海拔梯度下元謀干熱河谷植物群落特征
2018-12-05 06:47:02余嬌娥司宏敏吳雪濤張彧蘇文華周睿
生態(tài)環(huán)境學(xué)報 2018年11期
關(guān)鍵詞:物種
余嬌娥,司宏敏,吳雪濤,張彧,蘇文華,周睿*
1. 云南大學(xué)生態(tài)學(xué)與地植物學(xué)研究所,云南 昆明 650500;2. 吉林大學(xué)環(huán)境與資源學(xué)院,吉林 長春 130023
植物群落是植物在長期環(huán)境變化中自我調(diào)節(jié)和適應(yīng)而形成的集合體(Lieth,1975),物種多樣性是群落最基本的特征,是形成群落結(jié)構(gòu)的基礎(chǔ)。研究群落的物種組成及多樣性是認識植物群落結(jié)構(gòu)、生態(tài)、動態(tài)的基礎(chǔ)(王國明等,2017;許傳陽等,2014)。一般認為,物種多樣性是由資源可利用程度決定的(Morin,2000),會隨著生境的干旱的程度而降低(Bello et al.,2006;劉小丹等,2014)。α多樣性以及β多樣性構(gòu)成物種的總體多樣性。在植物群落多樣性研究中,群落α多樣性是反映群落內(nèi)物種組成狀況的一個重要指標,即物種豐富度和均勻度,β多樣性表示物種沿環(huán)境梯度發(fā)生替代的程度或物種變化的速率,表示生境的變異程度或指示生境被物種分隔的程度(馬克平等,1995)。植物群落α、β多樣性是當(dāng)前群落生態(tài)學(xué)乃至整個生態(tài)學(xué)研究中十分重要的內(nèi)容(David,1994;馬斌等,2008;陳杰等,2012;張曉龍等,2017)。研究植物群落多樣性特征對揭示群落、生態(tài)系統(tǒng)結(jié)構(gòu)和功能演變具有極為重要的意義(Pandey et al.,2003)。
植被蓋度(Vegetation coverage)通常是指植被冠層垂直投影面積占基準地表單位面積的比例或百分比(Purevdorj et al.,1998),是表示陸地表面植被數(shù)量及反映生態(tài)系統(tǒng)變化的重要基本參數(shù),也是地面蒸散、光合作用等多種地表過程研究的控制性因子,也是反映森林資源的一個重要參數(shù)(Price,1993;張云霞等,2003)。植被覆蓋及其變化不僅是區(qū)域生態(tài)系統(tǒng)環(huán)境變化的重要指標,而且對水文、生態(tài)、全球變化等都具有重要的意義(陳曉光等,2007)。
干熱河谷是對分布于中國西南地區(qū),具備干、熱兩個基本屬性河谷帶狀區(qū)域的總稱,該區(qū)域由于地形深陷封閉而產(chǎn)生的雨影作用以及海拔下降絕熱增溫和谷地焚風(fēng)效應(yīng)而形成了西南地區(qū)特殊的地形氣候類型(金振洲等,1987)。所謂“干熱”就是極端水分條件與熱量條件的結(jié)合。其中,“干”方面,全年的蒸發(fā)量大于降水量的3~6倍;“熱”方面,年均溫>20 ℃,>10 ℃年積溫>7000 ℃。全年干濕季分明,水分和光熱條件失衡,熱量高、降水量少以及蒸發(fā)強是干熱河谷氣候的典型特征(何永彬等,2000;張建平等,2000)。這一特殊氣候特點導(dǎo)致該區(qū)域谷底(海拔約1100 m)植被為稀樹灌草叢組成的“河谷型”薩瓦納植被(Savanna of valley type,金振洲,1999),而河谷頂部(海拔約2100 m)則分布有常綠闊葉林。從植物區(qū)系來看,1600 m以下河谷中植物有明顯的熱帶性質(zhì),而河谷頂部亞熱帶成分增加(歐曉昆,1988),表現(xiàn)出明顯的物種更替現(xiàn)象。劇烈的氣候和群落變化被壓縮在1000 m的海拔落差中,其間物種適應(yīng)特征、物種組成和生物多樣性也必然發(fā)生重大改變。該地區(qū)早期就有人類活動,是人類文明的發(fā)源地之一,是中國西部人口密集、開發(fā)較早的山地農(nóng)業(yè)區(qū),是云南農(nóng)業(yè)經(jīng)濟發(fā)展熱區(qū)(朱鐘麟等,2006)。