酵母菌發酵對常用飼料原料營養指標和抗營養因子含量的影響
2019-01-03 02:05:58謝鵬袁園葛瑩鐘佳祎蔡羽娟
江蘇農業科學 2019年21期
謝鵬 袁園 葛瑩 鐘佳祎 蔡羽娟


摘要:生物發酵飼料是目前動物營養研究的熱點領域,具有廣闊的應用前景。比較研究了5種常用飼料原料(小麥、玉米、豌豆、大豆和豆粕)經釀酒酵母、熱帶假絲酵母和馬克斯克魯維酵母發酵后營養物質和抗營養因子含量的變化規律。結果表明,小麥和玉米經3種酵母發酵后pH值顯著降低。豌豆、豆粕和大豆在發酵后期pH值均升高。除豆粕經熱帶假絲酵母發酵后干物質回收率上升外,其他原料經酵母菌發酵后干物質量均降低。玉米原料分別經釀酒酵母和馬克斯克魯維酵母發酵后還原糖含量顯著下降,而大豆、豌豆和豆粕原料經2種酵母發酵后還原糖含量均表現出先升高后降低的規律。熱帶假絲酵母發酵小麥、玉米、大豆和豆粕4種原料粗蛋白含量均顯著升高。3種酵母菌能夠有效降低飼料原料中的植酸含量,但僅對小麥和玉米中的抗性淀粉有一定的降解效果。綜上所述,通過控制酵母菌發酵時間,能夠有效提高飼料原料的營養價值并降低其中的抗營養因子含量。利用酵母菌對飼料原料進行預發酵處理成為提高飼料中營養物質消化吸收利用率的有效途徑。
關鍵詞:釀酒酵母;熱帶假絲酵母;馬克斯克魯維酵母;發酵;飼料;營養指標;抗營養因子
中圖分類號: S816.6文獻標志碼: A
文章編號:1002-1302(2019)21-0241-06
收稿日期:2018-08-02
基金項目:江蘇省淮安市科技計劃項目——重點研發計劃(現代農業)(編號:HAN201605)。
作者簡介:謝鵬(1986—),男,江蘇淮安人,博士,副教授,主要從事動物營養與生理學研究。E-mail:pengxiejqs@126.com。
隨著全球經濟的蓬勃發展和人們生活水平的不斷提高,人類對于畜產品的需求量逐年增大,從而促使畜牧業在規模與質量上不斷發展。在全球人口日益增多的背景下,如何利用生物技術方法開發新型飼料資源并提高動物對原有飼料原料利用率成為解決不斷增長的畜產品需求量和日益緊縮的耕地面積之間矛盾的有效途徑之一。
研究表明,飼料經益生菌發酵后,菌種產生的多種代謝產物不僅能夠提高飼料原料中蛋白質、脂肪和糖類等成分的營養轉化率,同時能夠降解常用飼料原料中的抗營養因子等有害成分,擴大飼料原料的使用范圍,提高原有飼料的營養價值[1-3]。另外,發酵飼料能夠改善飼料適口性,調節動物腸道的微生態平衡,提高了畜禽產品的食品安全性[4-5]。目前,應用于微生物發酵飼料的菌種類型主要有酵母菌、乳酸菌、芽孢桿菌、曲霉等。其中,酵母菌是單細胞真核微生物的一種,能夠發酵糖類物質,其本身含有豐富的氨基酸、維生素、酶類等物質,是直接食用最多的一種微生物,在畜牧業生產中已有廣泛的應用。本研究分別采用釀酒酵母、熱帶假絲酵母和馬克思克魯維酵母對5種常用飼料原料小麥、玉米、豌豆、大豆和豆粕進行發酵處理,對發酵前后的營養指標和抗營養因子含量進行比較測定,為發酵原料在飼料工業中的應用提供理論基礎和技術支撐。
