不規(guī)則趨化因子在急性肺栓塞后的表達(dá)及機(jī)制研究
2018-11-29 00:26:24謝小娜蔡學(xué)定
重慶醫(yī)學(xué) 2018年32期
關(guān)鍵詞:研究
謝小娜,蔡學(xué)定
(1.浙江中醫(yī)藥大學(xué)附屬溫州中醫(yī)院呼吸內(nèi)科,浙江溫州 325000;2.溫州醫(yī)科大學(xué)附屬第一醫(yī)院呼吸與危重癥醫(yī)學(xué)科,浙江溫州325000)
急性肺栓塞(acute pulmonary embolism,APE)后引起嚴(yán)重的肺損傷和急性肺動(dòng)脈高壓(pulmonary hypertension,PAH)是導(dǎo)致死亡的直接原因。有報(bào)道發(fā)現(xiàn),APE后形成的PAH與肺動(dòng)脈的機(jī)械阻塞作用密切相關(guān)[1],甚至認(rèn)為肺栓塞后血栓的機(jī)械阻塞是導(dǎo)致PAH的直接原因。近年來隨著研究的深入還發(fā)現(xiàn),肺動(dòng)脈機(jī)械阻塞嚴(yán)重程度與肺動(dòng)脈壓力的增高有時(shí)不成正相關(guān),當(dāng)機(jī)械性阻塞基本解除后,仍發(fā)現(xiàn)肺動(dòng)脈壓力持續(xù)升高的現(xiàn)象。此外,有研究結(jié)果顯示,APE時(shí)大量炎癥細(xì)胞聚集浸潤(rùn)在栓塞血管的周圍,這些聚集稍微炎癥細(xì)胞釋放的細(xì)胞因子進(jìn)一步加重肺血管內(nèi)皮損傷,因此,這些細(xì)胞因子在APE性PAH的形成中扮演重要的角色[2]。其中,不規(guī)則趨化因子(fractalkine,F(xiàn)KN)又名趨化因子CX3CLl,是趨化因子CX3C亞族的獨(dú)有成員,具有可溶和膜結(jié)合兩種形式,既有趨化性蛋白的功能也有細(xì)胞黏附分子的功能,還具有促進(jìn)平滑肌細(xì)胞增殖的生長(zhǎng)因子作用[3]。PERROS等[4]在PAH模型中發(fā)現(xiàn),肺動(dòng)脈病變周圍的炎癥細(xì)胞中FKN表達(dá)增加且在血管平滑肌細(xì)胞中的表達(dá)也明顯增加。本文研究大鼠APE后FKN的表達(dá)變化及其機(jī)制,從而探討肺栓塞后炎癥反應(yīng)的作用機(jī)制。
1 材料與方法
1.1材料 (1)動(dòng)物:SPF級(jí)健康雄性SD大鼠72只,體質(zhì)量300~350 g,由溫州醫(yī)科大學(xué)動(dòng)物實(shí)驗(yàn)中心生產(chǎn)提供,實(shí)驗(yàn)動(dòng)物合格證為SCXK(浙)2005-0019。動(dòng)物置于相同環(huán)境和條件飼養(yǎng)。(2)試劑與儀器:BCA 蛋白濃度檢測(cè)試劑盒購(gòu)自Thermo Fisher Scientific公司;DAB顯色試劑盒購(gòu)自福建邁新生物技術(shù)開發(fā)有限公司(MXB Biotechnologies);兔抗大鼠磷酸化p38抗體、兔抗大鼠總P38抗體和山羊抗兔HRP二抗購(gòu)自美國(guó)CST(Cell Signaling Technology)公司;兔抗大鼠FKN抗體購(gòu)自美國(guó)Biolegend公司。
