細鱗鮭對水溶性氨基酸的嗅覺行為反應(yīng)
2018-12-27 07:25:50蔡紅英陳春山張旭光郭弘藝郭明磊
海洋漁業(yè) 2018年6期
關(guān)鍵詞:實驗
魏 凱,蔡紅英,陳春山,張旭光,郭弘藝,李 超,馬 龍,郭明磊
(1. 北京市水生野生動植物救護中心,北京 102100; 2. 上海海洋大學水產(chǎn)與生命學院,上海 201306;3. 北京市鱘魚鮭鱒魚創(chuàng)新團隊,北京 102100; 4. 中國水產(chǎn)科學研究院東海水產(chǎn)研究所,上海 200090)
細鱗鮭(Brachymystaxlenok),隸屬鮭形目(Salmoniformes),鮭科(Salmonidae),細鱗鮭屬,多分布于西伯利亞、朝鮮半島和中國的黑龍江、圖們江、鴨綠江等水域,在秦嶺和北京地區(qū)部分水系有零星分布,屬冷水性珍貴魚類。春季雨水引起河水上漲時,細鱗鮭由江河進入具有清水的支流中,一般生活在支流的山澗深潭中,秋季支流的水變淺時,由支流進入江河中越冬。隨著不同年份洪水的情況,細鱗鮭向主河道洄游的時間略有不同。由于環(huán)境惡化和棲息地環(huán)境變化等原因,細鱗鮭的分布區(qū)域逐漸縮小,野生資源量也日益減少,對細鱗鮭野生種群的保護已迫在眉睫[1]。目前,國內(nèi)對細鱗鮭繁殖生物學[1]、棲息地的地理環(huán)境因子[2]等展開了研究,但對其洄游行為的研究相對較少。鮭類洄游機制具有嗅覺印跡和性外激素誘導兩種假設(shè)[3-5]。最近,有研究提出大馬哈魚(Oncorhynchusketa)可利用其出生地的單體氨基酸進行洄游的假設(shè)[6]。支流水體中化學成份的研究表明,其常見的水溶性單體氨基酸有谷氨酸(Glu)、天冬氨酸(Asp)、甘氨酸(Gly)、丙氨酸(Ala)、絲氨酸(Ser)等[7-10]。細鱗鮭由主河道進入支流的習性,尚不清楚是否為支流水體中主要單體氨基酸對其嗅覺行為的影響。本實驗采用Y型迷宮,研究了4種主要水溶性氨基酸對細鱗鮭嗅覺行為的影響,以期一定程度闡明細鱗鮭的洄游行為機理。
1 材料與方法
1.1 實驗材料
