豬白細胞介素-2和融合抗菌肽重組酵母菌構建及其對小鼠免疫和生長的協同效應
2018-12-06 10:59:02胡冰吳雪穎馬常俊萬小平肖永樂陳建林李江淩呂學斌王澤洲高榮
四川動物 2018年6期
胡冰, 吳雪穎, 馬常俊, 萬小平, 肖永樂, 陳建林, 李江淩, 呂學斌*, 王澤洲, 高榮*
(1.四川大學生命科學學院,生物資源與生態環境教育部重點實驗室,四川省動物疫病預防與食品安全重點實驗室,成都610065; 2.四川省畜牧科學研究院,成都610066; 3. 四川省動物疫病控制中心,成都610035)
目前,動物傳染病仍然嚴重阻礙我國畜牧業和水產養殖的發展。動物飼料中添加的傳統化學藥物和抗生素導致動物病原體耐藥性持續增長,這仍然是傳染病控制和預防的巨大挑戰(Thorne,2007)。同時,由于免疫機能弱或動物免疫抑制等復雜因素,生產中經常出現疫苗免疫應答差、免疫保護率低下,嚴重妨礙了動物感染性疾病的防控(Shinetal.,2013)。因此,急需開發安全、高效、經濟的新型免疫調節劑。
許多細胞因子已被用作佐劑,以增強動物疫苗的原發性和記憶免疫應答(Kayamuroetal.,2010),對免疫系統有重大的影響,并具有塑造和引導調節免疫應答的能力(Plotkin & Plotkin,1999)。白細胞介素-2(interleukin-2,IL-2)參與T細胞增殖和Th調節反應,可增強機體的細胞免疫應答、刺激活化的B淋巴細胞增殖并誘導免疫球蛋白分泌(Jensonetal.,2016),是在調節先天性和適應性免疫中發揮關鍵作用的細胞因子,可以增強清除細菌和病毒的吞噬反應。
抗菌肽(antimicrobial peptides,AMPs)是大多數生物機體先天免疫的重要組成部分,在皮膚和黏膜表面發揮天然免疫作用,并對各種細菌、病毒、真菌和寄生蟲具有抗菌活性(Gordonetal.,2005)。AMPs具有保守的前肽序列,已在幾種哺乳動物中鑒定出(Bals & Wilson,2003)。除了直接抗菌的作用外,AMPs作為炎癥介質影響多種過程,如細胞增殖和遷移、免疫調節、傷口愈合、血管生成、細胞因子和組胺釋放(Wuerth & Hancock,2011)。因此,AMPs可作為創新藥物的原型處理感染或調節免疫反應(Bals & Wilson,2003;Niyonsabaetal.,2009)。
為了開發新型、經濟實用的免疫調節劑,本實驗通過2A自剪切技術構建重組酵母菌,共同表達豬IL-2和融合APMs基因,并探索對小鼠的免疫應答和生長的影響。
1 材料和方法
1.1 重組畢赤酵母Pichiapastoris及其構建
從重組真核質粒VRIL4/6-2中克隆了豬IL-2基因(標記為IL2-2A-a)的cDNA,并從重組VASP質粒中克隆出豬融合AMPs基因(標記為2A-a-P)的cDNA。質粒VRIL4/6-2和VASP都保存在本實驗室。從質粒pGAPZαA(Invitrogen)中克隆含有FMDV 2A肽和α因子基因以保證分泌表達2A-a片段。用TIANGEN的第一鏈反轉錄試劑盒逆轉錄得到cDNA。逆轉錄反應體系為37 ℃孵育60 min,得到總RNA的cDNA后,以cDNA為模板擴增目的片段。實時熒光定量PCR(qPCR)擴增后的產物用1.5%瓊脂糖凝膠電泳檢測。在cDNA片段中,根據實驗要求設計不同的N端和C端。然后,通過重疊延伸拼接技術將cDNA與片段IL2-2A-a、2A-a、2A-a-P結合,形成完整的IL2-P基因。
