貝類殼-體質(zhì)量比和靜水沉降特性的相關(guān)性
2018-11-02 06:36:46汪學(xué)杰顧黨恩牟希東楊葉欣CesarDemayo胡隱昌
生態(tài)學(xué)報(bào) 2018年18期
羅 渡,汪學(xué)杰,徐 猛,顧黨恩,牟希東,韋 慧,楊葉欣,Cesar G. Demayo,胡隱昌,*
1 農(nóng)業(yè)農(nóng)村部休閑漁業(yè)重點(diǎn)實(shí)驗(yàn)室,廣東省現(xiàn)代休閑漁業(yè)工程技術(shù)研究中心,中國(guó)水產(chǎn)科學(xué)研究院珠江水產(chǎn)研究所,廣州 510380 2 Department of Biological Sciences, College of Science and Mathematics, Mindanao State University-Iligan Institute of Technology, Iligan City, Philippines
貝類物種豐富、利害兼有,其數(shù)量、分布與人類生產(chǎn)、生活密切相關(guān)[1]。由于貝類的主動(dòng)運(yùn)動(dòng)主要依靠肉足驅(qū)動(dòng),其種群擴(kuò)散在行為上更多地表現(xiàn)為被動(dòng)擴(kuò)散。物種擴(kuò)散的媒介或障礙往往能對(duì)種群、進(jìn)化或生態(tài)系統(tǒng)產(chǎn)生廣泛的效應(yīng)[2]。不僅是水生貝類,即使陸生螺類,水依然是其存活的重要條件,水流是其遠(yuǎn)距離遷移的主要媒介之一[3]。研究表明水流介導(dǎo)了釘螺的遷移,并最終導(dǎo)致血吸蟲(chóng)病的蔓延[4]。福壽螺、非洲大蝸牛等惡性入侵生物的滲透式擴(kuò)散和爆發(fā)成災(zāi)也往往隨洪水而發(fā)生。因此水介質(zhì)在貝類的物種傳播上起著重要的作用。貝類在水下的沉降規(guī)律是研究其運(yùn)動(dòng)規(guī)律的重要內(nèi)容之一[5]。
在貝類學(xué)的研究初期,貝殼特征幾乎成了分類特征的唯一來(lái)源[6]。長(zhǎng)期以來(lái),貝殼形態(tài)特征都是分類的主要依據(jù)[7],但是貝殼形態(tài)也會(huì)隨著環(huán)境變化而變化,貝殼的細(xì)微結(jié)構(gòu)在種的水平上也具有差異[8]。貝類具有較大的表型可塑性,即使是不同地理種群,生活的環(huán)境差異以及長(zhǎng)期的適應(yīng)性進(jìn)化在形態(tài)上都可能產(chǎn)生明顯分化[9]。盡管貝殼幾何學(xué)引起學(xué)者的注意已經(jīng)有200多年了,然而對(duì)構(gòu)造規(guī)則如何發(fā)揮作用以及形態(tài)進(jìn)化是如何發(fā)生的還知之甚少。運(yùn)動(dòng)需要能量,貝殼在發(fā)揮著保護(hù)和支持作用的同時(shí)也是一種運(yùn)動(dòng)負(fù)擔(dān)。有殼動(dòng)物在爬行、游泳或掘穴時(shí),肌肉所施加的力量都必須足以克服貝殼的重量。碳酸鈣的密度比水的大三倍左右,減少貝殼重量將增加其個(gè)體的機(jī)動(dòng)性能[10]。
