放牧干擾背景下蘚結皮對毛烏素沙地土壤水分與風蝕的影響
2018-10-29 08:15:24李新凱卜崇峰李宜坪李炳垠鞠孟辰王清玄
水土保持研究 2018年6期
關鍵詞:深度
李新凱, 卜崇峰, 李宜坪, 李炳垠, 鞠孟辰, 王清玄
(1.中國科學院 水利部 水土保持研究所, 陜西 楊凌 712100; 2.中國科學院大學, 北京 100049;3.西北農林科技大學 水土保持研究所, 陜西 楊凌 712100; 4.西北農林科技大學 水利與建筑工程學院, 陜西 楊凌 712100)
生物土壤結皮是由土壤顆粒與藻類、真菌、地衣、苔蘚以不同的比例緊密結合而形成,是干旱、半干旱地區常見的地表覆蓋物[1-3]。苔蘚、真菌和許多地衣的原絲體、菌絲和假根,以及許多藻類產生的絲狀體能夠固定土壤顆粒,與高等植物根系固定土壤顆粒的功能很相似[4]。大量研究證實生物結皮能夠增強土壤穩定性,提高土壤抵抗風蝕和水蝕的能力[5-7]。沙漠地區的維管植物通常低矮稀疏,個體間出現較大的空地,枯落物量也比較少,在干旱、半干旱地區抵御土壤風蝕的作用有限,生物結皮對土壤穩定性的作用在這種情況下顯得尤為重要[8]。
在干旱、半干旱地區,水是限制植物生長主要的因素之一[9]。生物結皮的存在會導致沙地生態系統土壤水分重分布[10],生物結皮通過改變土壤理化性質和生物學特性影響局部水文循環的不同方面[3,11]。而學者在騰格里沙漠和毛烏素沙地的研究表明,發育穩定且具有高蓋度的生物結皮影響降水向深層土壤入滲,導致土壤水分淺層化,影響當地灌木生長[12-13]。干擾影響生物結皮蓋度、物種組成和生理功能[14-15],生物結皮受到干擾后,土壤屬性、微地形等環境條件發生改變,土壤水文過程也隨之變化。因此,有學者提出了破壞生物結皮以增加干旱區植被生態用水、防治水土流失的構想,我國學者也在1991年提出對生物結皮進行適度干擾的生態管理理念[16]。少量研究初步證實,適度的移除、踩踏等人為干擾能夠在不顯著增加侵蝕的情況下增加入滲,改善土壤水分狀況[17-19]。調查發現,苔蘚主導的生物結皮在毛烏素沙地廣泛分布,放牧干擾是毛烏素沙地生物結皮主導的干擾類型。截止目前,有關生物結皮土壤水文、風蝕等對放牧干擾響應的研究鮮有報道,生物結皮適度干擾的可行性尚有爭議[20]。基于以上原因,本研究在前期調查的基礎上,在毛烏素沙地選定一處典型的蘚結皮樣地進行土壤水分和風蝕的長期定位觀測,探討在放牧干擾的背景下,生物結皮對毛烏素沙地土壤水分和風蝕的影響,并驗證適度干擾的可行性,以期為毛烏素沙地生物結皮資源的保護和管理提供借鑒。
1 試驗材料與方法
1.1 研究地概況