1材料與方法
1.1試驗材料
1.1.1菌種
熱帶假絲酵母(Candida tropicalis)和釀酒酵母(Saccharomyces cerevisiae)均購自中國工業微生物菌種保藏管理中心。馬克思克魯維酵母(Kluyveromyces marxianus)由淮陰師范學院生命科學學院張瞳老師饋贈。
1.1.2原料及處理
發酵原料為市售小麥、玉米、豌豆、大豆和豆粕。將原料除雜后進行粉碎,過40目篩,作為酵母菌發酵基礎料。
1.1.3培養基配制
1號試劑:酵母膏10 g,蛋白胨20 g,瓊脂粉20 g(液體培養基不加),蒸餾水900 mL;2號試劑:葡萄糖20 g,蒸餾水100 mL。將1號試劑瓶和2號試劑瓶分別配制滅菌,待溫度冷卻至60~70 ℃后混合均勻。
1.1.4試驗地點及時間
試驗地點為淮陰師范學院生命科學學院,試驗時間為2018年3—5月。
1.2試驗方法
1.2.1菌的活化及擴大培養
將酵母菌凍干粉瓶打碎后,用液體培養基溶解,吸取100 μL接種至固體培養基表面,均勻涂布,30 ℃培養24~48 h后,選擇菌落生長良好的培養皿,相同條件下進行劃線培養,挑取單菌落,裝入液體培養基中震蕩培養,菌懸液混合均勻后按照1 ∶50接種量擴大培養,測定菌液D值,制備接種菌液。
1.2.2發酵
每種原料稱取200 g,110 ℃加熱7 min,各種酵母的接種量均為1.0×107 cfu/g,原料與水分比例為1 ∶1。混合均勻后,裝入發酵袋中排空空氣靜置發酵,發酵溫度為30 ℃,發酵時間為24、48、72、96 h。每隔12 h檢查1次將袋中氣體排空,發酵完畢后樣品置于-20 ℃冰箱里待測。
1.3指標測定
1.3.1pH值的測定
取每株菌發酵的各時間段的每種發酵原料樣品5 g于小燒杯中,加入45 mL蒸餾水,每隔5 min攪拌1次,共攪拌3次,靜置10 min后,pH計測量。
1.3.2干物質回收率的測定
干物質測定方法參照GB/T 8303—2013《茶磨碎試樣的制備及其干物質含量測定》,計算公式:干物質回收率=(發酵原料干物質質量/發酵原料質量)/(原料干物質質量/原料質量)×100%。
1.3.3還原糖含量的測定
參照王俊剛等的3,5-二硝基水楊酸法測定[6]。準確稱取2.0 g樣品于100 mL三角瓶中,加入約50 mL蒸餾水,于50 ℃恒溫水浴中保溫20 min,期間不時攪拌使還原糖浸出。靜止三角瓶使樣品沉淀,吸取上清液8 mL于離心管中,3 000 r/min離心10 min,將上清液轉移至新離心管中,此即為還原糖測定樣品。還原糖含量以mg/g計算。
1.3.4粗蛋白含量的測定