將豬融合AMPs基因(P)和IL2-P基因分別克隆到用GAP啟動子控制的pGAPZαA中,通過限制酶消化法將釀酒酵母Saccharomycescerevisiae的N端α-因子作為分泌信號。然后,將重組質粒(pGAPZαA-P和pGAPZαA-IL2-P)轉化到大腸桿菌EscherichiacoliDH5α中,并將陽性轉化體在含有25 μg·mL-1博來霉素的低鹽培養基(LLB)平板上篩選,通過直接PCR和測序鑒定。
根據Invitrogen手冊,通過電穿孔將80 μL巴斯德畢赤酵母SMD1168的電感受態細胞用10 μg(10 μL無菌水)的AVRⅡ-線性化重組質粒(pGAPZαA-P或pGAPZαA-IL2-P)或AVRⅡ-線性化pGAPZαA載體轉化。轉化后,將200 μL轉化細胞在1 mL 0 ℃的1 mol·L-1山梨糖醇中孵育,并在含有100 μg·mL-1博萊霉素的YPDS平板上選擇。將通過基因組分析和PCR篩選的陽性轉化體置于10 mL試管中培養,其中,3 mL YPD(1%酵母提取物、2%蛋白胨、2%D-葡萄糖)培養基在250 r·min-1、30 ℃下振蕩培養24 h,添加20%甘油,儲存于-80 ℃,其陽性轉化子分別標記為融合AMPs重組酵母菌(SGP)、IL-2和融合AMPs重組酵母菌(SG2P)、空質粒對照酵母菌(SG)。
1.2 SGP和SG2P的免疫生物活性測定
準備發酵上清液、胰蛋白酶消化的發酵上清液和胃蛋白酶消化的發酵上清液(酶終濃度為0.5 mg·mL-1,在37 ℃恒溫水浴中預孵育1 h),通過細胞計數法(CCK8法)測定在3種發酵上清液中表達重組蛋白的免疫生物活性。首先,參考Collins等(1994)的描述,豬淋巴細胞用豆球蛋白A(ConA)培養刺激制備成淋巴母細胞。每份樣品包括50 μL含5×105個淋巴母細胞的細胞懸液和50 μL樣品上清,一式3份,置于37 ℃、5%CO2烘箱中。孵育48 h后,每孔加入10 μL CCK8,在37 ℃、5%CO2烘箱中再孵育2 h。使用酶標儀680(Bio-Rad,USA)在450 nm下測定每個樣品的吸光度。
1.3 抗菌試驗
用大腸桿菌和金黃色葡萄球菌Staphylococcusaureus正常菌株、抗生素抗性大腸桿菌和金黃色葡萄球菌檢測重組蛋白的抗菌活性,菌株由四川大學生命科學學院王紅寧教授提供。在37 ℃將細菌用新鮮的LB培養基稀釋至5.0×105CFU·mL-1。將100 μL細菌懸浮液轉移到96孔板中,然后向每個孔中加入100 μL不同稀釋倍數的重組肽上清液,每個樣品做3次重復;并將含有不同抗生素的相同培養基[氨芐青霉素(Amp):100 μg·mL-1、200 μg·mL-1、400 μg·mL-1;卡那霉素(Kana):100 μg·mL-1、200 μg·mL-1]作為陽性對照加入到孔中。將平板置于37 ℃烘箱中孵育4 h,并通過酶標儀在600 nm測吸光度。