實驗使用的細鱗鮭幼魚來自吉林省延邊細鱗鮭良種場,平均體質(zhì)量(6.0±2.4)g,平均體長(10.0±1.8)cm。試驗魚暫養(yǎng)于冷水機循環(huán)系統(tǒng)的圓形帆布水槽(1 m×1.6 m)中,平均水溫(13.0±1.5)℃,正常自然光照。暫養(yǎng)期間每日投喂一次人工顆粒餌料。實驗前從暫養(yǎng)水槽隨機取20尾健康的魚,分別放入位于水槽內(nèi)的隔離網(wǎng)籠進行分離暫養(yǎng)48 h,以適應(yīng)封閉環(huán)境,減少實驗應(yīng)激反應(yīng)。
1.2 實驗方法
實驗在自制的Y型迷宮水槽中(長80 cm×寬20 cm×高10 cm)進行,水槽的一端設(shè)有長30 cm的擋板,將水槽一端分成左右兩個同樣的選擇區(qū)A、B(圖1)。兩種相同濃度的氨基酸溶液通過調(diào)節(jié)閥以相同的速度(2 mL·s-1)流入選擇區(qū)。水槽另一端設(shè)有出水口,并通過水流閥控制氨基酸的擴散速度,調(diào)節(jié)氨基酸溶液擴散至選擇臂末端的時間為10~12 min。實驗時,試驗魚由活動網(wǎng)隔離在選擇區(qū)前方(圖1),適應(yīng)15 min后移去活動網(wǎng),同時打開氨基酸刺激液的調(diào)節(jié)閥,并同步視頻記錄試驗魚對不同選擇區(qū)的行為偏好反應(yīng)。為了避免兩種氨基酸在擋板末端混和,實驗記錄時間設(shè)置為10 min,并離線分析試驗魚的行為反應(yīng)。
分別測試了20尾細鱗幼魚對甘氨酸(Gly)、谷氨酸(Glu)、丙氨酸(Ala)、天冬氨酸(Asp)的選擇偏好性。前期預(yù)實驗表明,當氨基酸母溶液配制濃度為10-3摩爾每升(M)時,實驗水槽內(nèi)的氨基酸刺激液可對細鱗幼魚有較好的吸引效應(yīng)。經(jīng)計算,氨基酸母溶液擴散至擋板末端時實驗水槽內(nèi)實際濃度約為2.5×10-6M。為了比較試驗魚對上述氨基酸的偏好性,隨機選擇以甘氨酸為參照進行兩兩比較分析,分別進行了Gly vs Glu、Gly vs Ala、Gly vs Asp 3組實驗。實驗用水與暫養(yǎng)水同,以減少試驗魚的應(yīng)激反應(yīng)。
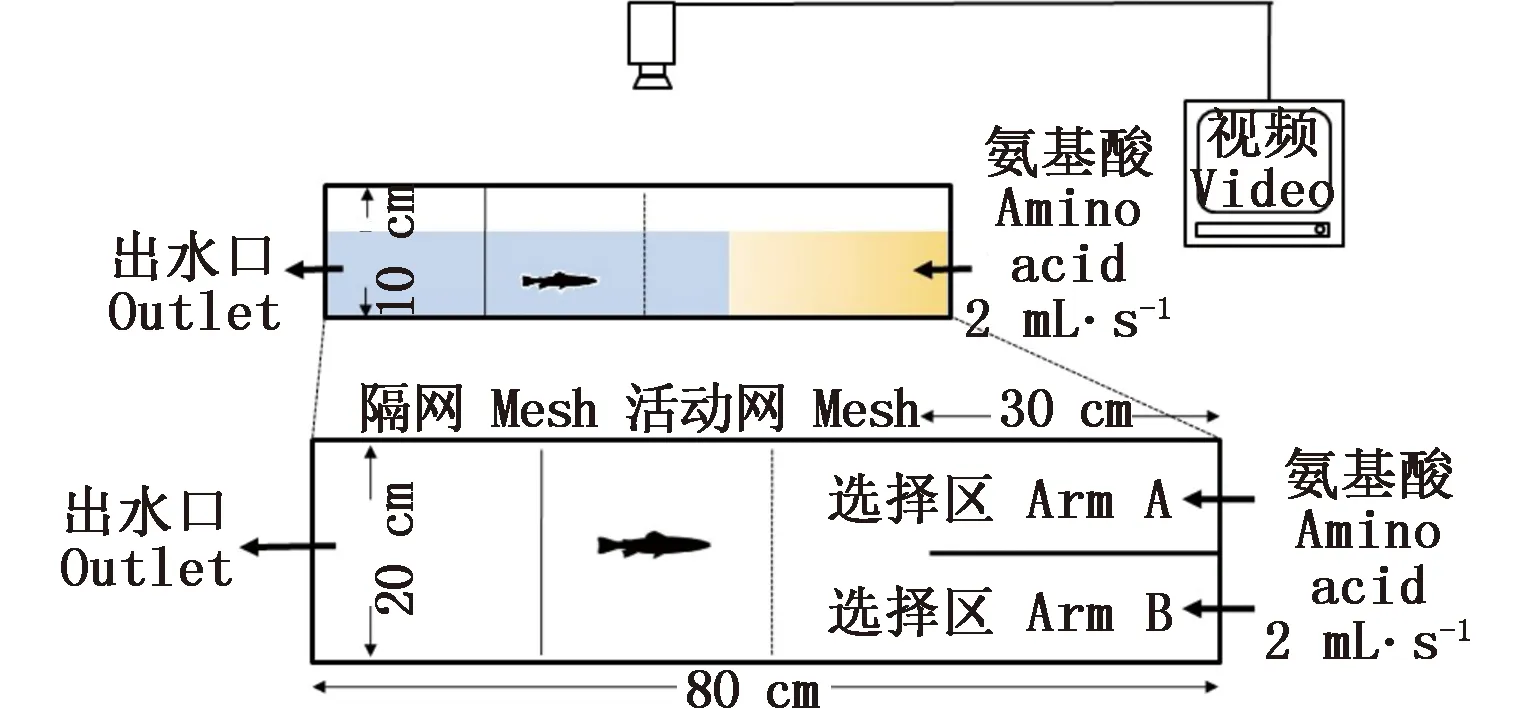
圖1 實驗裝置示意圖Fig.1 Schematic diagram of the Y-maze flow-through system
1.3 數(shù)據(jù)分析