依據GB/T 6432—1994《飼料中粗蛋白測定方法》凱式定氮法進行測定。稱取0.3 g左右測定干物質時烘干至恒重的樣品,加入催化劑和硫酸,進行消化,直至溶液透明呈藍綠色,待其冷卻,應用自動凱氏定氮儀測定其含氮量。
1.3.5植酸含量測定方法
采用分光光度法[7]測定植酸含量。步驟大致如下:(1)稱取0.5 g樣品,加入6 mL1.8 mol/L HCl,35 ℃水浴2 h,期間不斷振蕩,水浴后靜置過夜;(2)樣品4 000 r/min離心15 min,取上清液,加入0.4 mL鐵試劑,沸水浴45 min后再次離心,棄上清液;(3)用3 mL1.5 mol/L NaOH對沉淀進行溶解,再次離心,棄上清液;(4)加入1 mL 3.2 mol/L硝酸溶解沉淀后,依次加入0.2 mL 9%的檸檬酸三鈉、0.2 mL 1%對苯二酚、0.4 mL 0.4%鄰菲羅啉,純水定容至50 mL,pH值調節至3~4;(5)35 ℃水浴2 h后自然冷卻,于510 nm波長處測定吸光度。
計算公式:植酸含量=D510 nm×8.42÷樣品質量(mg)×125×100%。
1.3.6抗性淀粉含量的測定
參照高鑫(2012)的方法[8]。步驟大致如下:(1)準確稱取0.2 g樣品于帶塞玻璃試管中,加入蒸餾水0.4 mL,沸水浴20 min后冷卻至室溫;(2)加入HCl-KCl緩沖溶液(pH值為2.0)2.5 mL和10%胃蛋白酶105 μL,37 ℃恒溫振蕩1 h;(3)加入4 mL的醋酸鈉緩沖液(pH值=6.9)和淀粉酶-糖化酶混合溶液4 mL,37 ℃振蕩16 h;(4)4 000 r/min離心15 min,去掉上清液,50%乙醇溶液洗滌沉淀,重復2次;(5)依次加入2 mL 2 mol/L KOH溶液,8 mL 醋酸鈉緩沖液(pH值為3.8),100 μL糖化酶溶液,60 ℃ 水浴1 h;(6)冷卻至室溫,4 000 r/min離心15 min,保留上清液;(7)根據“1.3.3”節還原糖測定方法測定上清液中的還原糖含量。
計算公式:抗性淀粉含量=還原糖含量×0.9。
1.4數據處理
統計學分析采用SPSS 17.0(SPSS Inc.,Chicago,IL)軟件的One-Way ANOVA進行統計分析,計算結果以“平均值±標準誤”表示。P<0.05表示差異顯著。
2結果與分析
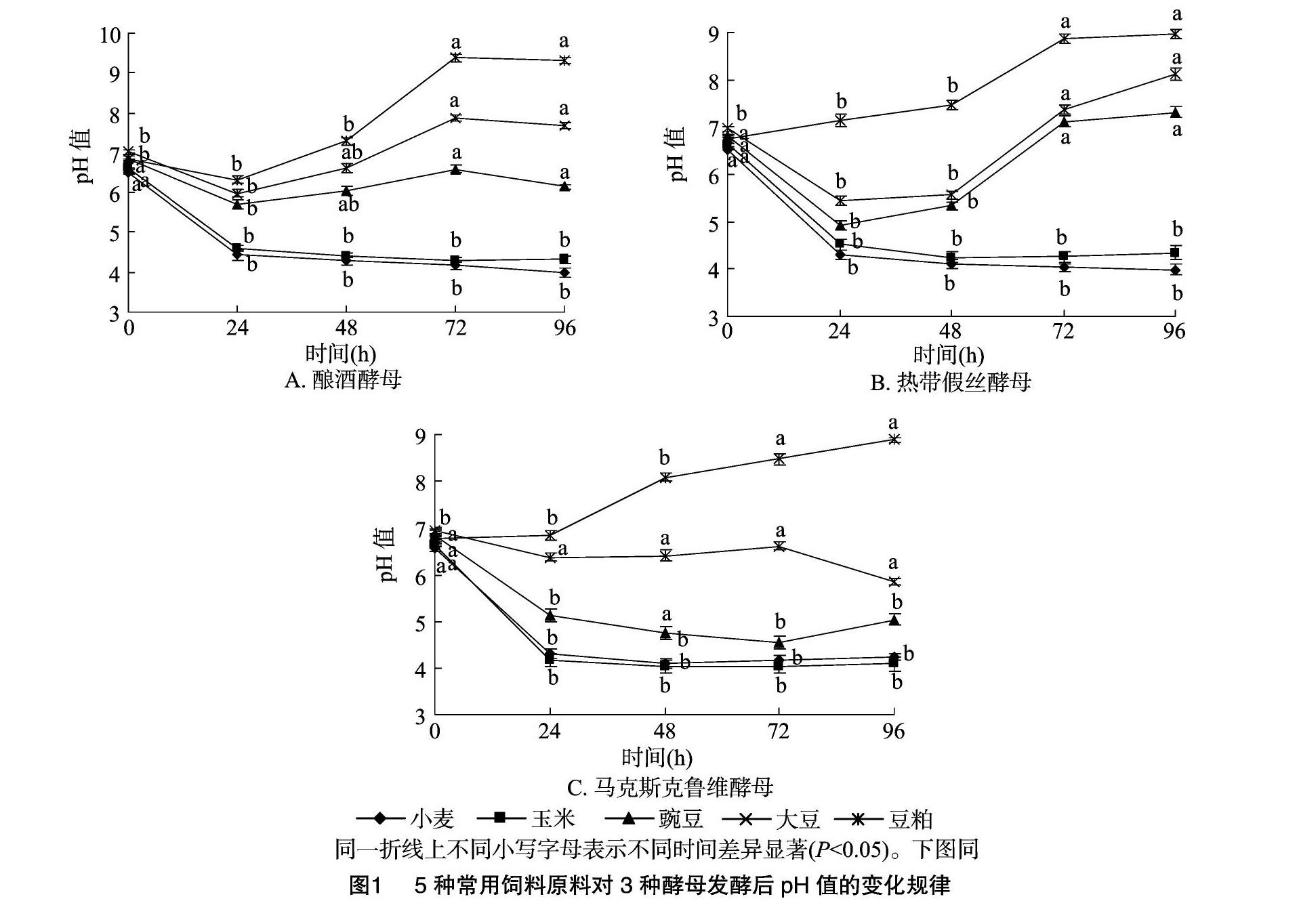
2.1不同飼料原料經3種酵母菌發酵后pH值的變化規律
由圖1可知,小麥和玉米分別經3種酵母發酵24 h后pH值均顯著降低(P<0.05),且于48~96 h內趨于穩定水平,呈明顯的酸性。豌豆和豆粕分別經釀酒酵母和熱帶假絲酵母發酵24 h后pH值均顯著降低(P<0.05),但隨后于72 h后逐漸恢復至初始pH值。大豆原料經3種酵母發酵72 h后pH值均顯著升高(P<0.05),呈明顯堿性。