1.4 小鼠實驗
本實驗使用30只健康的21 d雌性ICR小鼠[四川大學實驗動物中心,生產許可證號:SCXK(川)2013-026,使用許可證號:SYXK(川)2013-185],隨機分為2個處理組(SGP、SG2P)和1個對照組(SG),每組10只(表1)。準備3種酵母菌株,在100 mL容量瓶中加入30 mL YPD培養基和300 μL博來霉素(100 mg·mL-1),接種之前保存的重組巴斯德畢赤酵母和空白巴斯德畢赤酵母,每個接種30 μL。然后在培養箱搖床中以30 ℃、250 r·min-1培養24 h,直到OD600約25。3種畢赤酵母菌(SG、SGP、SG2P)均置于上述相同的發酵條件下培養。
所有小鼠在整個實驗期間均飼喂相同飼料。在實驗的第0、7、14、21、28和35天,采集每只小鼠0.03~0.04 mL EDTA抗凝血液樣品,分析處理組酵母菌對小鼠免疫功能的影響。

表1 動物實驗分組及口服喂養方案Table 1 Grouping and oral feeding scheme of animal experiments
注: 處理組和對照組的OD600約25, 每組小鼠通過管飼法同時給藥4周
Note: The OD600of experimentals and control were about 25, each group of mice was simultaneously given by gavage for 4 weeks
1.5 攻毒試驗
在接種28 d后,為了檢測小鼠對感染的免疫力和保護作用,在實驗室將有毒的大腸桿菌(100 μL/小鼠)和金黃色葡萄球菌(100 μL/小鼠)注射到小鼠腹腔內(每種5只),每24 h監測并記錄所有小鼠,直到第7天將所有存活的小鼠安樂死并解剖觀察內臟和組織。
1.6 小鼠體質量
在接種后第0、7、14、21、28和35天測量每只小鼠的體質量,計算每組的平均體質量和凈增體質量,以評估重組酵母制劑對小鼠生長性能的影響。
1.7 血免疫細胞數量
使用1.4中收集的小鼠血樣,每組30 μL,通過MIND-RAY BC-3000血液自動計數儀測定血液免疫細胞、血小板和血紅蛋白含量。
1.8 Th細胞和Tc細胞
將購自eBioscience的抗小鼠CD4和CD8分子抗體分別用PerCP-Cy5.5和PE進行標記。50 μL小鼠靜脈血與50 μL生理鹽水混合,加入1 μL抗小鼠CD4 PerCP-Cy5.5(0.25 μg/Test)和1 μL抗小鼠CD8a PE(0.25 μg/Test),在黑暗中孵育30 min。然后加入1 mL(5%V/V)裂解溶液(Becton Dickinson,USA)5 min,以確保紅細胞完全溶解,存活的細胞用磷酸鹽緩沖液(PBS)洗滌2次,每次2 500 r·min-1離心5 min。最后,將細胞重懸于150 μL PBS中,并在FACScan流式細胞儀(Becton Dickinson,USA)中進行分析。
1.9 IgG、IgG1和IgG2a