研究樣地位于陜西省榆林市榆陽區孟家灣鄉,地理坐標109°36′14.07″—109°36′24.68″E,38°32′50.57″—38°32′54.60″N,海拔1 148~1 153 m。該地區屬中溫帶半干旱大陸性季風氣候,年平均氣溫8.3℃,年均降水量405 mm,降水主要集中在6—9月[21]。該樣地處于毛烏素沙地東南緣,地貌類型以起伏的固定和半固定沙丘為主。主導的灌木為沙蒿(Artemisiadesertorum)、沙柳(Salixpsammophila)和檸條錦雞兒(Caraganakorshinskii),在樣地內多呈簇狀均勻散布。草本植物主要為沙竹(Psammochloavillosa)和狗娃花(Heteropappushispidus),在樣地內零星分布。蘚結皮在地勢低洼的丘間地多成斑塊狀或連片分布,沙蒿、沙柳和檸條的冠層下方及植被間空地都存在蘚結皮。通過對牲畜遺留的蹄印、糞便的分布情況的調查發現,放牧干擾多發生在植被間空地,分布于植被間的蘚結皮更易受到干擾,而植被投影下方的蘚結皮幾乎無干擾發生。樣地內的蘚結皮大多已發育至穩定階段,結皮形態多呈丘狀起伏,具有較強的抗侵蝕能力[3]。因受干擾的程度不同,結皮蓋度在不同位置差異較大,蓋度最高可達90%以上。
1.2 研究方法
1.2.1 小區布設 試驗小區設置及立地背景見表1,試驗小區實景見圖1。植被投影下方小區由植被下蘚結皮和植被下裸地小區組成,按植被類型分為3個對照組(以植被下裸地為對照),每個對照組選取大小和形態特征相似灌叢,在其投影下方建立小區。沙蒿、檸條和沙柳下蘚結皮小區建立在蓋度較高且發育良好的蘚結皮斑塊上,植被下裸地小區建立在植被下方無結皮的裸沙上。植被間空地小區由裸地和不同干擾程度的蘚結皮小區組成。裸地小區建立在周圍無植被的裸沙地塊上。放牧干擾在蘚結皮斑塊施加的壓力和剪力作用使完整的蘚結皮趨于破碎化,蘚結皮的蓋度與干擾強度呈正相關[14]。本研究中的放牧干擾為自然狀態下的隨機干擾,參考馮偉[18]、葉菁[19]等有關干擾強度的劃定方法,我們將樣地內植被間蘚結皮的放牧干擾劃分為5個干擾強度:無干擾(蓋度>80%)、輕度干擾(60%<蓋度<80%)、中度干擾(40%<蓋度<60%)、重度干擾(20%<蓋度<40%)和劇烈干擾(蓋度<20%)。在樣地內的植被間空地選定符合以上5個干擾強度的蘚結皮斑塊建立不同干擾強度下的蘚結皮小區。試驗小區呈邊長為1 m的正方形,每個處理2個重復,重復小區留1 m左右間隔,在小區的4個頂點釘上木樁以確定邊界。為防止小區在觀測期內再次受到放牧的影響,小區劃定完成后,在周邊設置鐵絲網圍欄加以保護。最后在每個小區的中心位置用專用土鉆打孔,埋設2 m長的TDR水分測管,用于剖面土壤水分的測定;并在每個小區中部垂直插入1根50 cm長,直徑2 mm的鋼釬,所有鋼釬起始時地上部分長30 cm,埋深20 cm,用于監測各小區風蝕量。

圖1 試驗小區實景