2.2不同飼料原料經3種酵母菌發酵后干物質回收率的變化規律
由圖2可知,小麥原料經3種酵母發酵72 h后干物質回收率顯著降低(P<0.05),玉米原料經釀酒酵母發酵后干物質回收率呈現先降低后升高的趨勢,而經熱帶假絲酵母和馬克斯克魯維酵母發酵72 h和96 h后干物質回收率則顯著降低(P<0.05)。豌豆經釀酒酵母和熱帶假絲酵母發酵96 h后干物質回收率顯著降低,但不受馬克斯克魯維酵母發酵影響(P>0.05)。大豆分別經熱帶假絲酵母和馬克斯克魯維酵母發酵72 h和48 h后干物質回收率顯著降低(P<0.05),豆粕原料分別經釀酒酵母和馬克斯克魯維酵母發酵72 h后干物質回收率顯著降低(P<0.05),而經熱帶假絲酵母發酵72 h后干物質回收率卻顯著上升(P<0.05)。
2.3不同飼料原料經3種酵母菌發酵后還原糖含量的變化規律
由圖3可知,玉米原料分別經釀酒酵母和馬克斯克魯維酵母發酵24 h后還原糖含量顯著下降(P<0.05),而大豆、豌豆和豆粕原料經2種酵母發酵后還原糖含量均表現出先升高后降低的規律。豌豆原料經熱帶假絲酵母發酵24 h后還原糖含量顯著降低(P<0.05),而其他原料均表現出先升高后降低的規律。
2.4不同飼料原料經3種酵母菌發酵后粗蛋白含量的變化規律
由圖4可知,釀酒酵母發酵豆粕原料96 h時粗蛋白含量顯著降低(P<0.05),而發酵其他4種飼料原料對蛋白含量無顯著影響(P>0.05)。熱帶假絲酵母發酵小麥、玉米、大豆和豆粕4種原料72~96 h內粗蛋白含量均出現顯著升高現象(P<0.05)。馬克斯克魯維酵母發酵玉米24 h時,粗蛋白含量顯著升高(P<0.05),但隨后降低至初始水平,而發酵豆粕原料72 h后粗蛋白含量顯著升高(P<0.05)。
2.5不同飼料原料經3種酵母菌發酵后植酸含量的變化規律
由圖5可知,釀酒酵母發酵24 h后,大豆、豆粕、小麥植酸含量顯著降低(P<0.05);豌豆發酵72 h后,植酸含量的去除率達到49.3%(P<0.05);玉米的植酸含量在發酵48 h內無顯著變化(P>0.05)。熱帶假絲酵母發酵24 h時,大豆、豌豆、小麥植酸含量顯著降低(P<0.05);發酵48 h之后豆粕植酸含量的去除率達到55.3%(P<0.05),玉米的植酸含量無顯著變化(P>0.05)。馬克思克魯維酵母發酵24 h后,大豆、豆粕、小麥植酸含量顯著降低(P<0.05),玉米與豌豆發酵48 h 后植酸含量顯著降低(P<0.05)。馬克思克魯維酵母對大豆植酸含量降低效果最為顯著,對其植酸去除率達到68.1%。
2.6不同飼料原料經3種酵母菌發酵后抗性淀粉含量的變化規律