社會資源的高度開發(fā)和利用以及其他人為因素(如放牧、開墾耕地等)直接或間接造成了生態(tài)環(huán)境的進一步退化和破壞(張建利等,2010)。且從 20世紀 50年代至今,該地區(qū)大肆開展植物造林工程,主要引進種植了以赤桉(Eucalyptus camaldulensis)、檸檬桉(Eucalyptus citriodora)、大葉相思(Acacia auriculaeformis)以及豆類等物種(李昆等,1999;李昆等,2004),迅速而直接地改變了干熱河谷的景觀類型、組分結(jié)構(gòu)及空間格局。該區(qū)域干濕季分明的氣候特征也造就了植被外貌明顯的季節(jié)性變化,旱季很多物種采用避旱對策呈現(xiàn)假死狀態(tài),只有很少完全適應(yīng)當(dāng)?shù)貥O端干熱環(huán)境的物種能在旱季保持常綠,這些物種是組成當(dāng)?shù)刂参锶郝涞闹匾锓N,是維持生物多樣性的基石。此外,雨季有人工種植物種逸出,這些物種未必適宜當(dāng)?shù)貧夂颍瑫ι锒鄻有哉{(diào)查造成一定干擾。
本研究從干熱河谷底部到兩側(cè)山地沿海拔梯度設(shè)置樣方,探索沿海拔梯度群落結(jié)構(gòu)的變化。試圖回答以下科學(xué)問題:該地區(qū)的群落結(jié)構(gòu)變化是怎樣的?物種替代現(xiàn)象主要發(fā)生在哪個海拔區(qū)間?如何劃分干熱河谷與常綠闊葉林植被的分布區(qū)域?研究結(jié)果不僅可為干熱河谷地區(qū)物種多樣性的深入了解提供科學(xué)依據(jù),還可為干熱河谷的植被恢復(fù)提供依據(jù)。
1 材料與方法
1.1 研究區(qū)概況
元謀干熱河谷位于金沙江流域干熱亞區(qū)(何毓榮 等 , 1999 ) , 地 理 位 置 25°31′~26°07′N ,101°36′~102°07′E,是金沙江干熱河谷的典型區(qū)域。屬南亞熱帶干熱季風(fēng)氣候。隨海拔高度升高及順河而上氣溫下降,濕度和降水量增加以及蒸發(fā)量減小,輻射增強。谷底干熱及沿海拔梯度的水熱急劇變化是干旱河谷生態(tài)環(huán)境的基本特點之一(周志瓊等,2008)。年均降雨量為642.2 mm,年平均蒸發(fā)量為 3911.2 mm,蒸發(fā)量大約是降雨量的6.2倍,表現(xiàn)了干熱河谷干旱的特征;多年平均氣溫為21.9 ℃,≥10 ℃年積溫為8003 ℃,年日照時數(shù)約為 2670.4 h(崔永忠等,2006)。其氣候與稀樹草原(Savanna)氣候相似(Ratnam et al.,2016)。基帶土壤為燥紅土,另有變性土、薄層土及紫色土。自然植被以坡柳(Dodonaea viscosa)+扭黃茅(Heteropogon contortus)群系為主(金振洲等,1987)。主要喬木樹種有云南松(Pinus yunnanensis)、余甘子(Phyllanthus emblica)、滇刺棗(Ziziphus mauritiana);主要灌木有坡柳、疏序黃荊(Vitex negundo)、小石積(Osteomeles anthyllidifolia)、鐵仔(Myrsine africana)、金合歡(Acacia farnesiana)、清香木(Pistacia weinmannifolia);主要草本物種有扭黃茅、孔穎草(Bothriochloa pertusa)。
1.2 樣地設(shè)置
本研究采用樣方調(diào)查法,于元謀干熱河谷的2017年旱季時期(2017年4月)共設(shè)置了10個海拔梯度。從海拔1100 m左右開始,每隔100 m海拔根據(jù)地形條件設(shè)置4個10 m×10 m的樣方。在10 m×10 m的大樣方內(nèi)再設(shè)置3個1 m×1 m的小樣方內(nèi)調(diào)查草本層。共設(shè)置40個10 m×10 m的樣方,120個 1 m×1 m的小樣方。喬木層記錄所有胸徑(DBH)≥1 cm的物種及株數(shù)、高度、胸徑和蓋度,灌木層及草本層記錄所有種的株數(shù)、高度和蓋度。胸徑用圍尺測量,高度和蓋度采用目測法。其中,總蓋度為10 m×10 m樣方內(nèi)所有植物垂直投影面積占樣方面積的比例,灌木層蓋度為10 m×10 m樣方內(nèi)灌木植物垂直投影所占的比例,草本層蓋度為10 m×10 m樣方內(nèi)草本植物垂直投影所占的比例,分層蓋度加和值大于或等于總蓋度。同一海拔4個樣地的蓋度為4個樣方的平均值。