實驗中,細鱗鮭幼魚對某一種氨基酸的行為選擇偏好性,主要因素為氨基酸種類。試驗魚選擇偏好性主要由試驗魚是否進入選擇區(qū),以及在選擇區(qū)的停留時間決定。依據(jù)ZHANG等[11]、張旭光[ 12]研究,試驗魚對某一氨基酸的絕對偏好度(preference degree)定義為P=(N×Ta)/Tl,式中:P為絕對偏好度;N為進入頻次;Ta為進入選擇區(qū)的平均停留時間(activity time);Tl為第一次進入選擇區(qū)的反應(yīng)時間或延遲時間(latency time)。該公式表明,絕對偏好度與試驗魚在選擇區(qū)的停留時間成正比,停留時間越長,選擇偏好性越大;與試驗魚第一次趨向選擇某種氨基酸的延遲時間成反比,延遲時間越長表明偏好性越小。
為了確定試驗魚群體對4種氨基酸的偏好性差異,將甘氨酸(Gly)的絕對偏好度標準化為1,根據(jù)公式Rx=Px/PGly進行標準化轉(zhuǎn)化計算對應(yīng)的氨基酸的相對偏好有效值[13],其中Rx為氨基酸的相對偏好有效值,Px為氨基酸的絕對偏好度,PGly為甘氨酸的絕對偏好度。根據(jù)相對偏好有效值的大小來確定細鱗鮭幼魚對不同氨基酸的行為選擇偏好性大小順序。所有數(shù)據(jù)均以平均值±標準誤(Mean±SEM)來表示,由Sigmaplot(Version 12.5)進行繪制圖表和顯著性分析。
2 結(jié)果與分析
2.1 嗅覺行為反應(yīng)
在實驗記錄時間內(nèi),大多數(shù)細鱗鮭幼魚(n=56,93%)都表現(xiàn)出對單體氨基酸的正趨向反應(yīng),但也有個別幼魚(n=4)在記錄結(jié)束才表現(xiàn)出趨向反應(yīng)。在經(jīng)過一定反應(yīng)延遲時間后能一次或數(shù)次進入前方的選擇區(qū),反應(yīng)延遲時間為289~398 s,平均時間為(329±14)s,該時間可能與氨基酸的擴散速度以及試驗魚對氨基酸嗅覺敏感性有關(guān)。細鱗鮭幼魚在選擇區(qū)通常停留一段時間,為12~400 s,平均停留時間為(90±11)s。實驗發(fā)現(xiàn)細鱗鮭幼魚(71%)會同時表現(xiàn)出對兩種氨基酸的偏好,它們往往選擇進入一側(cè)氨基酸區(qū)后也選擇進入另一側(cè)選擇區(qū),但實驗中也發(fā)現(xiàn)有的試驗魚(29%)只選擇在一側(cè)選擇區(qū)停留。
