長江口九段沙濕地海三棱藨草生物量分配特征及其影響因子
2018-11-14 12:38:54許宇田童春富
生態(tài)學報 2018年19期
許宇田,童春富
華東師范大學河口海岸學國家重點實驗室,上海 200062
克隆植物又稱無性系植物,它們將資源分配于各構件時會進行權衡,從而使生長和繁殖的效益達到最佳[1- 3]。這種資源分配通過植物生物量可以體現(xiàn)出來。生物量是植物的基本生物學特征和功能性狀,是物質(zhì)和能量積累的基本體現(xiàn);生物量分配是決定其資源獲取、生長競爭以及繁殖能力的重要因素,對植物生態(tài)策略的形成至關重要[4- 5]。濕地是水陸交替的過渡帶,是地球上重要的生態(tài)系統(tǒng)類型之一。生活于其中的濕地植物相當一部分具有克隆生長特征。它們通過功能性狀的可塑性來調(diào)節(jié)資源配置,選擇不同的生物量分配模式以適應復雜多變的環(huán)境,從而最大效率地利用資源來提高其生存適合度和與競爭能力[6]。濕地克隆植物生物量分配已成為濕地生態(tài)學研究中的一大關鍵性問題[7- 8]。
海三棱藨草(Scirpusmariqueter)為我國特有種,主要分布在長江口和杭州灣一帶的潮間帶鹽沼濕地[9]。它是莎草科藨草屬的一種多年生耐鹽草本植物,生長期為4—11月,花期為5—6月;可進行有性繁殖或營養(yǎng)繁殖,常形成單優(yōu)植物群落;具有促淤保灘、凈化水質(zhì),以及生物棲息地等多種生態(tài)功能[10- 12]。目前針對海三棱藨草的研究已涉及種群物候與分布格局、繁殖生態(tài)學、種間競爭能力、生態(tài)價值與功能、生物量動態(tài)與分配、植物生理等諸多領域[10,13- 22]。海三棱藨草生物量分配的相關研究表明,不同環(huán)境和干擾條件下海三棱藨草在生物量分配上表現(xiàn)出很強的生態(tài)適應性,密度等形態(tài)參數(shù)和高程梯度、沉積物有機碳含量等環(huán)境指標在海三棱藨草的生物量分配中扮演著非常重要的角色[15- 16,18,23- 24]。已有的研究分別在這兩個方面進行了有益的探索,為進一步開展相關研究提供了良好的基礎。本文參考已有研究,在現(xiàn)場取樣調(diào)研的基礎上,以海三棱藨草生物量分配特征為切入點,綜合植物形態(tài)特征與環(huán)境因子,展開系統(tǒng)分析,探究影響海三棱藨草生物量分配的主要因子及其作用機理。研究結果可以為海三棱藨草的培育與恢復,鹽沼濕地生態(tài)系統(tǒng)的保護與合理利用提供科學依據(jù)。
1 材料與方法
1.1 研究區(qū)域
研究區(qū)域位于長江口九段沙濕地國家級自然保護區(qū)。九段沙為長江河口新生沙洲,位于南北槽之間,主要由江亞南沙、上沙、中沙、下沙及鄰近淺水水域組成。九段沙土壤發(fā)育過程短,潮灘沉積物以淤泥和粉砂為主。該區(qū)域屬于亞熱帶季風性氣候區(qū),年降水量約1143 mm,年平均氣溫為15.7℃[25]。附近水域潮汐屬于非正規(guī)半日淺海潮,平均潮差2.67 m,最大潮差4.62 m;冬季鹽度被高鹽度咸水控制,約為3‰—15‰,夏季主要受淡水控制,約為1‰—6‰;在長江來水來沙與海潮的共同作用下,區(qū)域水動力條件復雜[26]。近年來,九段沙濕地處在長江口高強度開發(fā)的大環(huán)境中,在自然干擾和人為干擾下,平面擴張減慢,水下灘淤漲受到抑制,中高灘淤漲速度加快,呈現(xiàn)“長高不長大”的變化特點[27]。沙洲整體上北淤南沖,淤積區(qū)域集中在上、中、下沙中高灘區(qū)、江亞南沙與上沙沙尾,以及靠近北槽深水航道南導堤兩側;沖刷區(qū)域主要為九段沙南側區(qū)域[28- 29]。沙洲的沖淤動態(tài)與鹽沼濕地植物群落的發(fā)育與演替密切相關。