1.3 數(shù)據(jù)分析
1.3.1 數(shù)據(jù)處理
運用Office WPS(北京金山辦公軟件股份有限公司和珠海金山辦公軟件有限公司)對實驗數(shù)據(jù)進行初步整理與計算。運用SPSS 13.0(Chicago,IL,USA)對海拔與物種蓋度、物種多樣性進行Pearson相關(guān)性分析。
1.3.2 α多樣性計算
本研究采用了4種常用的物種多樣性指數(shù)(姜雋,2009;劉雨芳,2003;馬克平等,1995;王飛等,2017):豐富度指數(shù)(S)、Shannon-Wiener物種多樣性指數(shù)(H′)、Pielou群落均勻度指數(shù)(J)以及Simpson生態(tài)優(yōu)勢度指數(shù)(D)進行群落特征分析,其計算公式如下:

式中,S為物種數(shù)目;N為群落中的物種數(shù);pi為屬于種i的個體占全部個體的比例。
1.3.3 β多樣性計算
該指標是反映不同環(huán)境梯下群落之間物種組成的相異性或物種沿環(huán)境梯度的更替速率,表示生境的變異程度或指示生境被物種分隔的程度。不同環(huán)境梯度中不同樣地之間共有種越少,β多樣性就越高。一般采用相異性指數(shù)和Cody指數(shù)進行表征,其計算公式如下(孫榮等,2011):
相異性指數(shù)(SI):

Cody 指數(shù)(βc):

式中,a和b分別為兩樣方的物種數(shù);c為兩樣方的共有物種數(shù);g(H)是沿生境梯度H增加的物種數(shù)目;l(H)是沿生境梯度 H失去的物種數(shù)目,即在上一個梯度中存在而在下一個梯度中沒有的物種數(shù)目;相異性指數(shù)(SI)反映群落或樣方間物種組成的相似性;Cody指數(shù)反映物種沿梯度的替代速率。
2 結(jié)果與分析
2.1 植被蓋度的變化趨勢

圖1 蓋度隨海拔的變化趨勢Fig. 1 Tendency of coverage to change with elevation
由圖1可知,隨著海拔的升高,群落總蓋度與灌木層蓋度整體上均呈現(xiàn)出了隨之增高的趨勢。植被總蓋度整體波動相比灌木層的蓋度波動大。在海拔1400~1500 m處植被總蓋度出現(xiàn)一個急劇下降的趨勢,海拔1500 m之后,蓋度又有一個回升的趨勢,且在海拔2000 m時,植被總蓋度達到最大值。灌木層蓋度隨海拔高度逐漸上升,在海拔1100 m時,其蓋度最低,海拔2000 m時,灌木層蓋度同樣達到最高值。草本層蓋度則呈緩慢下降的趨勢,但未有較大的變化。表1顯示,海拔高度與群落總蓋度在P<0.05水平上顯著正相關(guān);海拔高度和灌木層蓋度在P<0.01水平上呈現(xiàn)極顯著正相關(guān);同時群落總蓋度和灌木層蓋度以及群落總蓋度與草本層蓋度在P<0.01水平上都存在極顯著正相關(guān)。由相關(guān)性分析可以得知,海拔與總蓋度存在顯著正相關(guān)關(guān)系,證明海拔的變化影響了植物蓋度的變化,海拔升高,植物總蓋度增加。分析其原因,河谷海拔越高,其水熱環(huán)境越好,越適宜植物生長故植物蓋度越高。而灌木及喬木層蓋度增加,導(dǎo)致了光熱條件的重新配置,使得草本層蓋度有所降低。海拔變化引起群落蓋度的變化,群落總蓋度受灌木層、草本層以及喬木層蓋度的共同作用。
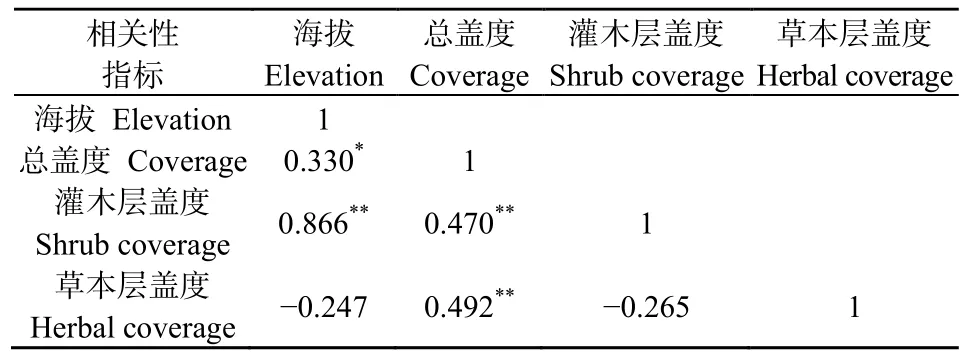
表1 海拔與蓋度相關(guān)性分析Table 1 Analysis of the correlation between elevation and coverage
2.2 α物種多樣性的變化趨勢
表2顯示了元謀干熱河谷不同海拔梯度灌木群落的α物種多樣性測定結(jié)果。分析可知,Gleason指數(shù)(D1)和Marglef指數(shù)(D2)變化最大,說明隨著海拔升高,灌木群落物種多樣性變化較大,且總體呈現(xiàn)出隨著海拔升高,物種多樣性增強的趨勢。