2.2 偏好度分析
在3組兩兩對比實驗中,試驗魚對參照組甘氨酸(Gly)的平均偏好度分別為(0.50±0.16)、(0.42±0.11)和(0.58±0.12),3組間試驗魚甘氨酸(Gly)的偏好度無顯著性差異(P>0.05)(圖2)。在Gly vs Ala比較組中,試驗魚對Gly的平均選擇偏好度(0.50±0.16)略小于Ala (0.60±0.13),但二者差異不顯著(P>0.05);但在Gly vs Glu組,試驗魚對Glu的平均偏好度(0.82±0.19)大于Gly(0.42±0.11),且二者差異達顯著性水平(P<0.05);在Gly vs Asp組,試驗魚對Asp的平均偏好度(0.70±0.15)略大于Gly(0.58±0.12),但二者未達顯著性差異水平(P>0.05)。
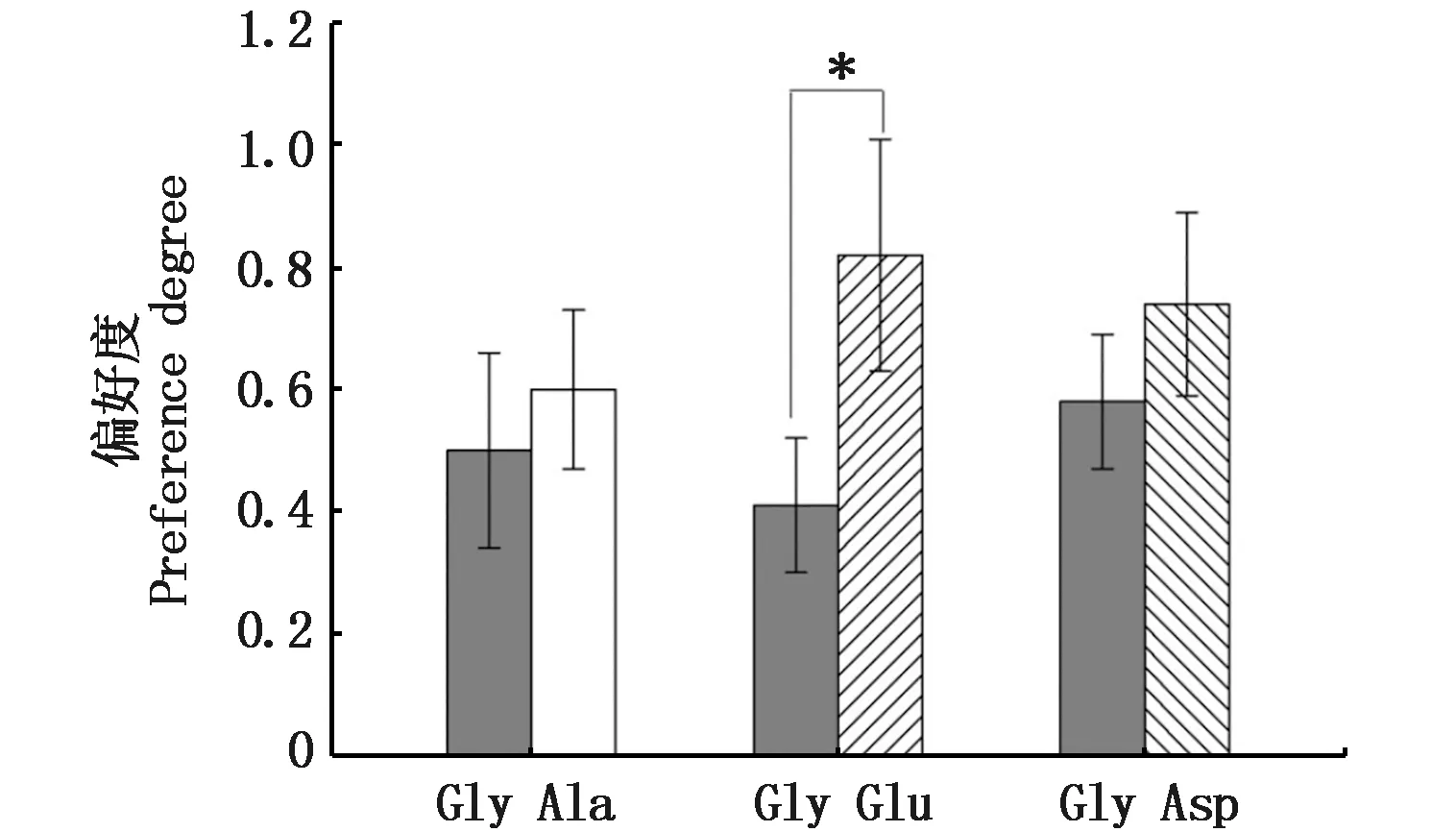
圖2 細鱗鮭幼魚對氨基酸的偏好度Fig.2 Preference degree to amino acids in Brachymystax lenok注:* 表示差異顯著(P<0.05)Note: * means significant differences (P<0.05)
根據(jù)Rx=Px/PGly計算,細鱗鮭幼魚對4種氨基酸的相對偏好度有效值分別為1(Gly)、1.18(Ala)、1.98(Glu)、1.27(Asp)。依據(jù)相對偏好度有效值大小,細鱗鮭對氨基酸的偏好性順序為谷氨酸(Glu)>天冬氨酸(Asp)>丙氨酸(Ala)>甘氨酸(Gly)。
3 討論
本實驗通過行為學的方法研究了細鱗鮭幼魚對谷氨酸(Glu)、甘氨酸(Gly)、天冬氨酸(Asp)、丙氨酸(Ala)4種單體氨基酸的偏好反應(yīng)。試驗魚對氨基酸的平均反應(yīng)延遲時間為300 s以上,表明試驗魚對氨基酸的反應(yīng)存在刺激閾值濃度。假設(shè)氨基酸母溶液以勻速擴散,那么引起試驗魚進入選擇區(qū)的實際氨基酸濃度約為1×10-6M,與引起魚類的嗅覺行為反應(yīng)的刺激閾值濃度相一致[14]。而高于閾值濃度的刺激則會引起魚類的逃避或抑制反應(yīng)[15]。這也可以解釋,試驗魚在進入選擇區(qū)后通常停留一段時間后離開的現(xiàn)象。
