鯉春病毒血癥病毒的研究進展
2018-11-06 02:37:10林婧楠趙景壯盧彤巖徐黎明
水產學雜志 2018年5期
關鍵詞:檢測
林婧楠,趙景壯,盧彤巖,徐黎明
(1.中國水產科學研究院黑龍江水產研究所,黑龍江 哈爾濱 150070;2.上海海洋大學水產與生命學院,上海 201306)
鯉春病毒血癥病毒屬單股負鏈RNA病毒目,彈狀病毒科Rhabdoriridae,暫定為水泡性病毒屬Vesiculorius,被類脂囊膜包裹的彈狀病毒[1]。鯉春病毒血癥的潛伏期較短,沒有典型的外部臨床癥狀,容易誤診。目前尚無有效控制該病的方法,一旦出現疑似該病的癥狀,應及早確診,隔離感染和帶毒的病魚,截斷水平傳播途徑及捕殺來控制。
1 病原生物學特征
CSVV病毒粒子在15%~60%的蔗糖溶液中浮力密度為1.16g/mL[2,3],在5%~25%的蔗糖梯度中沉降系數為38~40S[4]。外界環境的改變影響病毒的感染性,對酸(在pH3時30min,侵染率1%)、堿(在pH11時,侵染率50%~70%,pH7~10時最為穩定)、熱(56℃,30min)、脂溶劑以及乙醚最為敏感,其他化學試劑,如2%NaOH、500mg/L含氯消毒劑、0.01%碘,在特定濃度和作用時間可使病毒失活,可用于養殖消毒。血清可以保護病毒的侵染力,實驗室常用2%血清培養液保護病毒。含有血清時,經過4次反復凍融,侵染率在90%左右,而缺乏血清時,侵染率僅為5%左右。冷凍真空干燥可長期保存病毒[5]。
2 基因結構特征
CSVV粒子為典型的彈狀結構,一端圓弧形,另一端平坦,長90~180nm,寬60~90nm[5-7]。其基因組全長為 11 019bp[8],由非翻譯前導(Leader)區、基因區、基因間連接區及非轉錄拖尾(Trailer)區4個主要部分構成(圖1)。
基因前導區有69bp,前19bp可能為啟動子,其中50~56bp(GAAAAAT)為前導RNA轉錄終止信號,60~64bp(AACAG)為N基因轉錄起始信號。
基因區共包含5個開放閱讀框(ORFs),編碼5種結構蛋白:核蛋白(Nuclear protein,N)、磷蛋白(Phosphoprotein,P)、基質蛋白(Matrixprotein,M)、糖蛋白(Glycoprotein,G)和 RNA 聚合酶(RNA polymerase,L),3’端到 5’端的排列順序為 N-P-M-G-L。
核蛋白(N)在病毒粒子中含量最多,主要與RNA緊密結合構成直徑約50nm、呈螺旋對稱的核衣殼。該結構蛋白在轉錄調節中起重要的作用[6]。
磷蛋白(P)為核衣殼的組成部分,呈酸性,純化后SVCV的P蛋白在SDS-PAGE中分子量約50kD,高于推算的結果(35.5kD),其原因可能是P蛋白N端(45~88)存在一個大的負電荷結構域[8]。
基質蛋白(M)由223個氨基酸組成,呈磷酸化、高度堿性,構成囊膜內表面。糖蛋白通過M蛋白與螺旋狀核衣殼相連,共同實現病毒粒子的裝配、出芽的啟動功能。
糖蛋白(G)由509個氨基酸構成,病毒粒子的囊膜表面有長釘狀的糖蛋白突起。成熟的糖蛋白為三聚體囊膜粒子,由3部分構成:胞外結構域、跨膜結構域和胞漿結構域,是典型的跨膜蛋白,能選擇性地識別宿主細胞受體,促使二者的膜融合,介導病毒內吞,使宿主細胞感染致病;在免疫系統下,識別病原,誘導宿主產生中和抗體。糖蛋白含有與免疫相關的抗原決定簇,對血清學特征起決定性作用,是當前DNA疫苗設計研究的熱點。