表2 元謀干熱河谷不同海拔梯度灌木群落的的a多樣性指數(shù)Table 2 A diversity index of shrub communities at different elevation in dry-hot valley of Yuanmou
在海拔 2000 m時,物種豐富度達到最大值(15);海拔1100~1500 m時,物種豐富度比較穩(wěn)定,未發(fā)生較大變化。自海拔1500 m之后,物種豐富度迅速上升,且隨著海拔的升高呈現(xiàn)出物種豐富度增強的趨勢。Shannon-Wiener指數(shù)、Pielou指數(shù)隨海拔變化的趨勢基本與豐富度指數(shù)一致,在海拔2000 m處物種最為豐富,分布均勻,但Simpson指數(shù)的變化趨勢與之相反,整體上隨著海拔的升高呈現(xiàn)出降低的趨勢。
圖2顯示了元謀干熱河谷不同海拔梯度α物種多樣性中的Simpson指數(shù)和Shannon-Wiener指數(shù)的測定結(jié)果。從圖2中可以看出,Shannon-Wiener指數(shù)隨海拔梯度整體上呈現(xiàn)出上升的趨勢,Simpson指數(shù)的變化趨勢與 Shannon-Wiener指數(shù)的變化趨勢大致相反,整體上呈現(xiàn)出隨海拔梯度下降的趨勢。兩個指數(shù)隨海拔的變化均不呈現(xiàn)出線性的趨勢,且均在海拔1500 m處有較大的波動。

圖2 元謀干熱河谷不同海拔梯度α多樣性變化趨勢Fig. 2 Variation trend of alpha diversity at different Elevation gradients in dry-hot valley of Yuanmou
2.3 β物種多樣性的變化趨勢
元謀干熱河谷不同海拔梯度的β多樣性如圖3所示。在沿海拔梯度上,植物群落SI指數(shù)隨海拔的增加總體上呈現(xiàn)出“M”型變化趨勢。在海拔1400~1500 m之間的相似系數(shù)最大,為0.8。而在海拔 1100~1200、1200~1300、1300~1400 m之間的相似系數(shù)最小,僅為0.4。Cody指數(shù)與SI指數(shù)隨海拔的變化趨勢大致相反,在海拔1400~1500 m之間Cody指數(shù)達到最小值(0.5),且隨海拔的增加總體上呈現(xiàn)出先下降后上升的趨勢。可見,隨著海拔的增加,植物群落間的物種更替速率呈先降低再加快,最終達到最高點的特點。

圖3 元謀干熱河谷不同海拔梯度β多樣性變化趨勢Fig. 3 Variation trend of beta diversity at different elevation gradients in dry-hot valley of Yuanmou
3 討論
本研究聚焦于元謀海拔梯度上干熱河谷植物的物種多樣性。總體上,干熱河谷植物群落結(jié)構(gòu)簡單,物種組成稀少,10個海拔梯度下的河谷群落植物只有4個群落的物種組成大于5種,這與該地區(qū)干旱惡劣的生境條件有關(guān)。本研究中,植被總蓋度與灌木層蓋度總體上呈現(xiàn)出隨著海拔的升高,蓋度增強的趨勢,但兩者的波動范圍不一致,與王飛(2018)關(guān)于白龍江干旱河谷不同坡向主要灌叢群落隨海拔梯度變化的物種多樣性研究研究結(jié)果不一致,造成這種差異的可能原因是元謀干熱河谷地區(qū)隨著海拔的升高,水分越充足(周志瓊等,2008),且灌木層物種趨于穩(wěn)定,以坡柳為主,而木本植物與草本植物兩者的變化相對較大。