由圖6可知,釀酒酵母發酵48 h后,小麥原料的抗性淀粉含量顯著降低(P<0.05),而豌豆和玉米則在發酵72 h抗性淀粉含量顯著降低(P<0.05)。豌豆和小麥經熱帶假絲酵母發酵24 h后抗性淀粉含量顯著降低(P<0.05)。馬克思克魯維酵母發酵48 h后豌豆抗性淀粉含量降低44%(P<005);發酵72 h后,小麥與玉米抗性淀粉含量分別降低了43%和52%(P<0.05);大豆和豆粕則在發酵過程中抗性淀粉含量無顯著變化(P>0.05)。
3討論
pH值是生物發酵工藝中的一個非常重要的參數,飼料原料發酵過程中pH值降低主要歸結于發酵菌種增殖過程中產生并積累的有機酸[9],微生物發酵飼料中酸含量的增加反過來有利于飼料的長期保存[10]。本試驗中,小麥和玉米原料經釀酒酵母、熱帶假絲酵母和馬克斯克魯維酵母發酵后pH值均呈現顯著下降趨勢。研究發現,酵母菌本身產酸能力并不強,在發酵過程中可將葡萄糖代謝產物——丙酮酸轉化為乳酸、乙酸或乙醇[11],這可能是誘發原料pH值在發酵初期降低的因素之一。隨著發酵后期乙醇的積累,酵母細胞的生長受到抑制,丙酮酸代謝產酸途徑被切斷[12],因此pH值無法持續降低。也有研究人員認為,原料本身未被滅菌的乳酸菌在發酵過程中得到一定程度的增殖,且酵母菌增殖過程中產生的丙酮酸、琥珀酸等代謝產物能夠刺激乳酸菌的活動[13],為其提供營養物質,因而誘導原料pH值的迅速降低[14]。本試驗還發現大豆在酵母菌發酵后期pH值逐漸升高,說明蛋白類原料和淀粉類原料被酵母菌發酵的效果存在較大差異。作者推測這種差異可能歸結于酵母菌在不同發酵底物下積累的代謝產物不同所導致,在本試驗發酵過程中發現,大豆發酵后期出現腥味,可能是揮發性氨類和硫化物。劉倚帆等報道,酵母菌和米曲霉協同發酵整粒菜籽后期pH值也出現一定程度的升高[15]。因此,全脂大豆或菜籽中較高含量的蛋白質以及脂肪可能是酵母菌發酵后期pH升高的主要因素。本研究中豆粕和豌豆在3種不同酵母菌發酵下pH值呈現不同的變化規律也說明不同酵母菌在單一發酵底物下的代謝方式和途徑存在一定差異。
Terebiznik等認為益生菌固態發酵過程中干物質損失量和生物量呈正相關[16]。這一點在馬克斯克魯維酵母發酵中尤為明顯,酵母菌分解利用發酵底物中的物質合成自身成分,并釋放出代謝產物,在這一過程中,干物質量在發酵后期呈現明顯的下降趨勢。但本試驗也發現,熱帶假絲酵母發酵豆粕后期干物質回收率出現升高趨勢?說明發酵物料中的生物量增長速度超過了發酵底料的消耗速度,筆者推測這種現象可能只是短暫出現,隨著發酵的繼續進行,發酵底物的消耗速度勢必會超過生物量的增長。
酵母菌最重要的營養物質和能量來源大部分來自于單糖,而發酵纖維素等多糖的能力有限,這一點與枯草芽孢桿菌不同。大量研究報道,在果酒、泡菜等釀造發酵過程中,酵母菌能夠迅速降解底物中已有的還原糖成分,將其轉化為酸類、酯類或醇類物質[17-19]。本研究結果發現,玉米分別經釀酒酵母和馬克斯克魯維酵母發酵后還原糖含量持續下降,而大豆、豌豆和豆粕原料經2種酵母發酵后還原糖含量均表現出先升高后降低的規律。豌豆經熱帶假絲酵母發酵后還原糖含量持續降低,而其他原料均表現出先升高后降低的規律。由此說明,不同酵母菌作用于不同飼料原料底物,產生能夠分解多糖的酶的性質不同,后期當淀粉等多糖經酵母菌酶解成為還原糖后反過來又促進了酵母菌的增殖。