貝類在貝殼厚度、生活史策略等特征上都存在快速進(jìn)化的能力[11]。運(yùn)動(dòng)行為在軟體動(dòng)物的生態(tài)和進(jìn)化上具有重要作用,長(zhǎng)期的適應(yīng)性選擇往往可以使其運(yùn)動(dòng)擴(kuò)散能力出現(xiàn)特化[12]。貝殼是軟體動(dòng)物的支撐和保護(hù)器官,但是貝殼質(zhì)量的增加不利于其捕食和遷徙。此外,在進(jìn)化過(guò)程中,為了適應(yīng)不同的生境,貝殼會(huì)通過(guò)異速生長(zhǎng)產(chǎn)生差異,但是此種差異在“殼-體質(zhì)量比”上的體現(xiàn)以及此種變化對(duì)于運(yùn)動(dòng)擴(kuò)散功能的影響未見(jiàn)相關(guān)報(bào)道。生態(tài)水力學(xué)是研究生命體在水中的擴(kuò)散轉(zhuǎn)移規(guī)律及流場(chǎng)控制技術(shù)的新興交叉型學(xué)科[4]。因此本研究選取9種具代表性的常見(jiàn)貝類作為研究對(duì)象,運(yùn)用形態(tài)學(xué)與生態(tài)水力學(xué)相結(jié)合的交叉學(xué)科方法,通過(guò)分析殼-體質(zhì)量比和靜水沉降特性的種間差異,及殼-體質(zhì)量比與靜水沉降速度、靜水沉降加速度之間的相關(guān)關(guān)系,以期探討貝殼形態(tài)的適應(yīng)性進(jìn)化與水環(huán)境介導(dǎo)的運(yùn)動(dòng)擴(kuò)散能力進(jìn)化的關(guān)系,為闡明貝類種群擴(kuò)散的生態(tài)策略提供理論基礎(chǔ)。
1 材料與方法
1.1 實(shí)驗(yàn)材料
實(shí)驗(yàn)于珠江水產(chǎn)研究所外來(lái)水生生物入侵風(fēng)險(xiǎn)評(píng)估中心進(jìn)行,所用貝類為南方地區(qū)常見(jiàn)貝類,樣品主要采集于廣東省境內(nèi),每個(gè)物種的樣本量≥30。所有實(shí)驗(yàn)貝類樣品皆為成體,9種貝類樣品分屬于9個(gè)屬,其物種分類信息如表1所示。
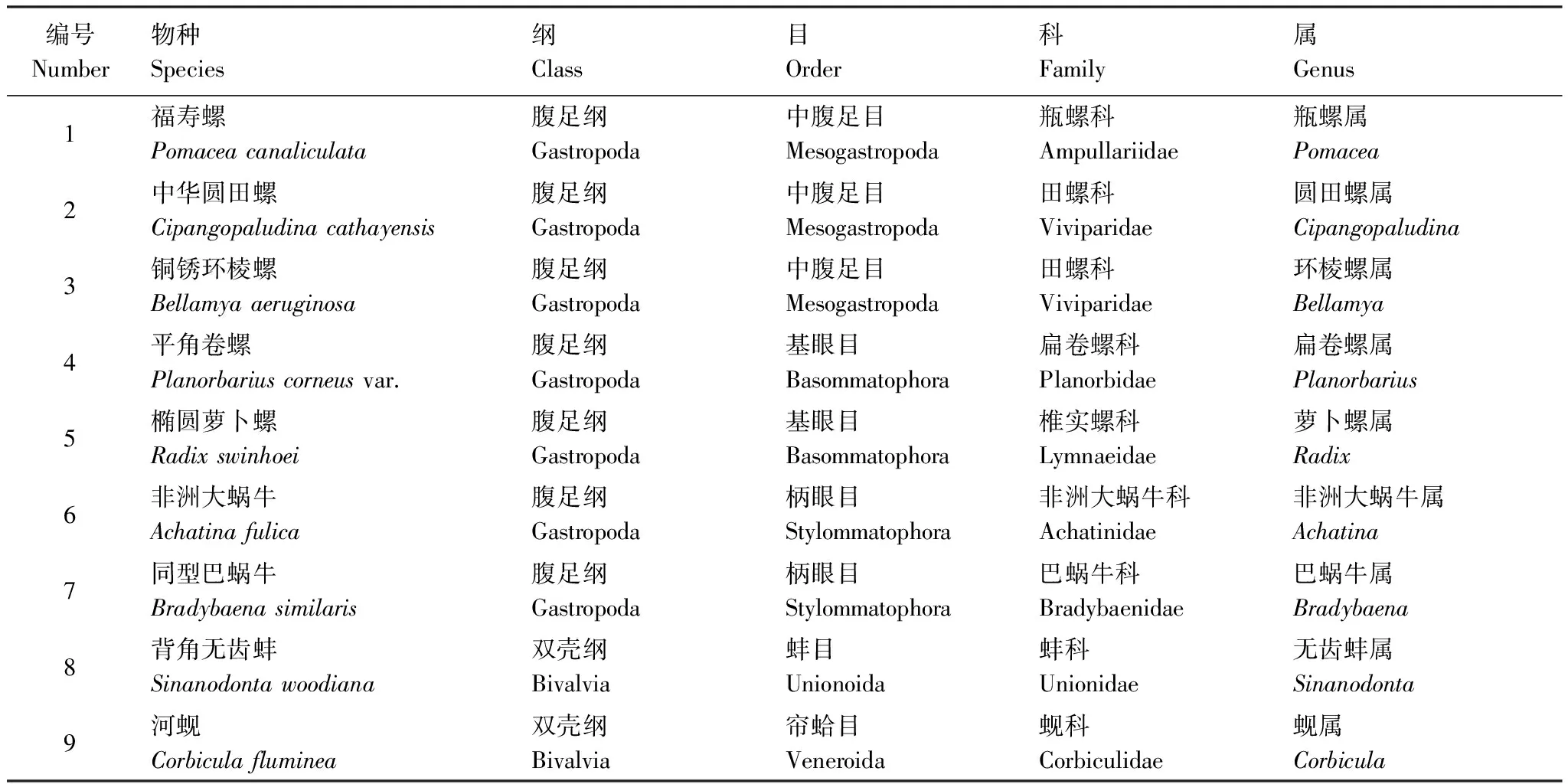
表1 實(shí)驗(yàn)所用貝類的分類信息
1.2 研究方法
(1)形態(tài)測(cè)量:貝類個(gè)體的形態(tài)參數(shù)用游標(biāo)卡尺(精確到0.001 cm)進(jìn)行測(cè)量,所測(cè)指標(biāo)包括:殼長(zhǎng),殼頂沿中軸到殼底部的最大長(zhǎng)度;殼寬,垂直于殼長(zhǎng)中軸的最大寬度[13-14];對(duì)于雙殼貝類,殼長(zhǎng)為殼體前后方向最大距離;殼寬為殼體背部與腹部方向最大距離[15]。用吸水紙吸干表面水分后,用電子天平稱量體質(zhì)量(精確到0.01 g)和殼質(zhì)量(精確到0.01 g),所有質(zhì)量均為濕重[16]。用變異系數(shù)(Coefficient of Variation,C.V.)表征數(shù)據(jù)的離散程度大小,其值為標(biāo)準(zhǔn)差與平均值之比。