植物群落多樣性指數(shù)可以客觀反映群落內(nèi)物種組成特征,在元謀所有海拔梯度下,植物群落 α多樣性指數(shù)偏低,物種豐富度指數(shù)、Shannon-Weiner多樣性指數(shù)和 Simpson優(yōu)勢度指數(shù)最大值分別為15、1.06、0.09,植物群落物種多樣性與干熱河谷區(qū)植被物種多樣性水平相當(dāng)(王飛等,2018;張建利等,2010;楊振寅等,2008)。Shannon-Wiener指數(shù)與Simpson指數(shù)之間的關(guān)系沿海拔呈現(xiàn)出相反的趨勢,Shannon-Wiener指數(shù)與海拔之間表現(xiàn)出較強的正相關(guān)關(guān)系,雖然受水熱條件以及人為干擾等因素的影響出現(xiàn)波動性,但總體呈現(xiàn)出隨著海拔升高,物種多樣性增強的趨勢(表 2,圖 2),與楊陽(2016)關(guān)于三江并流地區(qū)干旱河谷植物物種多樣性海拔梯度格局的比較研究結(jié)果,張大才等(2018)關(guān)于高寒草甸濕潤與干旱化生境中植物多樣性分布與植被的關(guān)系中的研究結(jié)果基本一致。
β多樣性用于反映環(huán)境梯度下植物群落間的差異性,當(dāng)一個群落向另一個群落過渡時,環(huán)境梯度變化較大,不同群落或樣方間共有種越少,β多樣性越大(王慶鎖等,2000;張曉龍等,2017;韓路等,2017)。本研究中,Cody指數(shù)與SI指數(shù)之間的變化趨勢基本相反。隨著海拔的增加,SI相似性系數(shù)在海拔 1400~1500 m 的區(qū)域達到最大值(0.8),而在 1100~1200、1200~1300、1300~1400 m區(qū)域的相似系數(shù)最小,僅為0.4。同時,Cody指數(shù)在海拔1400~1500 m的區(qū)域值最小,為0.5,說明在該海拔區(qū)域的 β多樣性最小,即在海拔1400~1500 m區(qū)域的植物群落結(jié)構(gòu)最穩(wěn)定,物種更替速率最小。
1400~1500 m總蓋度最低,植被稀疏,物種豐富度偏低,推測該海拔是區(qū)域內(nèi)環(huán)境條件最為嚴苛的地段,其穩(wěn)定性亦說明,只有最適于當(dāng)?shù)丨h(huán)境的物種得以生存,其他物種難以進入。所以,這個地段的適生物種可以用于當(dāng)?shù)刂脖浑y恢復(fù)地區(qū)的生態(tài)建設(shè)。與楊銳等(2015)及宋璐璐等(2011)關(guān)于在海拔梯度上各種環(huán)境因子會表現(xiàn)出連續(xù)的梯度性變化的研究結(jié)果不一致,造成這些變化的主要原因可能與該地區(qū)的物種替代有關(guān),即在海拔1400~1500 m處具有邊緣效應(yīng),是干熱河谷植被與常綠闊葉林植被的過渡帶。同時該研究結(jié)果與歐曉昆(1988)關(guān)于海拔1600 m是干熱河谷植被與常綠闊葉林植被的劃分帶的結(jié)論不一致,這可能是由于人為活動(植樹造林、放牧、開墾耕地)的影響而導(dǎo)致過渡帶下移。
4 結(jié)論
研究表明,元謀干熱河谷植物多樣性指數(shù)、植被蓋度與海拔之間均并不呈現(xiàn)連續(xù)的線性的變化趨勢,且都在海拔 1400~1500 m處出現(xiàn)明顯的波動,表明這個區(qū)段是生態(tài)恢復(fù)關(guān)鍵區(qū)段,這里的適生物種最適宜在干熱河谷環(huán)境中生存,是干熱河谷中的建群種。
猜你喜歡
課堂內(nèi)外·初中版(科學(xué)少年)(2025年1期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學(xué)少年)(2025年2期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學(xué)少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學(xué)生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學(xué)苑創(chuàng)造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(shù)(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學(xué)大眾(中學(xué))(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