2002年至2004年,劉葒等[9]從養殖的鯉科魚類和觀賞魚中分離出8株CSVV,測定了其G蛋白編碼的基因序列,發現我國的這8個分離株與USA株、980451株、980528株和970469株在系統發育樹上的進化方向一致。根據Stone等[10]對各地分離出的部分CSVV的G蛋白基因序列以及Basic等[11]1994年至2007年收集的22株G蛋白部分核酸序列的分析和系統發育樹比較,將CSVV的基因型分成Ia、Ib、Ic和Id 4個亞組,Ia和Id的變異度較高,又將有明顯差別的3個Id基因組毒株劃分為Id1、Id2及Id3。CSVV中的一個血清型能與狗魚苗彈狀病毒(Pike fry rhabdovirus,PFRV)產生交叉反應。前期研究發現,中國CSVV的分離株的基因型為Ia[12]。
RNA聚合酶(L)是核衣殼的組成成分,可與N、P蛋白共同組成病毒內核衣殼的RNA蛋白復合物,完成病毒的轉錄和復制。
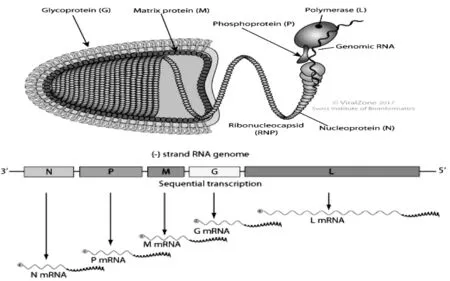
圖1 鯉春病毒血癥病毒(carp spring virernia virus,CSVV)病毒結構、基因組結構以及連接順序(引自http://viralzone.expasy.org/4657)Fig.1 Structure,genomic and linkage of carp spring virernia virus(CSVV)
在基因間連接區的G-L基因中存在4bp(CTAT)保守序列,其余基因間存在2bp(CT)保守序列[13]。該調控區存在于水泡病毒屬中;G-L基因間不存在類似粒彈狀病毒屬編碼的非結構蛋白(nonstructural protein,NVprotein)基因。該蛋白在病毒復制過程中起重要作用,這兩點均為區分粒彈狀病毒屬和水泡性病毒屬的重要標志。
拖尾區有49bp,其中TATG(A)7為L基因轉錄終止信號,末端的8個核苷酸序列(GTCTTCGT)與前導區末端(ACGAAGAC)呈反向互補,是彈狀病毒基因組及不分段負鏈RNA病毒的共同特征[8]。
與同屬的其他彈狀病毒相比,CSVV的蛋白序列中L蛋白間的同一性最高,與VSV的L蛋白氨基酸序列同一性達53.6%[14];與水皰性口炎病毒(Vesicular Stomatitis Virus,VSV)的M蛋白同一性達28%,在羧基末端的同一性較低[15]。
3 分類特征
迄今已報道的魚類彈狀病毒(Fish rhabdovirus)有十幾種,鯉春病毒血癥病毒(CSVV)是其中的一種,此外還包括病毒性出血敗血癥病毒(Viral haemorrhagic septicemia virus,VHSV)、傳染性造血器官壞死病毒 (Infectious hematopoietic necrosis virus,IHNV)、日本牙鲆彈狀病毒(Hirame rhabdovirus,HIRRV)、狗魚苗彈狀病毒、烏鱧彈狀病毒(Snakehead rhabdovirus,SHRV)、彈狀病毒 903/87(Rhabdovirus 903/87,903/87)、鱖彈狀病毒(Mandarin fish Siniperca chuatsi rhabdovirus,SCRV)、胭脂魚彈狀病毒(Chinese sucker rhabdovirus,CSRV)及新近分離出的海鮭彈狀病毒28/97(Sea trout rhabdovirus 28/97,STRV28/97)等[16-19]。