1.2方法
1.2.1實(shí)驗(yàn)動(dòng)物分組 按照完全隨機(jī)的方法將72只SD大鼠分為對(duì)照組、溶劑組、APE組,每組24只;再將每組分為處理后1、4、8 h 3個(gè)亞組,每組8只。
1.2.2APE大鼠模型建立 (1)自體血栓制備:取聚維酮碘棉球?qū)Υ笫箢i部進(jìn)行消毒,內(nèi)徑1.1 mm頭皮針(19G)置入左頸動(dòng)脈取血并靜置10 min凝血,再將血塊置于37 ℃水中20 min,切成統(tǒng)一規(guī)格1.1 mm×3.0 mm的栓子,并用無菌生理鹽水輕柔沖洗3 遍,放入對(duì)應(yīng)試管內(nèi),加無菌生理鹽水2 mL,4 ℃冷藏備用,所有操作均為無菌操作。(2) APE模型的建立:使用5%水合氯醛(3 mg/kg)經(jīng)腹腔注射麻醉大鼠,暴露頸正中線并行縱行切口,分離大鼠頸外靜脈并剪一小口,插入19G聚乙烯導(dǎo)管,通過導(dǎo)管注入之前準(zhǔn)備的栓子約50個(gè)(不致大鼠死亡),附以無菌生理鹽水0.6 mL(0.1 mL/min),血栓伴隨循環(huán)血液匯聚并嵌頓于肺動(dòng)脈,行核素掃描檢測(cè)發(fā)現(xiàn)栓塞部位核素?cái)z取量明顯減少,證明APE大鼠模型建立成功。對(duì)照組不作任何處理;溶劑組大鼠除注入等量的生理鹽水外,余操作同APE組。在成功建立APE模型后,分別于1、4、8 h 3個(gè)時(shí)間點(diǎn)處死大鼠,經(jīng)胸腔取大鼠肺組織。
1.2.3免疫組織化學(xué)(IHC)檢測(cè) 摘取大鼠左肺上葉組織,固定于4%多聚甲醛液24 h,經(jīng)漂洗、脫水、透明、浸蠟、和包埋后再行切片,通過IHC檢測(cè)FKN、p38蛋白的表達(dá)。組織切片經(jīng)脫蠟、水化、封閉及微波修復(fù)后,滴加抗FKN抗體(1∶50)或抗p38抗體(1∶50)于4 ℃過夜后,加入山羊抗兔二抗(1∶50),DAB顯色終止后行蘇木素復(fù)染,最終脫水、透明中性樹膠封片。置于顯微鏡下觀察p38、FKN在肺組織中的表達(dá)。
1.2.4Western blot檢測(cè)p38絲裂原活化蛋白激酶(p38 mitogen-activated protein kinases,p38MAPK)表達(dá) 稱取各組大鼠等量新鮮肺組織加入細(xì)胞裂解液后充分研磨,再經(jīng)超聲細(xì)胞粉碎儀充分破碎,于低溫超速離心機(jī)12 000 r/min 4 ℃離心 10 min,取上清液加入等體積2×上樣緩沖液及相應(yīng)體積PBS充分震蕩混勻并分裝至PCR管經(jīng)PCR儀維持99 ℃運(yùn)行5 min,多余蛋白于-70 ℃冰箱保存?zhèn)溆谩_M(jìn)行10 mol/L SDS-PAGE(電壓濃縮膠80 V,分離膠120 V)蛋白電泳,電泳后將蛋白轉(zhuǎn)印至PVDF膜(轉(zhuǎn)膜電流300 mA,30 min)后用5 g/L 脫脂奶粉在室溫下封閉1 h,分別加入5%牛血清清蛋白(BSA)稀釋的磷酸化p38單克隆抗體(1∶1 000,兔抗大鼠)和總p38單克隆抗體(1∶1 000,兔抗大鼠) 4 ℃冰箱孵育過夜后,用TBST充分洗滌條帶3 min后,室溫下孵育標(biāo)記辣根過氧化物酶(HRP)的羊抗兔二抗(1∶2 000)1 h,再次用TBST充分洗滌3 min,使用ECL試劑顯色及暗室內(nèi)膠片曝光。使用圖像分析系統(tǒng)對(duì)所得條帶進(jìn)行掃描保存及灰度分析。