小鼠IgG、IgG1和IgG2a定量ELISA試劑盒購自R & D Systems(USA)。方法參照小鼠IgG(或IgG1、IgG2a) ELISA試劑盒說明書。
1.10 qPCR分析免疫基因表達
100 μL血樣中加入1 mL RNAiso(TaKaRa),提取總RNA并在42 ℃反轉錄30 min作為向導基因(TransScriptTMOne-Step gDNA Removal and cDNA Synthesis SuperMix,TransGen)。根據GenBank上的相關基因序列,設計并合成了免疫基因PCR引物(表2)。
PCR程序為:95 ℃下進行初始變性3 min,然后在95 ℃ 10 s,60 ℃ 5 s,72 ℃ 10 s,循環40次,并在每次運行中進行陰性對照。以PPIA作為參照基因,使用幾何平均法和以下公式計算3組小鼠免疫相關基因的mRNA水平:相對水平=2-ΔΔCt。
1.11 統計分析
數據使用Systat 10(SPSS)進行統計學評估,通過雙因素方差分析和Tukey多重比較分析組間差異。當P<0.05時,差異有統計學意義。
2 結果
2.1 重組畢赤酵母的鑒定
用含有100 μg·mL-1博來霉素的YPDS培養基篩選重組酵母,然后提取其RNA并進行qPCR和電泳分析。結果表明,豬融合AMPs基因在SGP組中成功表達(圖1);豬IL-2和融合AMPs基因在SG2P組中成功表達(圖2)。

表2 qPCR引物Table 2 The primers for qPCR
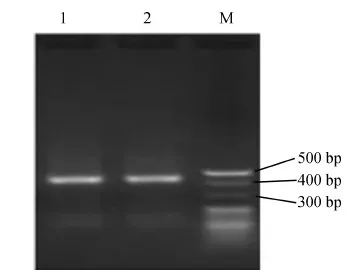
圖1 融合抗菌肽重組酵母菌qPCR電泳圖譜(1.5%瓊脂糖凝膠)Fig. 1 Electrophoresis of qPCR of recombinant Pichiapastoris with fusion antimicrobial peptide gene (1.5% agarose gel)
1、2.豬重組抗菌肽片段, M. 20 bp DNA Ladder Marker
1, 2. Porcine antimicrobial peptide fragment, M. 20 bp DNA Ladder Marker
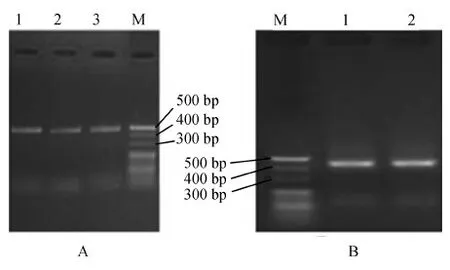
圖2 白細胞介素-2和融合抗菌肽重組酵母菌qPCR電泳圖譜(1.5%瓊脂糖凝膠)Fig. 2 Electrophoresis of qPCR of recombinant Pichiapastoris co-expressing porcine interleulkin-2 and fusion antimicrobial peptide gene (1.5% agarose gel)
A: 1、2、3.豬白細胞介素-2片段, B: 1、2. 豬融合抗菌肽片段; M. 20 bp DNA Ladder Marker
A: 1, 2, 3. Porcine interleukin-2 fragment, B: 1, 2. Porcine fusion antimicrobial peptide fragment; M. 20 bp DNA Ladder Marker
