均明顯上調[46]。miR399和miR395在植物面臨營養脅迫時,從接穗傳遞至砧木,而不是相反方向,miR171則不移動[46]。從而表明miRNA不僅可以在砧穗間長距離運輸,改變嫁接搭檔的表型和生理狀況,而且移動具有明顯的方向性。
即使sRNA的分子量很小,它們在細胞間并不能自由擴散,而是需要特異性sRNA結合蛋白(Small RNA binding protein,SRPB)的輔助[34]。sRNA在源組織中合成后,被運往庫組織的運輸形式可以是初級形態,也可以是成熟形態[40]。
5 小 結
綜上所述,DNA的交換局限于嫁接面,目前沒有證據表明它們對接穗或砧木的整體表型產生影響;蛋白質的運輸則更多起到輔助作用,即使作為信號分子存在,也如同小分子代謝物或內源性植物激素的作用,并不攜帶遺傳信息。而RNA在砧穗間的互作是非常值得關注的。mRNA的大規模系統性運輸,以及sRNA的遠程調控和表觀遺傳修飾作用,對嫁接體植株的表型變異影響顯著。大分子物質在韌皮部運輸流中的長距離運輸,尤其是RNA的運輸,及其所引起的表觀遺傳層面的變化,應該受到更加廣泛的關注。期待該領域后續的研究和進展,為砧穗組合效果和嫁接體生長發育的精準預測開辟捷徑,從而深入挖掘嫁接技術的潛力,使之更好地促進農業發展和農民收益。
猜你喜歡
家庭影院技術(2019年11期)2019-12-09 09:14:14
建材發展導向(2019年10期)2019-08-24 06:26:42
水利規劃與設計(2017年8期)2017-12-20 08:24:07
水利規劃與設計(2017年8期)2017-12-20 08:24:02
水利規劃與設計(2017年6期)2017-07-18 10:56:26
水利規劃與設計(2017年12期)2017-02-06 03:39:50
家庭影院技術(2017年12期)2017-02-06 02:32:16
焊接(2015年9期)2015-07-18 11:03:52
水利建設與管理(2015年10期)2015-05-09 08:29:47
現代企業(2015年6期)2015-02-28 18:51:53