(2)殼-體質(zhì)量比計(jì)算:按以下公式計(jì)算。

(3)靜水沉降特性的測(cè)定:采用靜水沉降法,設(shè)置沉降缸模擬自然條件下的沉降特征。沉降缸為圓柱體形,高2.4 m、直徑1.2 m,以清澈的池塘養(yǎng)殖用水灌裝。于水面中心點(diǎn)以下15 cm設(shè)置沉降起始點(diǎn),于起始點(diǎn)以下140 cm的垂直平面設(shè)置沉降計(jì)時(shí)終點(diǎn),終點(diǎn)距離缸底垂直距離大于25 cm,缸底設(shè)置“緩沖網(wǎng)”以使樣品免受機(jī)械刺激和損傷。所有受試個(gè)體皆以體長(zhǎng)軸平行于水面的姿態(tài)進(jìn)入自然沉降。以秒表記錄起點(diǎn)至終點(diǎn)的運(yùn)動(dòng)時(shí)間,測(cè)量重復(fù)3次(圖1)。依據(jù)公式v=S/t計(jì)算平均沉降速度(其中,v為沉降速度,S為沉降距離、t為沉降時(shí)間);假設(shè)樣品為勻加速運(yùn)動(dòng),依據(jù)公式a=2S/t2計(jì)算加速度(其中a為沉降加速度,S為沉降距離、t為沉降時(shí)間)。
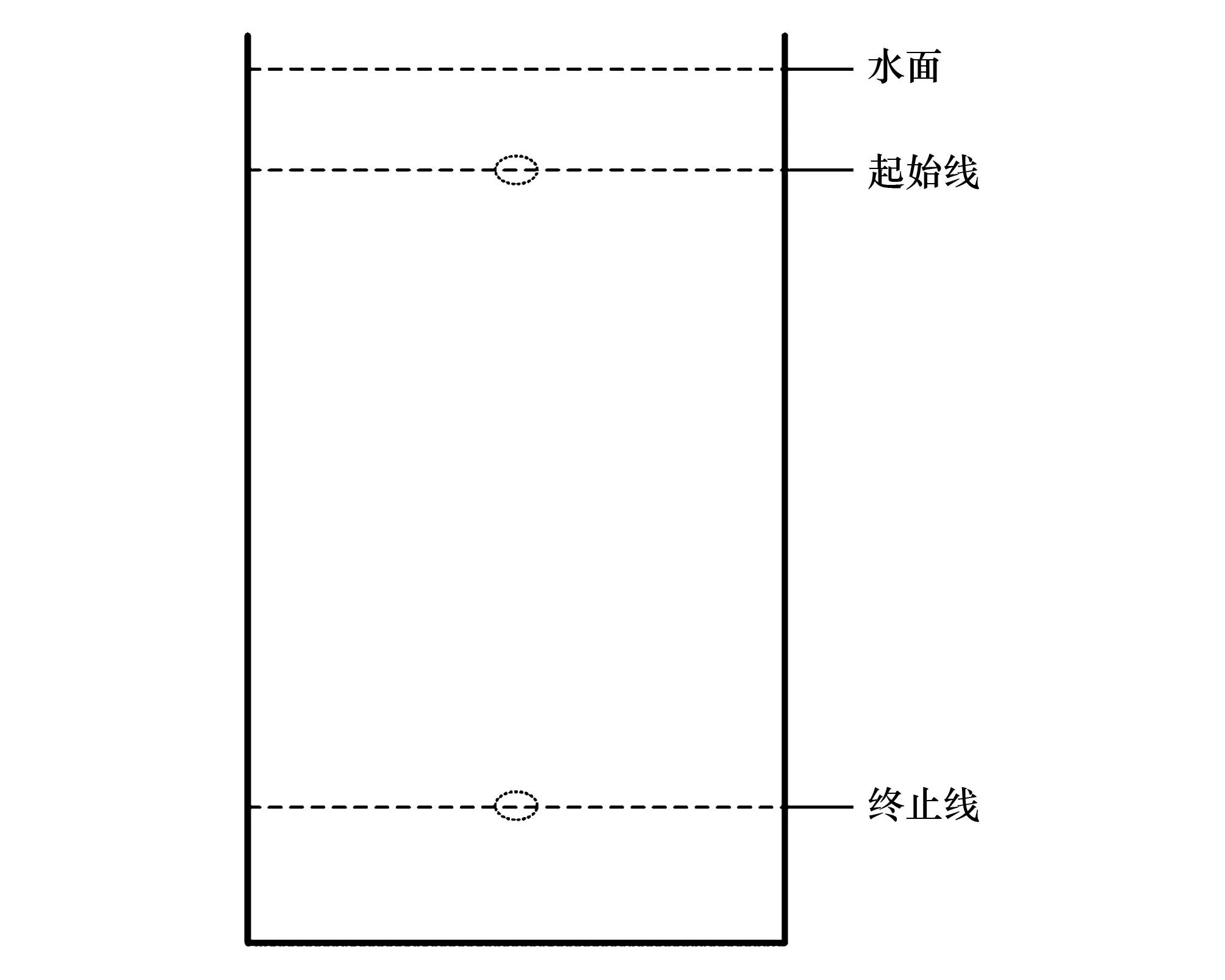
圖1 靜水沉降示意圖Fig.1 Settlement process of the molluscs in this study
1.3 數(shù)據(jù)分析
采用Microsoft office 2010軟件、OriginPro 8.6軟件和IBM SPSS Statistics 20軟件等進(jìn)行數(shù)據(jù)統(tǒng)計(jì)、差異性比較、回歸分析和作圖。依據(jù)前述公式計(jì)算殼-體質(zhì)量比和沉降速度、沉降加速度。結(jié)果表示為:平均值±標(biāo)準(zhǔn)差。以“殼長(zhǎng)”、“殼寬”、“體質(zhì)量”、“殼-體質(zhì)量比”等作為因變量,采用單因素方差分析法(ANOVA)分析種間差異顯著性(P<0.01)。
2 結(jié)果與分析
2.1 供試貝類的主要形態(tài)測(cè)量
分別測(cè)量9種貝類的殼長(zhǎng)、殼寬和體質(zhì)量三個(gè)主要的形態(tài)指標(biāo),其測(cè)量結(jié)果如表2所示。由表可知,背角無(wú)齒蚌(SinanodontawoodianaLea, 1834)的殼長(zhǎng)、殼寬、體質(zhì)量最大,平角卷螺(Planorbariuscorneusvar.)的殼長(zhǎng)最小,橢圓蘿卜螺(RadixswinhoeiH.Adams)的殼寬和體質(zhì)量最小。9種貝類的殼長(zhǎng)、殼寬、體質(zhì)量的總體變異系數(shù)(C.V.)分別為0.67,0.54,1.11,因此體質(zhì)量的變異>殼長(zhǎng)的變異>殼寬的變異。
2.2 殼-體質(zhì)量比的種間差異