九段沙鹽沼植被具有明顯的成帶分布特征,植物種類少,群落結構簡單,常形成單優(yōu)群落,在群落交錯帶出現(xiàn)分布范圍很窄的混生群落。原有自然植被優(yōu)勢種為蘆葦(Phragmitesaustralis)、海三棱藨草和藨草(S.triqueter)。1997年在中沙區(qū)域引種了互花米草(Spartinaalterniflora),目前已擴散至中沙和下沙大部分區(qū)域,形成了大面積的單優(yōu)群落。海三棱藨草主要分布在植被帶前緣,為重要的先鋒物種,受波浪與潮汐作用較大。在嚴重沖刷岸段基本沒有海三棱藨草分布,往往出現(xiàn)侵蝕陡坎,從光灘直接過渡到蘆葦、互花米草分布區(qū)。
1.2 研究方法
1.2.1 樣品采集
海三棱藨草的生物量分配隨季節(jié)變化[15],但其在 10 月份左右開始停止生長、結籽,并開始衰老,生物量達到最大[17],地下生物量在植株開始衰老后相對穩(wěn)定[14],可以進行生物量分配的分析與研究。因此作者于2016年10月在九段沙開展了現(xiàn)場取樣調(diào)研工作。海三棱藨草群落是該區(qū)域的特征植被,根據(jù)文獻資料以及現(xiàn)場觀測,遵循典型性、代表性和一致性的原則,分別在九段沙各沙洲選擇群落發(fā)育典型、空間分布均勻、原生狀態(tài)完好的研究區(qū), 利用GPS確定采樣站點。采樣站點設置如圖1所示。J1—J3 為3個取樣站點,分別為江亞南沙(J1)、上沙(J2)、中下沙(J3)。主要采樣區(qū)基本為淤漲區(qū)。
每站點設置5塊樣地,每樣地設置 2個植被取樣樣方,樣方面積為50 cm×50 cm。現(xiàn)場記錄各樣方海三棱藨草的蓋度、株數(shù)、株高,齊地刈割采集植物地上部分。在每個樣方內(nèi),設置25 cm×25 cm小樣方,用鐵鏟向下挖掘至約25—30 cm,確保將樣方內(nèi)植株地下部分全部挖出。挖出的泥樣用0.5 mm孔徑網(wǎng)篩淘洗,得到海三棱藨草地下部分。
沉積物樣品與植物樣品采集同步進行。在每個植物樣方內(nèi),使用內(nèi)徑3.5 cm的柱狀取樣器,采集深度30 cm的沉積物柱樣,現(xiàn)場分割成6段5 cm長的泥柱。現(xiàn)場用便攜式儀器測定記錄沉積物氧化還原電位、pH、溫度,孔隙水的鹽度等指標。每個指標在每個樣方中重復測定2次。

圖1 九段沙采樣站點圖Fig.1 Location of sampling sites in Jiuduansha Shoals
1.2.2 樣品處理
(1)海三棱藨草
在實驗室內(nèi),將植物樣品用自來水沖洗多遍后,再用Mill-Q 水沖洗。按照莖葉、果實分離地上部分,并記錄開花株數(shù)、結籽數(shù);按照須根、根莖、球莖、莖基(莖葉埋在土面以下部分)分離地下部分,并測量記錄莖基長與根莖長。分離后的各部分在60℃下烘干至恒重,稱量得到生物量干重,精確至0.01 g。
(2)沉積物
沉積物柱樣在實驗室稱取濕重后,于60℃烘干至恒重,稱取干重,精確至0.01 g,用以計算含水率、容積密度。烘干的土壤樣品分別用馬爾文激光粒度儀(MS2000)測粒度,用元素分析儀(Vario EL III)測定總碳、總氮含量。
1.2.3 數(shù)據(jù)處理
計算植株密度、平均株高和莖基(截留莖)高度,將其作為海三棱藨草整體表形參數(shù)。計算得到根莖、果實、球莖各部分生物量比例分別表征克隆生殖投資、有性生殖投資以及生長投資;計算果實與根莖重之比、果實與球莖重之比以及根莖與球莖重之比來表征有性和無性繁殖之間的對比關系[15]。
采用統(tǒng)計軟件包IBM SPSS Statistics 22進行數(shù)據(jù)處理。采用單因素方差分析(one-way ANOVA)對不同站點間生境因子、海三棱藨草的相關參數(shù)進行顯著性分析,在此基礎上,以boferroni檢驗方法進一步做組間兩兩比較,分析不同站點兩兩之間的差異。以P<0.05表示顯著差異,P<0.01表示極顯著差異。采用統(tǒng)計軟件Primer 5.2.8對海三棱藨草生物量分配特征與環(huán)境變量進行非線性回歸分析。用BVSTEP程序找出與海三棱藨草生物量分配特征匹配最好的生境因子組合,并計算相關系數(shù);再用RELATE程序檢驗兩者之間的相關性是否顯著。所有數(shù)據(jù)在使用時需經(jīng)過平方根變換,以滿足數(shù)據(jù)的正態(tài)性要求;生境因子的相關數(shù)據(jù)需要進行標準化處理,以避免量綱差異造成的影響[30]。