2.2 體外共表達IL-2和融合AMPs的生物活性
與對照組相比,3種發酵上清液的處理組均顯著促進ConA刺激的豬淋巴母細胞增殖(P<0.05),而處理組之間的差異無統計學意義(P>0.05)(圖3)。
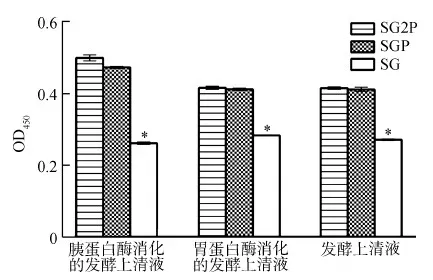
圖3 發酵上清液刺激豬淋巴母細胞的增殖
Fig. 3 The proliferation of porcine lymphoblasts stimulated with the supernatant samples
SG.空質粒酵母菌, SGP. 抗菌肽重組酵母菌, SG2P. 白細胞介素-2和融合抗菌肽重組酵母菌; 下同
SG.Pichiapastoris, SGP. recombinantPichiapastoriswith fusion antimicrobial peptide gene, SG2P. recombinantPichiapastorisco-expressing porcine interleukin-2 and fusion antimicrobial peptide gene; the same below
2.3 重組酵母抗菌活性分析
通過革蘭氏陰性菌和革蘭氏陽性菌分析重組酵母菌的抗菌活性。在初始濃度下,處理組的細菌生長比對照組受到更顯著抑制(P<0.05)。當稀釋1倍時,處理組的細菌生長也受到抑制,但與對照組的差異無統計學意義(P>0.05),且處理組間的差異無統計學意義(P>0.05)(圖4)。
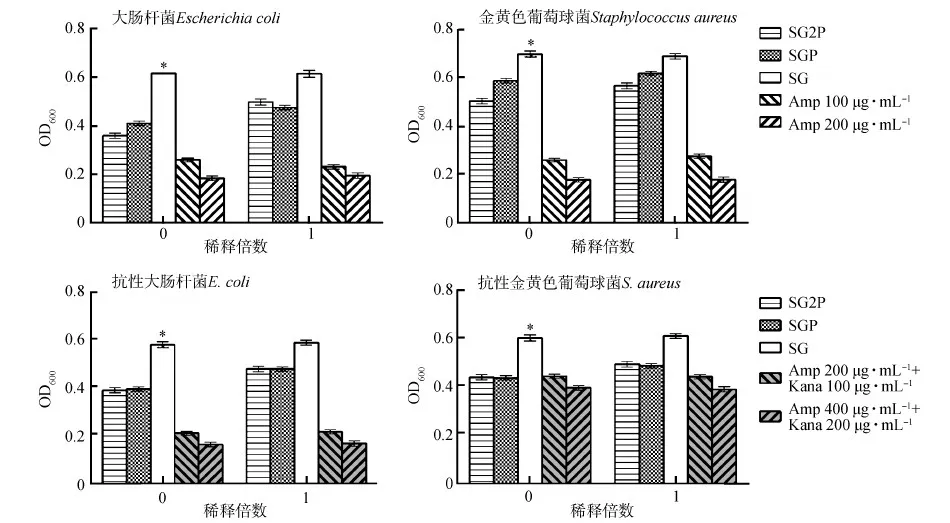
圖4 重組酵母菌的體外抗菌活性Fig. 4 In vitro antimicrobial activity of the recombinant Piciapastoris
Amp. 氨芐青霉素ampicillin, Kana. 卡那霉素kanamycin;*P<0.05; 下同the same below
2.4 攻毒后小鼠存活率
SG2P組和SGP組對大腸桿菌感染的保護率分別為100%和80%,而對照組的保護率僅為20%。同時,用強毒金黃色葡萄球菌注射小鼠腹腔7 d后,處理組的存活率為80%,對照組死亡率高達100%(圖5)。病理剖檢發現幸存小鼠的器官和組織正常,死亡小鼠病變明顯,肝臟和脾臟嚴重壞死,胃部擴散出血,十二指腸和空腸發生黏膜炎。
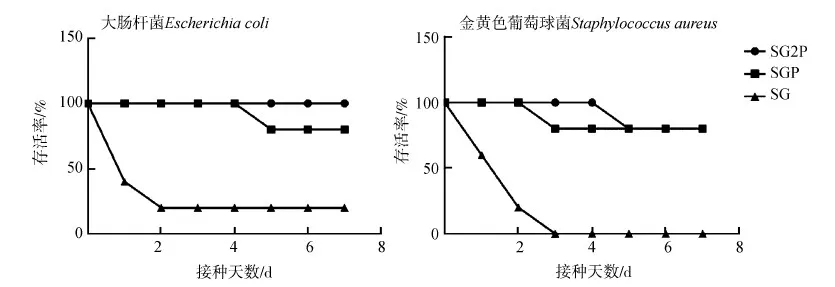
圖5 攻毒后小鼠存活率Fig. 5 Survival rates of mice challenged with virulent Escherichia coli and Staphylococcus aureus
2.5 小鼠體質量變化
飼喂一周后,處理組小鼠的體質量增加高于對照組(P<0.05)。處理組之間的差異無統計學意義(P>0.05)(圖6)。

圖6 小鼠的平均體質量變化Fig. 6 The average body mass change of mice after inoculation with SG, SGP and SG2P