研究結(jié)果顯示(圖2):9種貝類的平均殼-體質(zhì)量比為(0.32±0.13),所有受試貝類殼-體質(zhì)量比的種內(nèi)C.V.<0.36,平均C.V.=0.20,背角無(wú)齒蚌變異最大(C.V.=0.35),河蜆變異最小(C.V.=0.11),因此貝類殼-體質(zhì)量比的種屬特異性強(qiáng)。不同種貝類之間存在明顯的種間變異,其中非洲大蝸牛的殼-體質(zhì)量比最小(0.17±0.04),極顯著地低于其他貝類(P<0.01),河蜆的殼-體質(zhì)量比最大(0.54±0.06),極顯著高于其他貝類(P<0.01),福壽螺的殼-體質(zhì)量比接近0.50(0.50±0.06),銅銹環(huán)棱螺次之(0.38±0.06)。中華園田螺(0.22±0.03)、背角無(wú)齒蚌(0.23±0.08)、同型巴蝸牛(0.26±0.08)、平角卷螺(0.26±0.04)和橢圓蘿卜螺(0.28±0.04)的殼體質(zhì)量比為0.25左右,種間差異性較小(P<0.01),只有中華園田螺和橢圓蘿卜螺之間存在極顯著差異(P<0.01),其余差異不顯著。陸生貝類的總體殼-體質(zhì)量比(0.21)顯著小于水生貝類(0.35)(P<0.01),但并非所有陸生貝類的殼-體質(zhì)量比都較水生貝類小。
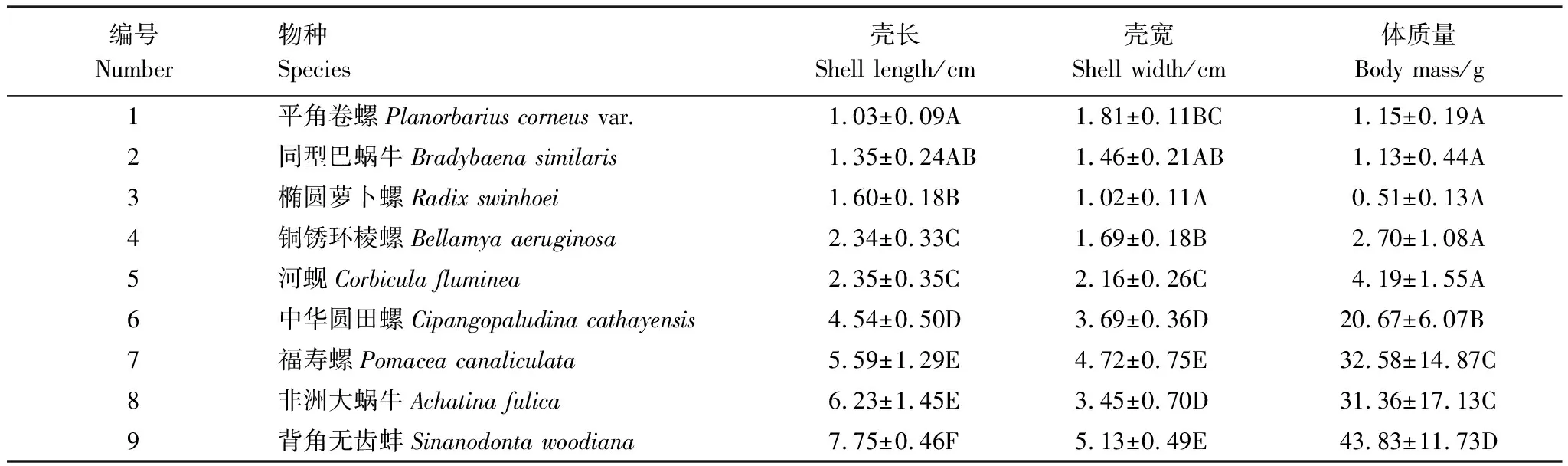
表2 實(shí)驗(yàn)貝類的主要形態(tài)測(cè)量
同列數(shù)據(jù)無(wú)相同字母表示在0.01水平上差異極顯著
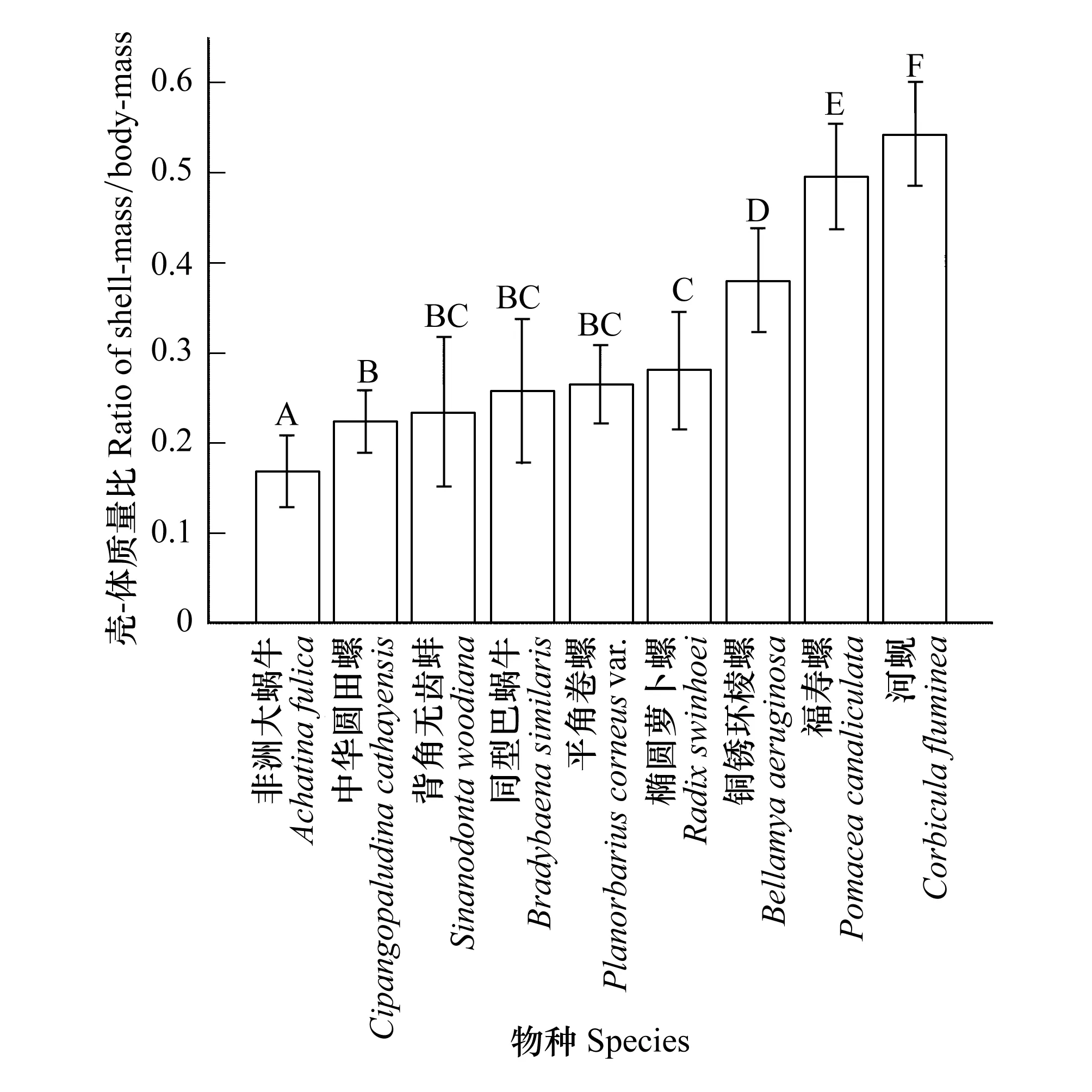
圖2 貝類殼-體質(zhì)量比的種間差異 Fig.2 Interspecific difference of the ratio of shell-mass to body-mass
2.3 靜水沉降特性的種間差異
觀察發(fā)現(xiàn),除中華園田螺、銅銹環(huán)棱螺和河蜆外,其余貝類均具備在水面漂浮的能力。在靜水沉降實(shí)驗(yàn)中,殼形是影響沉降姿勢(shì)的主要因素,雙殼貝類和螺類的沉降行為差異明顯,其中雙殼類的沉降穩(wěn)定性較差,沉降終點(diǎn)距離理想降落點(diǎn)的距離較大,而螺類多為螺旋形下沉或平穩(wěn)形下沉,殼形接近的物種沉降行為相似。不同貝類沉降速度的種間差異如圖3所示。由圖可知,貝類沉降速度的變異范圍較大,其中橢圓蘿卜螺的沉降速度最低((4.13±0.96) cm/s),只與平角卷螺((9.86±3.04) cm/s)差異不顯著,但極顯著低于其他貝類(P<0.01);而河蜆的沉降速度最大((24.99±4.22) cm/s),極顯著高于其他貝類(P<0.01)。同為水生貝類,河蜆的沉降速度是橢圓蘿卜螺沉降速度的6倍;平角卷螺、同型巴蝸牛((12.73±3.93) cm/s)和背角無(wú)齒蚌((13.04±3.73) cm/s)3種貝類的種間沉降速度差異不顯著,但與其他貝類差異極顯著(P<0.01);非洲大蝸牛((18.30±3.64) cm/s)、銅銹環(huán)棱螺((19.48±3.14) cm/s)、中華園田螺((21.44±3.92) cm/s)和福壽螺((21.77±5.23) cm/s)4種貝類的種間沉降速度差異不顯著,但是與其他貝類差異極顯著(P<0.01)。