2 結果與分析
2.1 海三棱藨草表形特征

圖2 不同站點海三棱藨草表形特征(平均值±標準誤)Fig.2 Morphological characteristics of S. mariqueter in the different sites (Mean ± SE)上方標注不同小寫字母的柱子表示相互之間存在顯著差異(P<0.05)
不同站點海三棱藨草表形特征如圖2所示。單因素方差分析結果表明,植株密度和植株高度在不同站點間存在顯著差異(P<0.05),而莖基高度并未發(fā)現(xiàn)顯著不同(P>0.05)。不同站點兩兩比較的結果表明,植株密度在J3站點顯著高于J2站點(P<0.05),J1介于兩者之間,與J2、J3均無顯著差異(P>0.05);植株高度在J1站點顯著高于J3站點(P<0.05),J2介于J1、J3之間,與兩者差異均不顯著(P>0.05);莖基高度在各站點兩兩之間均不存在顯著差異(P>0.05)。
2.2 海三棱藨草生物量分配特征
不同站點海三棱藨草各部分的生物量分配如圖3所示。單因素方差分析結果表明,海三棱藨草的莖葉、果實、須根、根莖生物量所占比例在不同站點間存在顯著差異(P<0.05),而球莖生物量所占比例并未發(fā)現(xiàn)顯著不同(P>0.05)。不同站點兩兩比較的結果表明,莖葉生物量所占的比例在J2站點顯著高于J3站點(P<0.05),J1介于兩者之間,與J2、J3均無顯著差異(P>0.05);果實生物量所占的比例在J1站點顯著高于J2和J3站點(P<0.05),J2和J3之間差異并不顯著(P>0.05);須根生物量所占的比例在J3站點顯著高于J2站點(P<0.05),J1介于兩者之間,與J2、J3均無顯著差異(P>0.05);根莖生物量所占的比例在J3站點顯著高于J2站點(P<0.05),J1介于兩者之間,與J2、J3均無顯著差異(P>0.05);球莖生物量所占比例在不同站點兩兩之間并未發(fā)現(xiàn)顯著不同(P>0.05)。
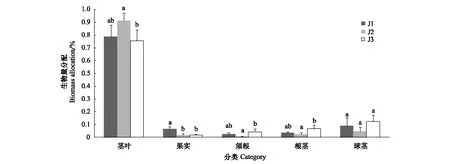
圖3 不同站點海三棱藨草生物量分配特征(平均值±標準誤)Fig.3 Biomass allocation characteristics of S. mariqueter in the different sites (Mean± SE)上方標注不同小寫字母的柱狀圖表示同一類別生物量在不同站點間存在顯著差異(P<0.05);J1—J3 為3個取樣站點,其中,J1:江亞南沙,Jiuduansha Shoals 1;J2:上沙,Jiuduansha Shoals 2;J3:下沙,Jiuduansha Shoals 3
不同站點海三棱藨草繁殖與生長之間的對比關系如圖4所示。單因素方差分析結果表明,果實與根莖之比、果實與球莖之比在不同站點間差異顯著(P<0.05),而根莖與球莖之比在不同站點間并未發(fā)現(xiàn)顯著差異(P>0.05)。不同站點兩兩比較的結果表明,果實與根莖之比在J1站點顯著高于J2和J3站點(P<0.05),J2和J3之間并無顯著差異(P>0.05);果實與球莖之比在J1站點顯著高于J3站點(P<0.05),J2介于兩者之間,與J1、J3均無顯著差異(P>0.05)。

圖4 不同站點海三棱藨草繁殖與生長特征(平均值±標準誤) Fig.4 Characteristics of reproduction and growth of S.mariqueter in the different sites(Mean± SE)上方標注不同小寫字母的柱狀圖表示同一類別生物量比率在不同站點間存在顯著差異(P<0.05);FRR:果實/根莖比率,Ratio of fruit to rhizome biomass;FCR:果實/球莖比率,Ratio of fruit to corm biomass;RCR:根莖/球莖比率,Ratio of rhizome to corm biomass