彈狀病毒科包括水泡性病毒屬(Vesiculovirus)、狂犬病毒屬(Lyssavirus)、暫時熱病毒屬(Ephemerovirus)和粒彈狀病毒屬(諾拉彈狀病毒屬,Novirhabdovirus)。其中粒彈狀病毒屬是國際病毒分類委員會第7次報告上設立的一個新屬。根據是否含有編碼非結構蛋白的基因,將傳染性造血器官壞死病毒、病毒性出血敗血癥病毒(viral haemorrhagic septicemia virus,VHSV)、鱧彈狀病毒、牙鲆彈狀病毒歸為粒彈狀病毒屬。而鯉春病毒血癥病毒和狗魚苗彈狀病毒暫定為水泡性病毒。
4 流行情況
鯉春病毒血癥病毒感染的魚類較為廣泛,主要感染鯉科魚類,如錦鯉Cyprinus carpio、草魚Cten opharyngodm idellus、鯽 Carassius auratus、鰱 Hypophthalmichthys molitrix、鳙 Aristichthys nobilis、金魚 Carassius auratus、擬鯉 Rutilus rutilus、圓腹雅羅魚Leuciscus idus和丁鯛Tinca tinca等。此外,自然感染的魚類還有六須鲇Silurus glanis[20]。一齡以上的鯉最易感染患病。在中世紀,該病在歐洲開始流行,對歐洲各國鯉科魚類養殖業造成了巨大的經濟損失[21],而后向美洲和亞洲等地傳播。我國于2003年首次在天津養殖場的觀賞魚中分離出SVCV[22]。目前,該病已成為全球性魚類的疾病。
該病毒可在13~22℃下生長,最適為17℃。因此,SVC常在水溫15~20℃的春季爆發,17℃左右發病率高,水溫超過20℃有所下降,超過22℃不再發病。
染毒魚、病魚和病死魚是主要的傳染源,以水平傳播為主,垂直傳播次之。水平傳播包括直接接觸傳播和間接傳播,間接傳播包括非生物媒介(水);生物媒介(水生吸血寄生蟲)以及污物傳播。病毒隨附著污物(糞便、尿液、皮膚黏液和性腺分泌液等)排出體外,4~10℃時水中和泥中仍具有感染性,可保持4~6周[5,23],可通過鰓進入魚體內,在血液中保持11周左右。
5 臨床癥狀及病理變化
5.1 臨床癥狀
行為變化:早期病魚呼吸困難,需氧量高,常聚集在池塘進水口附近,食欲減退,行動遲緩,對應激的反應遲鈍,身體平衡力降低;后期病魚萎靡,身體傾斜,常臥于池底[21,24],嚴重時停止游動。
臨床癥狀:病魚體表發黑、眼球突起、鰓絲蒼白、皮膚表面和肌肉有出血點、腹部腫脹、內有積水,肛門處拖有一細長、黏稠、糞便樣的腸黏膜炎癥產物與排泄物的混合產物,常稱之為“假糞”,但不屬于該病的特征性癥狀。
5.2 病理變化
染毒魚最終因體內水鹽平衡失調致死。剖檢病魚發現,主要以全身性出血、水腫為主要特征癥狀。病毒通常在毛細血管內皮細胞、造血組織和腎元細胞等部位增殖。在感染期,肝、腎、脾、鰓、腦髓病毒含量較高,常作為檢驗組織,發病3周后,含量迅速下降消失。血液中病毒滴度不高,但持續時間長。肝臟實質組織出現多個病灶壞死、充血,可見黃疸現象;胰臟組織出現炎癥以及大量壞死病灶;脾臟充血,網狀內皮大量增生;腸道嚴重發炎,出現卡他性腸炎。
6 檢測技術
CSVV病毒可在鯉上皮瘤細胞(EPC)、草魚性腺細胞(CO)、鳙尾柄細胞株(FHM)、藍鰓太陽魚成纖維細胞(Lepomis macrochirus,BF-2)和虹鱒性腺細胞(RTG-2)等魚類細胞株上增殖,造成細胞病變效應(Cytopathogenic effect,CPE)。病變細胞內出現顆粒,H&E染色可見胞質萎縮空泡化,核膜增厚,顯微鏡下可見脫落的細胞變亮[20]。FHM和EPC細胞最敏感,為OIE推薦的分離病毒的細胞。EPC和FHM的最適溫度為20~25℃和20~22℃,產生的最大感染性為109TCID50/mL。該病毒除了在魚類細胞中增殖,還能在禽類、哺乳類和兩棲類動物細胞內增殖。