2.6 外周血免疫細胞的變化
接種后7~35 d,處理組小鼠白細胞數量明顯高于對照組(P<0.05),但處理組間的差異無統計學意義(P>0.05);盡管實驗期間有一些波動,處理組與對照組紅細胞數量變化之間的差異無統計學意義(P>0.05);接種后7~35 d,處理組的血紅蛋白含量顯著高于對照組(P<0.05)(圖7)。
2.7 Th細胞和Tc細胞的變化
處理組小鼠血漿中的CD4+和CD8+T細胞數量在接種后7~35 d顯著高于對照組(P<0.05),SG2P組CD4+T細胞數量極顯著高于對照組(P<0.01);35 d時,CD4+和CD8+T細胞百分比均達到最高(圖8)。
2.8 IgG、IgG1、IgG2a的變化
接種后7~35 d,處理組小鼠的IgG、IgG1、IgG2a均高于對照組,35 d達到最高;但2個處理組間各指標差異無統計學意義(P>0.05)(圖9)。
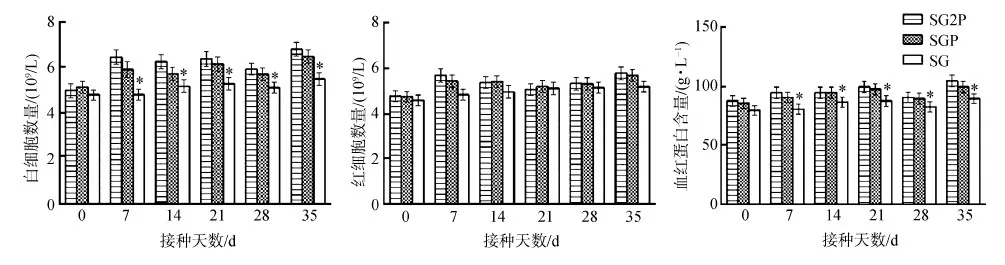
圖7 小鼠外周血中血細胞的變化Fig. 7 Change of the blood cells in the peripheral blood of experimental mice
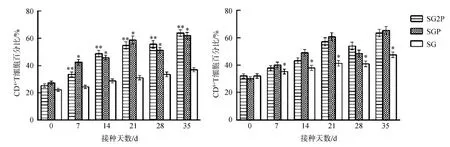
圖8 小鼠外周血中CD4+和CD8+ T細胞數量Fig. 8 CD4+和CD8+ T cells quantities in the peripheral blood of mice
**P<0.01
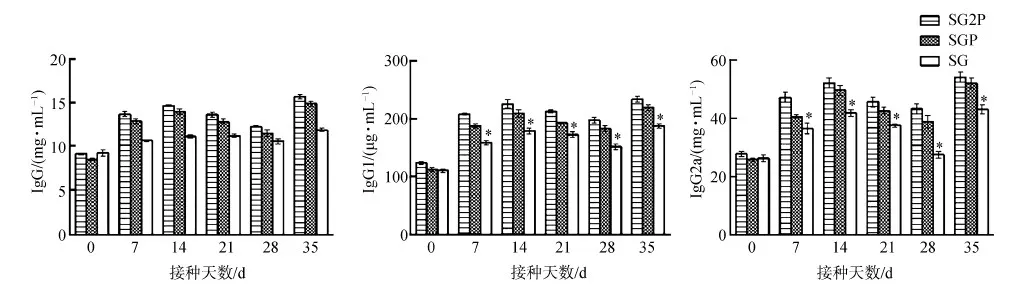
圖9 小鼠IgG、IgG1、IgG2a的變化Fig. 9 Changes of IgG, IgG1 and IgG2a in the mice
2.9 免疫基因表達的變化
2.9.1TLRs基因的變化接種后7~35 d,處理組4種TLRs基因表達水平均顯著高于對照組(P<0.05)。接種后35 d,這4種基因的表達水平達到最高。7~21 d,SGP組TLR6 mRNA水平明顯高于SG2P組(P<0.05)。2個處理組間TLR1、TLR4和TLR9表達水平差異無統計學意義(P>0.05)(圖10)。
2.9.2免疫記憶相關基因的表達變化接種后7~35 d,處理組的IL-7、IL-15、IL-23和CD62L基因的表達水平均顯著高于對照組(P<0.05),但IL-7基因的表達水平在接種第28天與對照組差異無統計學意義。這4個基因的表達水平在接種后35 d達到最高,2個處理組間的差異無統計學意義(P>0.05)(圖11)。
2.9.3細胞因子基因的表達變化處理組的IL-2、IL-4、IL-6和IL-12基因表達水平在接種后7~35 d顯著高于對照組(P<0.05),所有基因表達水平在35 d達到最大值。2個處理組間的基因表達水平差異無統計學意義(P>0.05)(圖12)。
2.9.4AMPs基因的表達變化接種后7~35 d,處理組的CRP4和CAMP基因表達水平均顯著高于對照組(P<0.05),接種后35 d達到最大值。2個處理組間的差異無統計學意義(P>0.05)(圖13)。