海三棱藨草不同參數(shù)相關特征如表1所示。植株密度與根莖、須根生物量比例呈極顯著正相關(P<0.01),而與莖基高度顯著負相關(P<0.05)。莖基高度與根莖生物量比例呈極顯著負相關(P<0.01),與須根生物量比例顯著負相關(P<0.05),與莖葉生物量比例顯著正相關(P<0.05)。莖葉生物量比例與球莖、根莖、須根生物量比例均呈現(xiàn)極顯著負相關關系(P<0.01)。須根生物量比例與球莖、根莖生物量比例呈現(xiàn)極顯著正相關關系(P<0.01);同時球莖與根莖生物量比例也呈現(xiàn)極顯著正相關關系(P<0.01)。
2.3 主要生境因子特征
不同站點沉積物理化性質(zhì)如圖5所示。單因素方差分析結果表明,不同站點的孔隙水鹽度、沉積物電導率、沉積物的容積密度(5—10、25—30 cm土層)、總碳含量(10—15、20—25、25—30 cm土層)、總氮含量(5—10 cm土層)存在顯著差異(P<0.05),而pH、氧化還原電位以及同一深度的含水率、其他深度的柱段的容積密度以及總碳總氮含量在不同站點間并未發(fā)現(xiàn)顯著不同(P>0.05)。不同站點兩兩比較的結果表明,沉積物電導率在J1站點顯著低于J2和J3(P<0.05),J2和J3兩者之間并無顯著差異(P>0.05);孔隙水鹽度在J3站點顯著高于J1和J2站點(P<0.05),J2低于J1,但兩者之間并無顯著差異(P>0.05)。深度5—10 cm沉積物容積密度在J2站點顯著高于J3站點(P<0.05),J1介于兩者之間,與J2、J3均無顯著差異(P>0.05);深度25—30 cm沉積物容積密度在J2站點顯著低于J1和J3站點(P<0.05),而J1和J3則不存在顯著差異(P>0.05)。10—15 cm的碳百分含量在J1站點顯著高于J3站點(P<0.05),J2介于兩者之間,與J1、J3均無顯著差異(P>0.05);20—25 cm的碳百分含量在J2站點顯著高于J3站點(P<0.05),J1介于兩者之間,與J2、J3均無顯著差異(P>0.05);25—30 cm的碳百分含量在J3站點顯著低于J1和J2站點(P<0.05),J1低于J2,但是兩者之間無顯著差異(P>0.05)。5—10 cm的氮百分含量在J1站點顯著高于J2和J3站點(P<0.05),J2和J3之間差異不顯著(P>0.05)。

表1 海三棱藨草特征參數(shù)自相關矩陣
**,P<0.01;*,P<0.05
2.4 海三棱藨草生物量分配主要影響因子
不同站點海三棱藨草生物量分配特征與沉積物理化指標、海三棱藨草表形參數(shù)非線性回歸分析結果如表2所示。海三棱藨草的果實與根莖的生物量之比和5—10、25—30 cm總碳含量,0—5、5—10 cm總氮含量,0—5 cm容積密度,莖基高度等6種因子的組合極顯著相關(P<0.01);而其他生物量分配指標與本文涉及到的環(huán)境因子、植物表形參數(shù)均不存在顯著相關性(P>0.05)。
3 討論
3.1 海三棱藨草自身生長特征與生物量分配
已有研究表明,具有克隆生長特征的植物的生物量分配受植株高度和密度等自身生長特征的影響[3,31- 34]。植物種內(nèi)存在對有限空間與資源的競爭,生物量分配格局是決定植物資源獲取、競爭以及繁殖能力的重要因素[35]。植物自身生長情況如密度變化等會影響種內(nèi)競爭強度,使得植物生物量分配發(fā)生變化[20,34,36]。克隆植物生物量分配普遍支持最優(yōu)化理論,即通過對器官間生物量分配的權衡,對資源競爭作出反應,以保證植物能最大化利用受限資源,應對不斷變化的環(huán)境條件[6,32,37]。


圖5 不同站點沉積物理化性質(zhì)Fig.5 Physical and chemical characteristics of the sediment in the different sites圖中標注不同小寫字母表示不同站點間的顯著性差異(P<0.05)