沉降加速度的種間差異如圖4所示,由圖可知橢圓蘿卜螺的沉降加速度最小(0.25±0.11 cm/s2),平角卷螺、同型巴蝸牛和背角無(wú)齒蚌的沉降加速度較為接近,處于較低水平。非洲大蝸牛、銅銹環(huán)棱螺、中華園田螺和福壽螺4種貝類的沉降加速度較為接近,處于較高水平,河蜆的沉降加速度最大(9.17±3.20 cm/s2),極顯著的高于其他貝類(P<0.01)。貝類的沉降速度和沉降加速度的種間差異不盡相同,但是種間差異的趨勢(shì)基本一致。
2.4 殼-體質(zhì)量比與水力學(xué)沉降特性的關(guān)系
9種受試貝類的沉降速度和沉降加速度與殼-體質(zhì)量比的相關(guān)關(guān)系如圖5所示,由圖可知,總體上,沉降速度、沉降加速度與殼-體質(zhì)量比的均值之間存在一定的相關(guān)性,相關(guān)性系數(shù)R2分別為0.28和0.39,但是沒(méi)有明顯的線性關(guān)系。而指數(shù)函數(shù)(0.15, 0.15)、冪函數(shù)(0.09, 0.09)和對(duì)數(shù)函數(shù)(0.19, 0.28)回歸分析得到的相關(guān)性系數(shù)更低,擬合度更小。
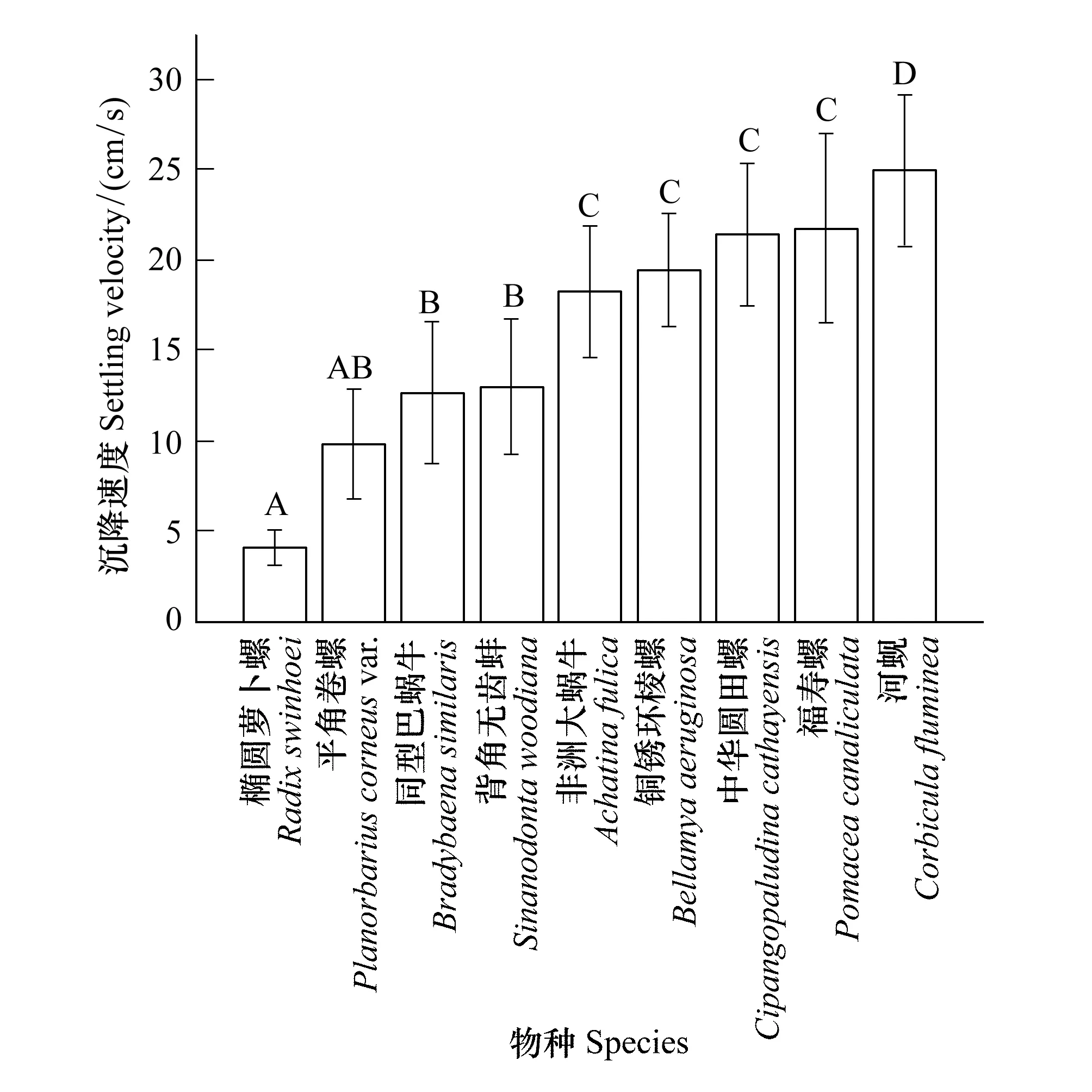
圖3 靜水沉降速度的種間差異Fig.3 Interspecific difference of the settling velocity in still water
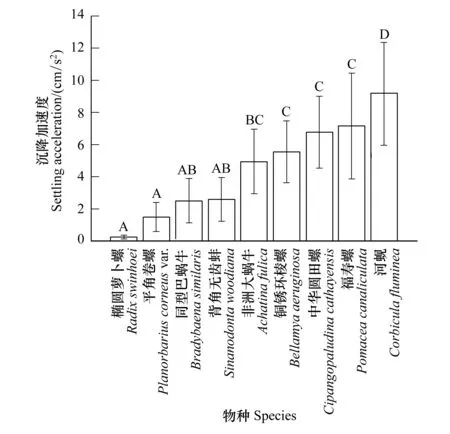
圖4 靜水沉降加速度的種間差異Fig.4 Interspecific difference of the settling acceleration in still water
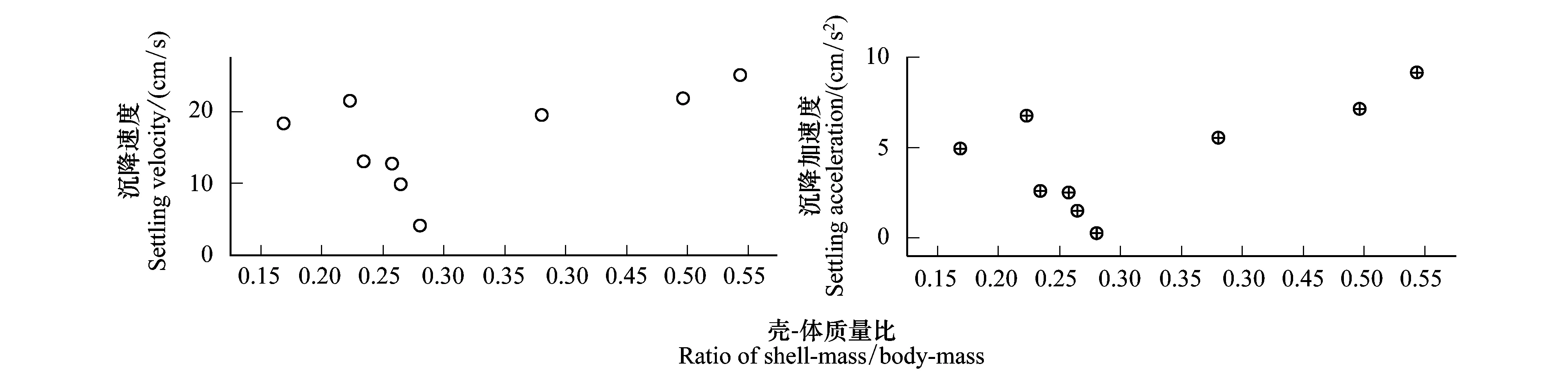
圖5 殼-體質(zhì)量比與沉降速度、沉降加速度的相關(guān)關(guān)系Fig.5 The relationship between shell-mass: body-mass and the settling characteristics (velocity and acceleration)
3 討論
3.1 殼-體質(zhì)量比的種間差異及其進(jìn)化形成原因
軟體動(dòng)物種類繁多且分布廣泛,現(xiàn)存種和化石種共有近15萬(wàn)種,貝殼是最重要的保護(hù)器官,其形成主要依靠外套膜表皮細(xì)胞的分泌物。瓣鰓綱(Lamellibranchia)貝類約2萬(wàn)種,全部營(yíng)水生生活,大部分海產(chǎn),少部分淡水種。腹足綱(Gastropoda)貝類約10萬(wàn)種,一般通稱螺類,包括前鰓亞綱(Prosobranchia)、后鰓亞綱(Opisthobranchia)及肺螺亞綱(Pulmonata)。廣布于海洋、淡水和陸地。貝類的干重、濕重與殼長(zhǎng)、殼寬、殼高等主要形態(tài)學(xué)參數(shù)存在明顯的線性關(guān)系,這些關(guān)系在河蜆[15]、環(huán)棱螺[16]、蘿卜螺[16]、福壽螺[14]等大多數(shù)物種中都得到了驗(yàn)證。帶殼濕重(體質(zhì)量)是估算淡水螺類生物量和能量值的最有效指標(biāo)[16]。因此,殼-體質(zhì)量比是形態(tài)對(duì)環(huán)境變化適應(yīng)性的一種綜合反應(yīng)。