本研究結果表明,熱帶假絲酵母發酵小麥、玉米、大豆和豆粕4種原料可提高原料的粗蛋白水平,這一點優于釀酒酵母和馬克斯克魯維酵母。研究報道,生產上利用淀粉、葡萄糖、玉米深加工副產物等作為熱帶假絲酵母、釀酒酵母大量培養的底物,酵母本身蛋白質含量為45%~60%,其大量增殖的同時提高了發酵底物的粗蛋白含量[20-22],從而使底物的可飼性大大提高,可最大限度上替代魚粉等其他動物源蛋白質飼料。本研究中單一飼料原料的成分及含量均有所不同,可能是造成不同酵母增殖效果差異的重要因素。
植酸在植物種子的胚芽和谷物的麩皮中含量較高,尤其是在豆科植物的種子中含量最高。植酸可與常見的礦物元素產生不溶性化合物,大大降低其有效性,因而成為重要的抗營養因子之一。本試驗測定結果顯示,大豆和豌豆中植酸含量較高,但豆粕中植酸含量甚至低于玉米和小麥。這可能與大豆在壓榨獲得油脂過程中經歷高溫高壓等物理因素有關。研究報道,酵母菌能夠有效降解植物籽實中的植酸[23],因而有利于單胃動物對飼料中營養物質的吸收和利用,這可能與酵母菌能夠產植酸酶有直接關系[24-25]。本試驗中馬克思克魯維酵母在發酵48 h后各原料的植酸含量下降均較為明顯,顯示出較好的去除效果,因而在實際生產上具有一定的借鑒意義。
抗性淀粉是一種細胞壁成分,部分與水相溶形成粘性物質,阻礙了動物對營養物質的吸收。本試驗中酵母菌對玉米、小麥和豌豆等抗性淀粉含量較高的原料降解效果較好,而對大豆和豆粕的抗性淀粉降解效果較差。目前利用微生物發酵法降解植物籽實中的抗性淀粉尚未見報道。筆者認為不同原料中抗性淀粉的類型存在差異,酵母菌在不同底物上的發酵效果也有所不同,分解抗性淀粉可能主要靠分泌的纖維素酶或淀粉酶發揮作用,但由于酵母菌對復雜多糖的利用效果有限,因此僅依靠酵母菌發酵難以達到有效降解飼料中抗性淀粉的目的。
綜上所述,不同種類酵母菌對能量型和蛋白型飼料原料的發酵效果存在差異。總體上,經過3種酵母菌發酵后的能量型飼料(小麥和玉米)酸度顯著增加,而蛋白型飼料(豌豆、大豆和豆粕)經釀酒酵母和熱帶假絲酵母發酵后期pH值升高。通過控制菌種和時間可在發酵早期增加原料中還原糖和粗蛋白的含量,但隨著發酵的進行,還原糖逐漸被消耗。此外,3種酵母菌能夠有效降解飼料原料中的植酸,但僅對能量型飼料的抗性淀粉有一定的降解效果。
參考文獻:
[1]張宇紅. 不同復合菌比例發酵飼料對飼料營養價值及生長豬生產性能的影響[D]. 雅安:四川農業大學,2004.
[2]李龍. 復合益生菌發酵飼料工藝參數優化及品質評定[D]. 上海:上海交通大學,2010.