特征Characteristics相關性系數(shù)Correlation篩選因子組合Selection combinationP果實/球莖比Fruit/corm biomass ratio0.0946,8,14,19,220.212果實/根莖比Fruit/rhizome biomass ratio0.3417,11—13,24,320.006根莖/球莖比Rhizome/corm biomass ratio0.1722,50.124根莖生物量比例Rhizome biomass alloca-tion0.1296,11—14,16,21,320.151果實生物量比例Fruit biomass allocation-0.0155,12,310.443莖葉生物量比例Shoot biomass allocation0.1721,8,14,16,22,24,26,300.087球莖生物量比例Corm biomass allocation-0.1572,4,8—10,12,15,19,26,300.99須根生物量比例Root biomass allocation0.1653,6,7,11—13,18,20,21,23,26—300.138
因子: 1: 沉積物孔隙水鹽度;2: 沉積物電導率;3: 沉積物溫度;4: 沉積物pH;5: 沉積物氧化還原電位;6:0—5 cm沉積物總碳含量;7: 5—10 cm沉積物總碳含量;8: 10—15 cm沉積物總碳含量;9: 15—20 cm沉積物總碳含量;10: 20—25 cm沉積物總碳含量;11: 25—30 cm沉積物總碳含量;12: 0—5 cm沉積物總氮含量;13: 5—10 cm沉積物總氮含量;14: 10—15 cm沉積物總氮含量;15: 15—20 cm沉積物總氮含量;16: 20—25 cm沉積物總氮含量;17: 25—30 cm沉積物總氮含量;18: 0—5 cm沉積物含水率;19: 5—10 cm沉積物含水率;20: 10—15 cm沉積物含水率;21: 15—20 cm沉積物含水率;22: 20—25 cm沉積物含水率;23: 25—30 cm沉積物含水率;24: 0—5 cm沉積物容積密度;25: 5—10 cm沉積物容積密度;26: 10—15 cm沉積物容積密度;27: 15—20 cm沉積物容積密度;28: 20—25 cm沉積物容積密度;29: 25—30 cm沉積物容積密度;30:海三棱藨草植株密度;31:海三棱藨草株高;32: 海三棱藨草莖基高度
目前針對海三棱藨草生物量分配的研究中,有結果顯示隨著植株高度增加,海三棱藨草的營養(yǎng)投資減少,而分配更多的資源給有性繁殖[15]。但是本次研究并未發(fā)現(xiàn)海三棱藨草植株高度與各部分生物量間存在顯著相關性。同時,本次研究顯示,植株密度和根莖以及須根生物量比例呈極顯著正相關,這在已有研究的基礎上進一步證明了密度是影響克隆植物生物量分配的一個重要因子[20,36]。根莖生物量比例可以反映出植物對無性繁殖的投資,因此由本文研究結果可知,隨密度的增加,海三棱藨草對無性繁殖的投資增加,這與之前的部分研究結果相一致[18,38]。但目前關于繁殖投資對密度的響應的研究,也存在不同的結論,如有研究表明在生物因素(種間、種內(nèi)競爭等)的干擾下,海三棱藨草主要采取有性繁殖的策略來維持適合度[15,39],因為種子傳播可使植物避開不利場所,而增加對無性繁殖的資源分配可能會增加其種間競爭強度[1]。之所以出現(xiàn)這種情況,可能與本文研究中海三棱藨草的地下器官所需資源受限的程度有關。增加無性繁殖投資這一生物量分配模式可以被認為是克隆植物的保守策略。由于海三棱藨草對土壤營養(yǎng)的需求較低[40],并且通過無性系分株比通過種子形成幼苗的方式成活率更高,無性繁殖形成聚集的群體利用不同小生境中克隆分株的形態(tài)可塑性和克隆生理整合作用[41- 42],可以最大限度地吸收和利用環(huán)境資源,緩解環(huán)境中資源不均或干擾脅迫對生長和繁殖帶來的不利影響,提高個體的生存和競爭力。海三棱藨草生物量分配符合最優(yōu)化理論,即隨著密度增加,生存空間受到限制,種內(nèi)競爭加劇,海三棱藨草通過權衡生物量分配來實現(xiàn)對礦質(zhì)營養(yǎng)和水分等資源與環(huán)境的最大化利用,提高對生境的適合度。