殼形的可塑性和異速生長(zhǎng)在貝類中已被廣泛研究,環(huán)境因素可以誘導(dǎo)殼形的微結(jié)構(gòu)產(chǎn)生變異,受環(huán)境因子時(shí)空變異的選擇作用,螺類的生態(tài)表型可在殼的厚薄和大小之間產(chǎn)生進(jìn)化權(quán)衡[17]。因此盡管本研究中由于不同貝類生境特征的差異性和多樣性[18],水文環(huán)境特征可能并非是貝類殼-體質(zhì)量比種間差異形成的主要原因,但是陸生貝類與水生貝類之間、雙殼類與螺類之間受水力學(xué)作用的差異性是顯而易見(jiàn)的,只是這種具體選擇作用的實(shí)現(xiàn)過(guò)程還有待進(jìn)一步研究[19]。
有研究表明,淡水貝類在殼厚、殼型、生長(zhǎng)以及繁殖等特征上的表型可塑性是海水貝類的三倍以上,因此淡水貝類的表型可塑性在種間以及種內(nèi)的差異可能是極其顯著的[20],這與本研究發(fā)現(xiàn)淡水貝類的殼-體質(zhì)量比呈現(xiàn)出極顯著的種間差異是相一致的(P<0.01)。有研究表明生活在海岸帶的貝類,其形態(tài)和行為都會(huì)受到水流的影響并產(chǎn)生適應(yīng)性變化[10]。一般而言,貝殼的保護(hù)作用和貝類的運(yùn)動(dòng)速度的進(jìn)化方向是不相容的[10]。但是,某些貝殼較厚的螺類更能夠在一定的時(shí)間內(nèi)抵御水禽的消化作用,從而實(shí)現(xiàn)遠(yuǎn)距離傳播,這可能是貝殼同時(shí)具有保護(hù)作用和促進(jìn)擴(kuò)散能力的另一種體現(xiàn)[21]。
環(huán)境對(duì)貝殼形態(tài)不具備決定作用,而只有調(diào)控作用[22]。古代湖泊中的軟體動(dòng)物貝殼形態(tài)與其捕食者的進(jìn)化有關(guān),對(duì)于捕食者驅(qū)動(dòng)的貝類貝殼形態(tài)進(jìn)化,淡水貝類不如海洋貝類那么明顯[10]。具備最堅(jiān)固貝殼的淡水貝類只在古湖泊和古河流中有發(fā)現(xiàn),然而這種普遍的反捕食特征在現(xiàn)代貝類中已經(jīng)不復(fù)存在了[23]。大量研究表明,淡水螺類的殼形態(tài)指數(shù)與體質(zhì)量顯著相關(guān),但是相關(guān)關(guān)系會(huì)隨著生境條件的變化而不同[24]。除了本文所研究的成體貝類的殼-體質(zhì)量比特征以外,貝殼的強(qiáng)度、硬度和彈性模量等力學(xué)性能指標(biāo)也會(huì)隨著貝殼的生長(zhǎng)而增大[25]。因此,殼-體質(zhì)量比的種內(nèi)時(shí)空變異還需要進(jìn)一步的研究。
3.2 貝類水力學(xué)沉降特性的種間差異