6.1 傳統的檢測方法及免疫學方法
目前,國內外常采用細胞培養法分離、檢測CSVV病毒,再通過中和試驗確定。該方法雖然準確率高,但檢測大約需要2周左右,耗時費力。
在細胞培養分離病毒的基礎上,可采用免疫學的方法,如酶聯免疫吸附檢驗(Enzyme-linked immunosorbent assay,ELISA)進行鑒定、間接熒光抗體試 驗 (Indirect fluorescent antibody test,IFAT),但CSVV的囊膜來自宿主的細胞膜,易產生非特異性的抗體,因而病毒的抗血清效價不高,影響檢測結果[25]。CSVV在血清學上區別于其他彈狀病毒,采用ELISA可能會與其他魚類彈狀病毒如VHSV、IHNV等發生交叉反應,還與PFRV有共同的抗原決定簇,在ELISA和間接熒光(IF)中有明顯的交叉反應[26,27]。高隆英等[26]根據CSVV糖蛋白基因序列設計逆轉錄-聚合酶鏈式反應(RT-PCR)和半嵌套PCR(semi-nested-PCR)的引物 F1、R2、R4,其中 F1和R2可擴增出CSVV核酸中的714bp片段,F1和R4可擴增出606bp片段。特異性檢測結果表明,與VHSV、IHNV和PFRV無交叉反應,具有良好的特異性,可在42h內得到結果。
6.2 OIE檢測標準
根據OIE公布的《水產動物檢測手冊》,目前我國出入境檢驗檢疫行業標準SN/T1152-2002以及我國規定的GB/T 15805.5-2008魚類檢疫方法,常規檢測CSVV方法是通過細胞培養分離病毒后,采用RT-PCR方法進行診斷。該過程需經過兩代的細胞培養,時間長,檢測效率低。王姝等[28]在國家行業標準的基礎上,有機結合細胞培養法,利用酶聯免疫吸附試驗ELISA進行初步的篩選,再用RT-PCR復查,即實現高通量檢測,整個用時又比國家標準用時縮短了一半。
6.3 核酸探針檢測方法
Svetlana F等[29]根據CSVV-S30株的M和G基因的核酸序列設計引物,構建生物素標記的核酸探針,應用RT-PCR和斑點雜交技術成功在感染CSVV的細胞培養液和魚組織中檢測出病原體,其中在腦組織中最易檢測出該病毒。利用生物素標記的核酸探針方法,既實現了快速檢測,又提高了其靈敏度。
6.4 熒光RT-PCR檢測方法
張利峰等[30]根據鯉春病毒血癥病毒的核酸保守區序列,建立了快速檢測CSVV的熒光RT-PCR檢測技術,僅需4h就可以檢測出10-6病毒稀釋液,不僅試驗周期短、靈敏,還實現了對病毒模版RNA的定量。安偉等[31]根據CSVV的核蛋白的保守序列,成功建立了SYBR Green I熒光定量RT-PCR檢測技術,病毒檢測下限100.76TCID50/mL,靈敏度高于常規PCR 100倍。
6.5 數字RT-PCR檢測方法
吳斌等[32]利用“油包水”的形式,對RT-PCR的擴增反應進行計數,建立了數字RT-PCR的技術。與實時定量RT-PCR相比,在兩者均能檢測病毒的104稀釋倍數基礎上,擁有更高的靈敏度。
6.6 間接ELISA檢測方法
郭闖等[33]以超速離心法純化CSVV,免疫大白兔,制備了兔抗CSVV免疫血清,同時利用間接ELISA方法快速檢測CSVV,以兔抗SVCV血清1∶300稀釋后包被96孔酶標板,滴度篩查CSVV陽性毒株,結果與RT-PCR鑒定結果相同。整個過程需要1 d,具有周期短、準確的優點。