1.2.5RT-PCR檢測(cè)FKN mRNA表達(dá) 使用Trizol試劑盒提取肺組織總RNA并測(cè)出A值,A260 nm/A280 nm為1.8~2.0。用日本TakaRa 公司RT-PCR 試劑盒進(jìn)行逆轉(zhuǎn)錄擴(kuò)增,逆轉(zhuǎn)錄擴(kuò)增條件:65 ℃ 1 min,30 ℃ 5 min,65 ℃ 15 min,98 ℃ 5 min,5 ℃ 5 min,轉(zhuǎn)錄成cDNA后,各組取5 μL加入PCR反應(yīng)體系,PCR擴(kuò)增條件:94 ℃預(yù)變性5 min,94 ℃ 1 min,56.2 ℃退火1 min,72 ℃延伸1 min,循環(huán)30次。各組PCR產(chǎn)物等量注入2 %瓊脂糖凝膠孔中行水平電泳并在紫外凝膠成像系統(tǒng)下觀察、攝像,隨后進(jìn)行吸光度值(OD)分析。各樣本的基因表達(dá)量計(jì)算方法為產(chǎn)物膠上條帶OD與β-action基因條帶OD的比值。

2 結(jié) 果
2.1APE大鼠FKN的表達(dá)部位 IHC檢測(cè)結(jié)果顯示,F(xiàn)KN在APE大鼠的肺實(shí)質(zhì)、肺動(dòng)脈血管壁上均有不同程度的表達(dá);p38在APE大鼠的肺實(shí)質(zhì)、肺動(dòng)脈血管壁細(xì)胞核內(nèi)均有表達(dá)。

表1 各組大鼠各時(shí)間點(diǎn)磷酸化P38蛋白OD比值比較
a:P<0.05,與APE組比較
2.2各組大鼠各時(shí)間點(diǎn)肺組織磷酸化p38OD比值比較 APE組各時(shí)間點(diǎn)大鼠肺組織磷酸化p38OD比值高于對(duì)照組和溶劑組(P<0.05),見表1、圖1。

圖1 各組大鼠處理后8 h磷酸化p38的表達(dá)水平
2.3各組大鼠各時(shí)間點(diǎn)肺組織FKN mRNAOD比值比較 APE組各時(shí)間點(diǎn)大鼠肺組織FKN mRNAOD比值明顯高于對(duì)照組和溶劑組(P<0.05),見表2。
表2 各組大鼠各時(shí)間點(diǎn)FKN mRNA的OD比值比較
a:P<0.05,與APE組比較
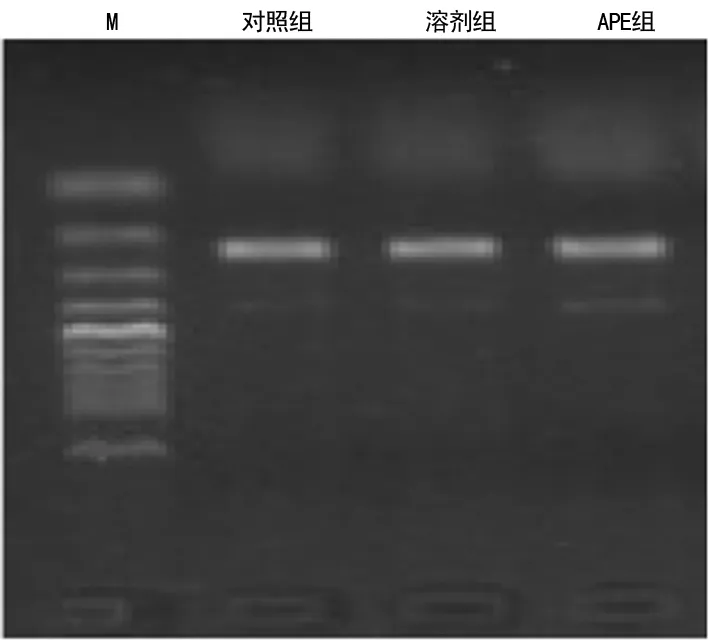
圖2 各組大鼠處理后8 h肺組織FKN mRNA的表達(dá)水平
3 討 論
3.1FKN在APE后的表達(dá)及其作用 FKN是1997 年由BAZAN等[5]在用趨化因子的基因探針檢索表達(dá)序列標(biāo)簽數(shù)據(jù)庫(kù)時(shí)發(fā)現(xiàn)的,是目前CX3C-類趨化因子中惟一成員,它是含有多個(gè)結(jié)構(gòu)域的大分子蛋白,人的FKN基因定位在染色體16q13[6]。膜結(jié)合型Fractalkine 存在于活化的細(xì)胞表面,主要起著黏附作用;分泌型Fractalkine主要是對(duì)單核、NK細(xì)胞等具有強(qiáng)有力的趨化作用[7-8]。近年來有報(bào)道指出,炎癥反應(yīng)時(shí),F(xiàn)KN在單核細(xì)胞和T細(xì)胞募集到血管壁的過程中起重要作用,而且FKN還具有類似于生長(zhǎng)因子促肺動(dòng)脈平滑肌細(xì)胞增殖的作用[3]。在APE后,對(duì)FKN的研究較少報(bào)道。