[3]劉艷新,劉占英,倪慧娟,等. 微生物發酵飼料的研究進展與前景展望[J]. 飼料博覽,2017(2):15-22.
[4]胡新旭,周映華,劉惠知,等. 無抗發酵飼料對斷奶仔豬生長性能、腸道菌群、血液生化指標和免疫性能的影響[J]. 動物營養學報,2013,25(12):2989-2997.
[5]王長彥. 微生物發酵飼料替代飼用抗生素技術在商品豬生產中的應用研究[D]. 楊凌:西北農林科技大學,2008.
[6]王俊剛,張樹珍,楊本鵬,等. 3,5-二硝基水楊酸(DNS)法測定甘蔗莖節總糖和還原糖含量[J]. 甘蔗糖業,2008(5):45-49.
[7]李桂榮. 燕麥籽粒植酸含量影響因素及生理機制研究[D]. 北京:中國農業大學,2007.
[8]高鑫. 山藥塊莖抗性淀粉形成的影響因素及理化特性的研究[D]. 呼和浩特:內蒙古農業大學,2012.
[9]劉曉明. 乳酸菌、枯草芽孢桿菌、酵母菌發酵不同飼料原料參數的研究[D]. 泰安:山東農業大學,2015.
[10]Muck R E. Factors influencing silage quality and their implications for management[J]. Journal of Dairy Science,1988,71(11):2992-3002.
[11]Hartikainen K,Poutanen K,Katina K. Influence of bioprocessed wheat bran on the physical and chemical properties of dough and on wheat bread texture[J]. Cereal Chemistry,2014,91(2):115-123.
[12]張秋美,趙心清,姜如嬌,等. 釀酒酵母乙醇耐性的分子機制及基因工程改造[J]. 生物工程學報,2009,25(4):481-487.
[13]Leroi F,Pidoux M. Detection of interactions between yeasts and lacticacid bacteria isolated from sugary kefir grains[J]. Journal of Applied Bacteriology,1993,74(1):48-53.
[14]崔晨曉,朱科學,郭曉娜,等. 酵母菌發酵對小麥麩皮成分的影響研究[J]. 中國糧油學報,2016,31(7):25-29.
[15]劉倚帆,王星凌,岳壽松,等. 復合微生物固態發酵整粒菜籽的研究[J]. 中國飼料,2017(1):36-40,44.
[16]Terebiznik M R,Pilosof,R A M. Biomass estimation in solid state fermentation by modeling dry matter weight loss[J]. Biotechnology Techniques,1999,13(3):215-219.
[17]王嘉怡,宋艷,呂曉華. 泡菜和酒渣中酵母菌的分離及其發酵作用初探[J]. 中國調味品,2017,42(7):25-28,32.
[18]朱娟娟,鄭少陽,李炎杰,等. 不同釀酒酵母對臍橙果酒發酵特性的影響[J]. 南方農業學報,2017,48(5):870-875.
[19]姜越,潘婷,惠竹梅. 模擬葡萄汁中可同化氮和還原糖對酵母發酵特性的影響[J]. 食品科學,2018,39(2):131-137.
[20]尹瑋,溫海琴,汪群慧,等. 玉米秸稈乳酸發酵殘渣制取飼料的研究[J]. 黑龍江大學自然科學學報,2008,25(4):480-484.
[21]張敏,侯先志,考桂蘭,等. 酵母菌發酵生產飼料酵母條件的優化[J]. 飼料工業,2010,31(8):28-30.
[22]賓冬梅,黃河清濤,易誠,等. 基于飼料酵母二次發酵啤酒糟條件響應面優化研究[J]. 湖南生態科學學報,2017,4(4):14-19.
[23]黃耀武,周建平. 發酵法降解油菜籽廢水中植酸的工藝研究[J]. 現代食品科技,2012,28(1):66-68,73.
[24]王晨祥,張苓花. 克魯斯假絲酵母植酸酶的篩選及其發酵條件[J]. 大連輕工業學院學報,2002,21(3):184-188.
[25]李曉宇,柳志強,田富,等. 響應面法優化熱帶假絲酵母ypy06植酸酶發酵條件的研究[J]. 熱帶農業科學,2011,31(6):41-44.