6.7 新型液相芯片檢測技術
尹偉力等[34]根據GenBank中CSVV的G基因序列,找出25對保守堿基作為檢測探針,設計引物并進行生物素標記,通過液相芯片儀器檢測偶聯熒光編碼微球與CSVV的RT-PCR產物雜交反應產生的熒光信號,發現液相芯片檢測體系在鑒別CSVV上具有高特異性和靈敏度,最低檢出量為10 pg。
6.8 環介導等溫擴增技術(Loop-mediated isothermalamplification,LAMP)
Shivappa等[35]對美國卡羅萊納州分離的CSVV菌株的G蛋白基因序列,設計出4套引物,分別進行LAMP和RT-LAMP擴增,證實建立的RT-LAMP對CSVV的檢測具有特異性,與RT-PCR的靈敏度相似,最低檢測量達101TCID50/mL。
陳進會等[36]依照G基因保守序列,設計3套LAMP引物,通過恒溫實時熒光檢測系統ESE-Quant tube scanner,第1套和第3套有峰值,進而引入環引物。結果顯示,第1套LAMP引物的檢測穩定性、靈敏度和特異性效果最佳,能夠實現高效檢測CSVV。因此,實時熒光LAMP法具有極佳的靈敏度和快速檢驗的優勢,適用于基層檢驗,檢測限度可達到每反應2.4 pg。
7 防治方法
目前對于該病沒有特效藥和有效的控制方法,只能通過化學、物理消毒的方式預防該病。一旦發現該病,應及時中斷傳染源,隔離或撲殺病魚和帶毒魚,防止大規模疫病的爆發[31]。在20世紀80年代,歐洲出現了含CSVV和PFRV的二階滅活疫苗,能夠誘導鯉科魚類產生高效的中和抗體。隨后,成功開發了CSVV弱毒疫苗,在養殖漁業中有良好的應用前景。給體質量40~100g的鯉腹腔注射6.3×104TCID50后,免疫保護率在90%以上[37]。我國的CSVV疫苗研制還處于探索階段,主要的研究領域集中在減毒疫苗、亞單位疫苗及核酸疫苗等。
傳統滅活疫苗、減毒疫苗具有毒力返強的缺點[25],需要添加佐劑,存在一定的安全隱患,應用效果不穩定。
DNA疫苗是90年代的最新一代疫苗,無需像傳統的滅活疫苗添加佐劑,卻仍具備傳統疫苗的優點。DNA疫苗構型更接近天然構象,潛在致病性低,適于工廠化集中生產,相對安全,具有一定的應用價值[38]。CSVV疫苗的研發有一定的進展,Kanellos等[39]以CSVV G蛋白基因,設計了構建穩定性較好的DNA疫苗。為評估已構建的4組CSVV糖蛋白DNA疫苗的免疫效果,歐陽征亮等[40]通過肌肉注射的免疫途徑誘導機體產生免疫。結果顯示:第一組重組質粒DNA編碼全長的G蛋白疫苗保護率為14.47%;第二組重組質粒DNA編碼去C端信號肽的G蛋白疫苗保護率為22.22%;第三組重組質粒DNA編碼去N端跨膜區的G蛋白疫苗保護率為17.32%;重組質粒DNA編碼同時去C端信號肽和N端跨膜區的G蛋白疫苗保護率為17.32%。Emmenegger等[41]的鯉攻毒試驗驗證,已構建的CSVV的G蛋白DNA疫苗的保護效率為50%~88%,能夠有效降低感染鯉的死亡率。
8 展望
鯉春病毒血癥是一種高致病性、高死亡率的鯉科魚類傳染性疾病。目前,對該病毒的疫苗研制尚處于探索階段。盡管SVC的DNA疫苗已取得一定的進步,但是在DNA疫苗的效率優化、安全性以及疫苗的免疫途徑上仍然存在一些問題。同時,DNA疫苗的作用及代謝機制還有待進一步探索[37]。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