貝類水力學(xué)沉降特征研究屬于生態(tài)水力學(xué)的范疇。生態(tài)水力學(xué)作為一門新興交叉學(xué)科,關(guān)注水力學(xué)與生態(tài)過(guò)程之間的關(guān)聯(lián),研究水力條件的改變對(duì)水生態(tài)系統(tǒng)平衡以及對(duì)生物多樣性的影響是其最重要的研究?jī)?nèi)容之一[26-27]。貝類水力學(xué)沉降特性種間差異的研究是對(duì)水文與生物體之間互作研究的一個(gè)有益補(bǔ)充[28-29]。極端氣候和人類活動(dòng)影響下,貝類水力學(xué)沉降特征的明確有助于對(duì)入侵、進(jìn)化等相關(guān)生態(tài)過(guò)程的理解[30]。
有研究表明河蜆形態(tài)在流域間存在顯著的區(qū)域變化,區(qū)域間殼體形態(tài)差異是水流速度的差異所致, 而水體化學(xué)組成和溫度變化與此種形態(tài)變化之間相關(guān)性不大[31]。沉降速度與沉降加速度是體質(zhì)量與形態(tài)的綜合作用結(jié)果。許多底棲無(wú)脊椎動(dòng)物都沒(méi)有對(duì)抗強(qiáng)烈水流的游泳能力[32],因此只能作為近似顆粒在水流中被動(dòng)運(yùn)動(dòng),隨水漂流和吸附能力對(duì)于擴(kuò)散、傳播和定殖尤其重要[33]。在底棲動(dòng)物生活史中利于種群擴(kuò)散的時(shí)期可能不盡相同[34],種群的擴(kuò)散傳播往往是通過(guò)整個(gè)生活史策略實(shí)現(xiàn)的。因此本研究中沉降速度較大的物種并不一定就是隨水流擴(kuò)散能力弱的物種,不過(guò)其他生活史階段的擴(kuò)散能力還有待進(jìn)一步研究。
3.3 貝類殼-體質(zhì)量比與水力學(xué)沉降特性的相關(guān)關(guān)系
本研究所涉及的貝類為我國(guó)南方地區(qū)常見(jiàn)的貝類,相關(guān)性分析表明殼-體質(zhì)量比和沉降特性之間的相關(guān)性總體上不顯著,并非殼-體質(zhì)量比越大沉降速度或沉降加速度就越大。殼-體質(zhì)量比是形態(tài)參數(shù)之間的一種關(guān)系,沉降特性是形態(tài)、行為功能與水環(huán)境之間互作的一種反應(yīng)。有研究表明螺類運(yùn)動(dòng)特征的進(jìn)化通常是不穩(wěn)定的,但是體型大小在進(jìn)化上則是相對(duì)較為保守的,而體型大小是運(yùn)動(dòng)能力最主要的限制因子之一[10]。福壽螺和非洲大蝸牛的體型與同類型螺類相比優(yōu)勢(shì)極其顯著,都具有較強(qiáng)的運(yùn)動(dòng)、擴(kuò)散能力,此二種螺類極強(qiáng)的攝食、破壞能力也與體型大小相關(guān)[35],并且成體階段都具有水中漂浮的能力,這可能部分解釋了其成為惡性入侵生物的原因。這與水文格局變化、長(zhǎng)距離運(yùn)輸和極端氣候改變了腹足類在人類世的地理分布的研究結(jié)果也是一致的[36]。
盡管體型對(duì)于貝類在生態(tài)上具有無(wú)可爭(zhēng)議的重要性,但這并不意味著在宏觀進(jìn)化上就一定會(huì)有直接的體現(xiàn)[37]。形態(tài)上的特化將降低貝類的運(yùn)動(dòng)、擴(kuò)散能力,但是擴(kuò)散能力是一個(gè)綜合性狀,并不是由某一特征單獨(dú)決定的。殼-體質(zhì)量比和沉降特性之間的相關(guān)關(guān)系不顯著,可能預(yù)示著貝殼特征在很大程度上是綜合生境適應(yīng)的結(jié)果。