3 討論
細胞因子廣泛參與調節各種免疫反應,誘導細胞內信號通路觸發一系列生物過程,作為免疫佐劑具有重要的研究價值。目前,許多細胞因子已被用作佐劑以增強原發性和記憶免疫應答(Kayamuroetal.,2010)。關于IL-2、IL-12、IFN-γ等細胞因子的研究工作表明,當細胞因子與某種疫苗共同接種時,它們會調節抗原特異性免疫應答(Playfair & Souza,1987;Hsiehetal.,1993;Tagliabue & Boraschi,1993)。IL-2作為重要的T細胞生長因子,可以增強巨噬細胞和自然殺傷細胞的活性(Smith,1988;Seretietal.,2004),同時IL-2基因的轉錄和合成常被用作T細胞成功激活的關鍵指標。AMPs可以直接作用于多種病原體,發揮多功能效應因子天然免疫作用,如抑制結合DNA復制和細胞分裂、破壞細胞代謝通路、穿透細胞膜(Yangetal.,2001)。在本研究中,AMPs包含Tritrpticin、PR-39、PMAP-23和PG1基因。Tritrpticin具有回文序列、高度陽離子性質和色氨酸殘基中心簇(Lawyeretal.,1996),已被證明能有效對抗各種微生物、真菌和原生動物(Yangetal.,2002,2003)。PR-39是主要針對革蘭氏陰性菌如大腸桿菌和沙門氏菌Salmonella的有效抗生素,具有抗后生血管作用,抑制沙門氏菌侵入大腸上皮細胞(Isabeletal.,2012)。據報道,PMAP-23在無溶血活性的情況下,對革蘭氏陰性菌和革蘭氏陽性菌有顯著的抗菌活性(Parketal.,2002)。采用2A自剪切技術連接豬IL-2和AMPs基因,研制出一種新型免疫調節分子,使多種基因在同一構建體內表達(Szymczak & Vignali,2005)。
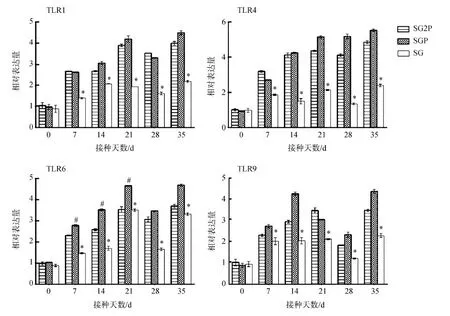
圖10 小鼠TLR1、TLR4、TLR6、TLR9基因的表達水平Fig. 10 Relative expression levels of TLR1, TLR4, TLR6 and TLR9 genes in the mice
#SG2P vs. SGP:P<0.05
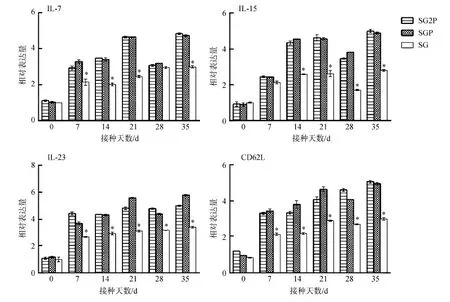
圖11 小鼠IL-7、IL-15、IL-23和CD62L基因的表達水平Fig. 11 Relative expression levels of IL-7, IL-15, IL-23 and CD62L genes in the mice
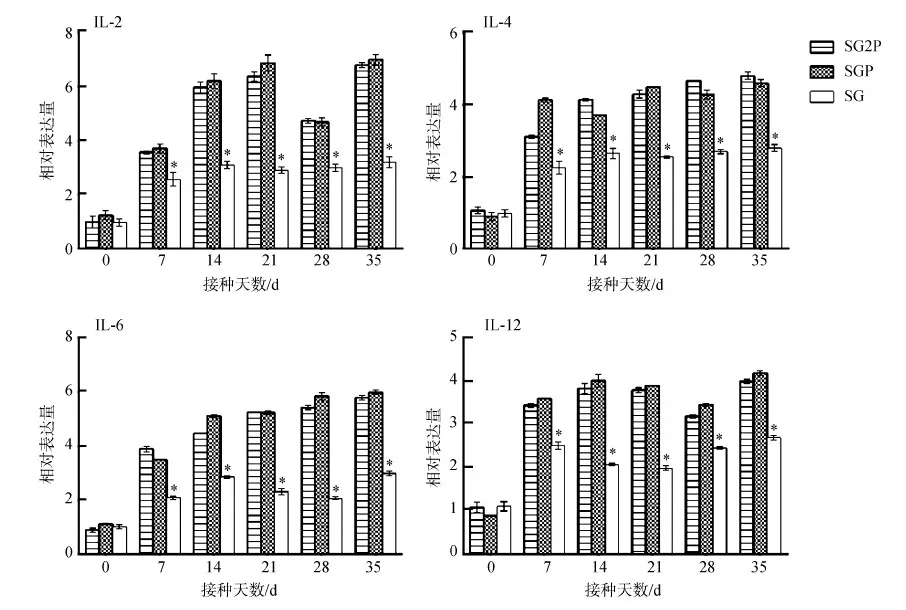
圖12 小鼠IL-2、IL-4、IL-6和IL-12基因的表達水平Fig. 12 Relative expression levels of IL-2, IL-4, IL-6 and IL-12 genes in the mice
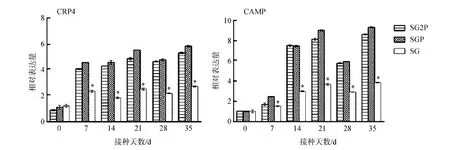
圖13 小鼠CRP4和CAMP基因的表達水平Fig. 13 Relative expression levels of CRP4 and CAMP genes in the mice
在免疫細胞增殖和抑菌試驗中,發現SG2P組發酵上清液對體外培養的豬淋巴母細胞的增殖效應明顯強于對照組,與SGP組發酵上清液差異無統計學意義,說明重組體SGP和SG2P具有比SG更好的免疫生物活性;酶消化的處理組發酵上清液也有顯著的免疫生物活性。SGP組和SG2P組細菌的生長受到明顯抑制。這些結果表明,重組體體外具有免疫生物活性和殺菌活性。小鼠體內實驗發現,處理組的重組酵母顯著提高了TLRs基因(TLR1、TLR4、TLR6、TLR9)、免疫記憶相關基因(IL-7、IL-15、IL-23、CD62L)、細胞因子基因(IL-2、IL-4、IL-6、IL-12)以及AMPs基因(CRP4和CAMP)的表達水平;同樣,處理組小鼠血液中Th和Tc細胞、IgG、IgG1和IgG2a含量明顯增加;與此相應,強毒細菌注射攻毒也證實處理組小鼠比對照組呈現更高的存活率。
據報道,TLRs在檢測哺乳動物和昆蟲中的微生物感染方面具有關鍵作用(Medzhitov,2001),這些受體通過識別不同病原體中的保守分子模式參與先天免疫應答,并且在激活病原體中發揮核心作用——特異性體液和細胞適應性免疫應答(Kumaretal.,2009;Kawai & Akira,2010)。Th1細胞產生IFN-γ、TNF-α、IL-2和IL-12,對細胞免疫應答有關鍵作用。Th2細胞產生IL-4、IL-6和IL-10,主要調控體液應答反應(Parronchietal.,1991;Romagnani,1991;Sher & Coffman,1992)。免疫記憶相關基因對于特異性免疫細胞的發育、增殖、分化和存活是必需的(Stevcevaetal.,2006)。而細胞因子基因和AMPs基因在動物免疫反應中也發揮著重要作用。因此,血液中這些免疫基因表達和白細胞的顯著增加,表明重組酵母可以明顯增強小鼠的先天和適應性免疫反應。
因此,重組酵母共表達豬IL-2和融合AMPs能顯著增強小鼠的先天免疫和獲得性免疫力,提高動物的系統性體液和細胞免疫應答,提示可研發成為新型安全高效的免疫調節劑,增強動物對傳染病的免疫抗病力。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
發明與創新·初中生(2024年6期)2024-06-16 00:00:00
昆明醫科大學學報(2022年1期)2022-02-28 07:43:38
軍事文摘·科學少年(2021年1期)2021-02-04 08:03:45
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
閱讀與作文(小學低年級版)(2015年8期)2015-05-30 10:48:04
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
食品工業科技(2014年9期)2014-03-11 18:15:31
食品科學(2013年19期)2013-03-11 18:27:43