本研究結(jié)果顯示,APE后FKN、p38在肺血管內(nèi)皮、肺泡上皮、支氣管內(nèi)皮上有明顯表達(dá)(P<0.05)。本研究結(jié)果顯示,APE組各時(shí)間點(diǎn)大鼠肺組織FKN mRNA的表達(dá)量明顯高于對(duì)照組和溶劑組(P<0.05)。目前研究認(rèn)為,APE后PAH的形成可能與栓塞后血管周圍大量炎癥細(xì)胞浸潤(rùn)及其釋放的細(xì)胞因子進(jìn)一步損傷肺動(dòng)脈內(nèi)皮有關(guān)[9],而表達(dá)增多的FKN對(duì)炎癥細(xì)胞的聚集起到促進(jìn)作用。
3.2p38MAPK通路在APE后被激活從而上調(diào)FKN的表達(dá) 1993年科學(xué)家研究發(fā)現(xiàn)p38MAPK信號(hào)轉(zhuǎn)導(dǎo)通路,該通路一旦被激活,細(xì)胞質(zhì)中的非磷酸化p38MAPK轉(zhuǎn)化為磷酸化p38MAPK,再移位到細(xì)胞核內(nèi)[10]。本研究中采用Western blot檢測(cè)肺組織磷酸化p38的表達(dá),結(jié)果顯示隨時(shí)間延長(zhǎng)磷酸化p38的表達(dá)有明顯增高趨勢(shì)(P<0.05),提示在大鼠APE后,p38MAPK通路被激活。因此本課題組推測(cè),APE后p38MAPK通路被激活,上調(diào)了FKN的表達(dá),使得FKN在APE后明顯升高。
此外,有研究報(bào)道FKN在PAH的發(fā)生、發(fā)展中起到重要作用,值得注意的是損傷的內(nèi)皮細(xì)胞可過度表達(dá)FKN[11-12],其是否在APE后PAH的形成中起作用有待進(jìn)一步研究。也有研究顯示FKN 所引起的趨化白細(xì)胞、纖維化、凋亡等均可能是心力衰竭的誘因[13-15]。右心功能不全也是APE后PAH的形成的原因之一。
綜上所述,本研究結(jié)果顯示在APE后8 h內(nèi),大鼠肺組織中FKN的表達(dá)即明顯升高,作為炎癥趨化因子,F(xiàn)KN對(duì)APE后肺部炎癥反應(yīng)有明顯促進(jìn)作用,從而加重了APE后炎癥反應(yīng)及PAH的形成。
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國(guó)語大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學(xué)報(bào)(2017年2期)2017-07-05 08:13:02
國(guó)際商務(wù)財(cái)會(huì)(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