根據(jù)相對偏好度的有效值,細鱗鮭幼魚嗅覺反應(yīng)最敏感的為谷氨酸,其次為天冬氨酸、丙氨酸和甘氨酸。這可能與細鱗鮭嗅覺對氨基酸的生理敏感性有關(guān)。但對于上述4種氨基酸,虹鱒(Salmogairdneri)嗅覺電生理研究表明,其嗅覺上皮對丙氨酸(Ala)最敏感,其嗅電圖反應(yīng)的相對有效幅值較高,達108,而其它氨基酸的有效幅值較低(Gly:102; Glu:59.2; Asp:42)[16-17],虹鱒嗅覺上皮對氨基酸敏感性大小順序為:丙氨酸(Ala)>甘氨酸(Gly)>谷氨酸(Glu)>天冬氨酸(Asp)。雖然現(xiàn)階段沒有基于氨基酸刺激的細鱗鮭嗅覺電生理研究資料,但基于已報道的虹鱒的嗅覺電生理研究資料和本實驗數(shù)據(jù),推測細鱗鮭對氨基酸的行為敏感性可能與其嗅覺上皮的氨基酸生理敏感性相匹配。這一假設(shè)還需要在后續(xù)的實驗中開展細鱗鮭嗅電圖反應(yīng)的研究來驗證。
嗅覺行為研究也證實了單體氨基酸可以有效地刺激魚類的攝食行為[18]。例如,未開口的虹鱒和大西洋鮭幼魚,經(jīng)丙氨酸、甘氨酸、谷氨酸等單體氨基酸刺激,可誘發(fā)其攝食相關(guān)的行為反應(yīng),如咬合(snapping)、游泳加速(darting)等[19]。因此,鮭類對氨基酸的敏感性及其反應(yīng)可能是一種內(nèi)源性行為。單體氨基酸也可以誘發(fā)鮭類幼魚形成嗅覺印跡[20]。嗅覺印跡是魚類一種主要洄游機制,產(chǎn)生嗅覺印跡的因素還涉及各種性外激素[5, 21]或同類的氣味[3, 6]。行為學和電生理的研究都顯示單體氨基酸的嗅覺印跡是鮭類生殖洄游的重要機制[8, 20, 22-23]。Y型迷宮選擇實驗也表明,生殖期的馬蘇大馬哈魚(Oncorhynchusketa)對氨基酸種類和含量配比與其出生地相似的人工刺激液具有明顯的選擇偏好[22]。同時,該人工刺激液誘發(fā)了馬蘇大馬哈魚幅值較高的嗅電圖電位,而對無機鹽或膽酸配比相似的人工刺激液的反應(yīng)則較小[8]。這些研究都表明鮭類對氨基酸的敏感性在其洄游行為中起著重要的線索作用[8, 22-23]。
在對馬蘇大馬哈魚的棲息地天鹽川河(Teshio River)中水溶性氨基酸成分和含量進行的5年(2006-2010年)跟蹤調(diào)查表明,在自然條件下,氨其酸的成分是基本穩(wěn)定的,7種氨基酸丙氨酸、天冬氨酸、谷氨酸、甘氨酸、亮氨酸、絲氨酸和纈氨酸在5年內(nèi)變化差異較小,其中丙氨酸和天冬氨酸含量最高[8]。同樣,對細鱗鮭棲息河流的水溶性氨基酸調(diào)查也表明,其氨基酸總量為4.2~13.8 μM,其中單體氨基含量20%左右,約為0.84~2.76 μM[7]。本實驗中,氨基酸的實際效應(yīng)濃度為1.00~2.58 μM,與自然水域的氨基酸濃度相接近。細鱗鮭對谷氨酸保持的顯著敏感性,一方面由其嗅覺上皮的生理敏感性決定,另一方面與自然棲息水域中谷氨酸最高含量的特點相匹配。但同時,細鱗鮭也保持了對其它幾種主要氨基酸的行為偏好性。這表明,不同流域或其支流的主要氨基酸成分的差異可能決定不同種鮭類的氨基酸行為選擇偏好。馬蘇大馬哈魚對氨基酸比例相似的人工刺激液表現(xiàn)出與棲息地水溶液相似的反應(yīng)[8],也可以說明這一點。本實驗中,只驗證了試驗魚對單個氨基酸的行為反應(yīng),對兩種或多種的混和氨基酸行為反應(yīng)還待進一步研究。
細鱗鮭肌肉中的谷氨酸(1.75%干重)、天冬氨酸(1.59%)、甘氨酸(0.85%)和丙氨酸(0.75%),也是其所有氨基酸中含量較高的幾種[24]。每年的4-6月,性成熟的細鱗鮭有從中游溯河至上游進行生殖洄游的習性,繁殖后有部分親魚死于棲息地[25],單體氨基酸從魚體釋放和溶解可能是部分水體中的主要氨基酸的來源。另一方面,上游新鮮程度較高的有機物排放也影響水體中的氨基酸成分和含量變化。保護細鱗鮭野生資源的最終目標是使其能夠在自然條件下繁殖,恢復(fù)群體數(shù)量[25]。在細鱗鮭生殖期,保證其上游水體以及其溯河通道水域的氨基酸成分的穩(wěn)定是提高細鱗鮭溯河生殖洄游效率的重要保證。如今,人類活動的各種化學污染可能會阻斷細鱗鮭的洄游路徑,從而影響其自然繁殖的成功率。因此,對細鱗鮭野生資源的保護還涉及水體中氨基酸的長期監(jiān)控,以及時發(fā)現(xiàn)和控制排放源。
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數(shù)理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