3.2 環(huán)境因子對海三棱藨草生物量分配的影響
根據(jù)已有研究,除自身生長因子之外,具有克隆生長特性的鹽沼植物生物量分配還受到環(huán)境因子的影響[43- 45]。如氮素可通過改變氮在葉片中的分配格局影響葉片的光合作用,并通過改變碳水化合物的庫源關系和能量消耗水平而調(diào)節(jié)碳同化物質(zhì)在體內(nèi)的分配,從而影響其生長和生物量分配[46- 49];土壤的含水率、透氣性等對克隆植物的生物量積累與分配等均有顯著影響[50- 52];溫度變化通過直接改變植物的光合能力、生長速率,間接改變土壤含水率、營養(yǎng)物質(zhì)的可利用性,影響植物生長以及生物量分配[53- 54]。不同區(qū)域的環(huán)境因子往往存在明顯、甚至顯著的差異,這可能是導致海三棱藨草具有不同的生物量分配特征的關鍵所在。本文研究結果顯示,特定深度的沉積物總碳、總氮含量、容積密度以及莖基高度等因子組合與部分海三棱藨草生物量分配指標呈極顯著相關關系。結合已有研究,進一步證實了土壤營養(yǎng)元素(碳氮含量、總磷含量等)對海三棱藨草生物量及生物量分配存在重要作用[20,23,39,55]。盡管有控制實驗顯示海三棱藨草須根、莖葉和果實生物量分配受到鹽度和水位的顯著影響[56],然而本文研究中并未發(fā)現(xiàn)海三棱藨草生物量分配與沉積物鹽度[57- 59]、含水率[40]以及溫度[60]等環(huán)境因子存在顯著相關性。這可能是由于環(huán)境因子的變化在實驗控制條件下與自然狀態(tài)下存在差異,并且環(huán)境因子間還存在著相互作用,可能與高程[15,20,24]、潮汐水動力[61- 62]等其他因子的影響有關。未來需要涵蓋更多的環(huán)境因子,進行綜合分析。
本文研究中的莖基高度可以反映出環(huán)境中泥沙堆積深度。莖基高度越大,植物根莖埋藏越深,海三棱藨草受泥沙堆積的影響越大。從本文研究結果來看,莖基高度不僅是非線性回歸篩選出的與果實根莖之比相關的因子之一,而且與根莖生物量比例呈極顯著負相關,與植株密度、須根生物量比例顯著負相關,與莖葉生物量分配顯著正相關。與之前對鹽沼克隆植物短尖苔草(Carexbrevicuspis)的研究相比[63],沉積物堆積產(chǎn)生的影響在植株密度變化上較為一致,對其他指標的影響卻明顯不同。根據(jù)本文研究結果推測,隨著沉積物堆積加厚,沉積壓力可能會對植株地下部分,包括根尖分生組織形成物理脅迫,不利于地下器官的生長發(fā)育,海三棱藨草會傾向于將更多的資源分配到莖葉,為有性繁殖提供條件,減少無性繁殖投資比例。這與已有研究認為,在生物量分配的權衡中,克隆植物在非生物因素的壓力下更傾向于采取無性繁殖的方式[39]的觀點存在明顯不同,即如果非生物因素直接影響或限制了無性繁殖,克隆植物同樣會選擇有性繁殖策略。
綜上所述,海三棱藨草生物量分配特征是其自身生長特征與環(huán)境因子綜合作用的結果。從本文研究結果來看,除了本文涉及的環(huán)境因子外,可能還存在其他因子也會對海三棱藨草的生物量及分配特征產(chǎn)生影響。在今后的研究中需要拓展生境因子涵蓋范圍,從而進一步系統(tǒng)、綜合研究多因子對于海三棱藨草生物量分配特征的影響。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
鐵道通信信號(2020年9期)2020-02-06 09:15:22
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
數(shù)學大王·趣味邏輯(2019年5期)2019-06-13 20:27:43
小學科學(學生版)(2019年5期)2019-05-21 01:00:18
經(jīng)濟技術協(xié)作信息(2018年30期)2018-11-22 06:20:24
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