沉降速度和沉降加速度是成體貝類隨水流擴(kuò)散能力的一種反應(yīng),但生態(tài)水力學(xué)對(duì)于種群擴(kuò)散的綜合作用可能還需考慮整個(gè)生活史階段的作用以及貝類的運(yùn)動(dòng)、繁殖行為特征[38]。例如,福壽螺與中華園田螺和銅銹環(huán)棱螺的沉降速度差異并不顯著,但是前者在非應(yīng)激狀態(tài)下還具有在水中漂浮的能力,使其具備更強(qiáng)的擴(kuò)散能力。作為廣布種,這些貝類都具有較強(qiáng)的適應(yīng)能力[39],除了福壽螺和非洲大蝸牛等世界性入侵物種以外,河蜆、背角無(wú)齒蚌、同型巴蝸牛等在某些地區(qū)也被當(dāng)做入侵種,種群擴(kuò)散能力的研究是其入侵生態(tài)學(xué)研究的重要內(nèi)容[40]。不過(guò),貝殼表型與沉降行為特征各自的進(jìn)化路徑與生態(tài)功能以及二者之間的相關(guān)關(guān)系對(duì)于種群擴(kuò)散的作用力還有待深入研究[41]。
4 結(jié)論
貝類的殼-體質(zhì)量比存在極顯著的種間差異,種屬特異性強(qiáng),水生貝類的殼-體質(zhì)量比顯著大于陸生貝類,“殼-體質(zhì)量比”參數(shù)能較好的反應(yīng)貝類在生境適應(yīng)性上對(duì)運(yùn)動(dòng)與保護(hù)功能的權(quán)衡。貝類的靜水沉降速度和沉降加速度存在極顯著的種間差異,成體貝類的殼-體質(zhì)量比與沉降特性之間不存在普適性的顯著相關(guān)關(guān)系,其相關(guān)性的進(jìn)一步揭示還需考慮水文對(duì)不同生活史時(shí)期貝類種群擴(kuò)散的作用。同時(shí),貝類入侵種的危害性與其擴(kuò)散能力密切相關(guān),而擴(kuò)散能力是物種長(zhǎng)期適應(yīng)性進(jìn)化的重要結(jié)果,是包含物種個(gè)體形態(tài)特征、生理生態(tài)行為以及所處生境等多因素的綜合策略。
猜你喜歡
音樂(lè)探索(2022年2期)2022-05-30 21:01:37
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2021年12期)2021-12-31 03:23:08
中學(xué)生數(shù)理化·中考版(2020年10期)2020-11-27 01:59:48
小天使·一年級(jí)語(yǔ)數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國(guó)生殖健康(2019年2期)2019-08-23 08:12:08
中國(guó)特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
產(chǎn)品可靠性報(bào)告(2017年7期)2017-09-05 09:49:12
汽車觀察(2016年3期)2016-02-28 13:16:26
鄭州大學(xué)學(xué)報(bào)(醫(yī)學(xué)版)(2015年2期)2015-02-27 14:50:46