1.2.2 測定項目與方法 觀測試驗開始前1周測定蘚結皮蓋度、厚度和土壤容重指標(表1)。生物結皮蓋度(%)用目估法測定,由2位不同的試驗員分別進行估計,每個處理得到2個蓋度估計值。生物結皮厚度(mm)用游標卡尺測定,在每個生物結皮小區隨機選2小塊剖面完整的蘚結皮,用游標卡尺測定剖面厚度,每個小區獲得2個厚度值。土壤容重用環刀在樣地內隨機選3個裸地和3個蘚結皮地塊取土樣,用烘干法測定土壤容重。土壤體積含水量(%)用TDR(TRIME-PICO-IPH/T3)測定,1 m以內測定間隔10 cm,1~2 m測定間隔20 cm,從2017年7月24日起每周測定1次。風蝕量(t/hm2)觀測從2016年9月3日起,每隔2個月測定1次。用鋼尺定期測定鋼釬露出土壤表面的高度,得到前后兩次高度差Δh,結合測得的不同斑塊類型所對應的土壤容重ρb,由下述公式[16]計算得到風蝕量。土壤含水率和風蝕量觀測截止日期為2017年9月23日。
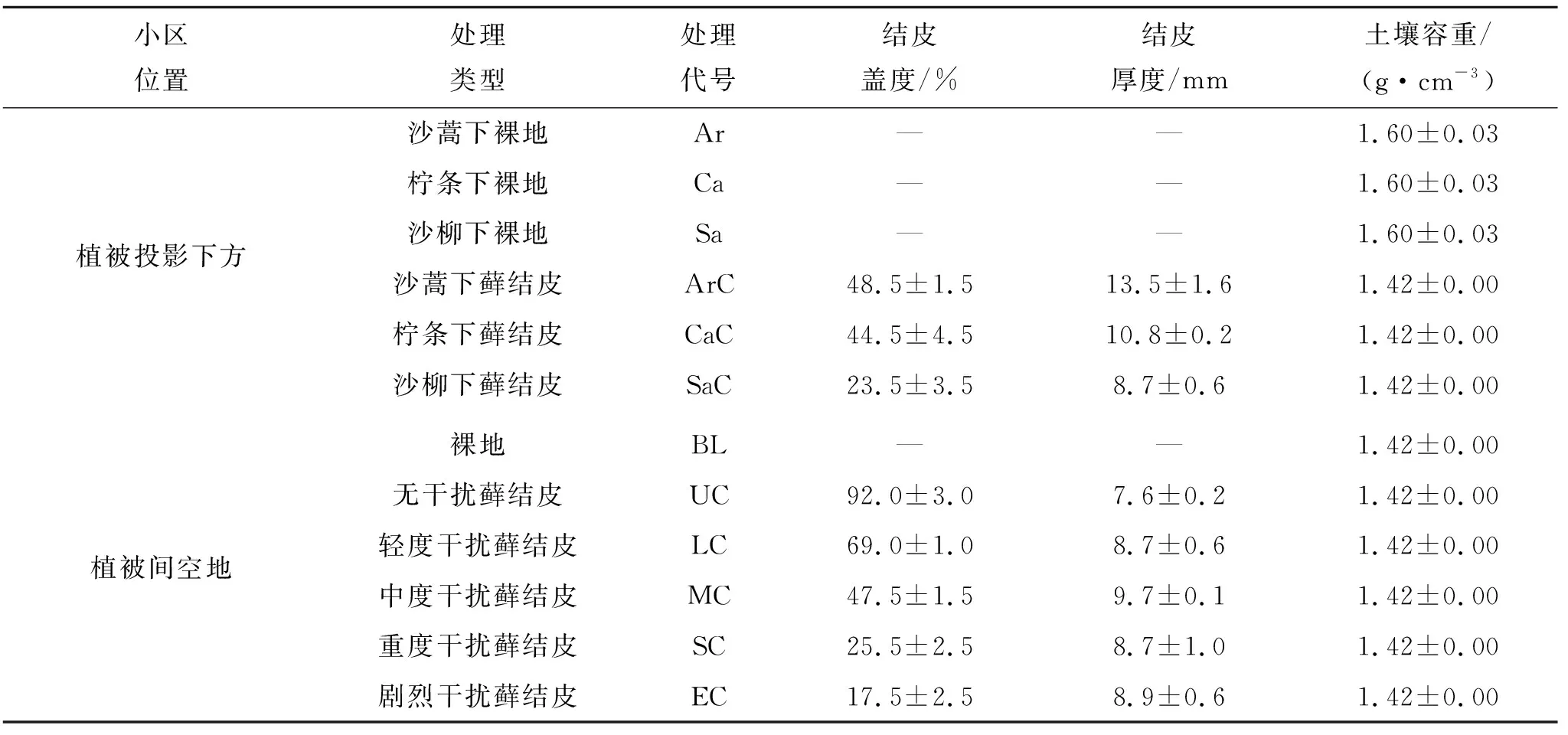
表1 試驗小區設置及立地背景
W=100ρbΔh
式中:W為風蝕量(t/hm2);Δh為風蝕/風積厚度(前后兩次測釬讀數差值,風蝕為正,風積為負值)(cm);ρb為土壤容重(g/cm3)。
1.3 數據處理
試驗數據用Microsoft Excel 2007處理,用GraphPad Prism 7進行相關性分析和繪圖。
2 結果與分析
2.1 蘚結皮對土壤水分的影響
2.1.1 植被下土壤含水量 裸地小區土壤含水量在0—30 cm深度內逐漸增大,30 cm深度處達到峰值,在30—120 cm深度內波動下降,在120—200 cm深度內波動上升。沙地淺層土壤對降水響應迅速,受蒸散發影響強烈,地下水位呈波動下降趨勢;深層土壤對降水的響應相對滯后,受蒸散發影響微弱,接受降雨入滲補給后,地下水位呈緩慢上升趨勢[22-23]。學者們在毛烏素沙地的研究表明,植被影響土壤含水量在土壤剖面的分布,植被的存在導致根系層土壤含水量的下降[23-25]。沙蒿、沙柳和檸條根系垂直方向主要分布在0—90 cm土層[24,26-30],從圖2A可以看出,3種植被下裸地小區的土壤含水量在0—70 cm深度范圍內明顯低于裸地,證實了植被對土壤含水量的影響。在0—80 cm深度范圍內,沙蒿、檸條和沙柳下蘚結皮小區的土壤含水量與對應的植被下裸地小區相比,分別提高了62.8%,56.2%和67.5%,說明蘚結皮的存在能夠提高植被根系層土壤水分。在80—200 cm深度范圍內,植被下裸地和植被下蘚結皮小區土壤含水量均低于對應的植被下裸地小區(表2),植被下蘚結皮小區在120—200 cm深度內土壤含水量的增幅也明顯低于對應的植被下裸地小區。蘚結皮的存在使植被根系層下方80—200 cm深度內的土壤含水量下降,這可能與蘚結皮限制降水入滲有關[12-13]。灌叢下方的蘚結皮在干燥情況下會吸收水分,結皮中微生物分泌的化學物質吸水膨脹后會阻塞土壤孔隙,影響水分入滲[4]。因此,在毛烏素沙地,蘚結皮的存在有助于提高沙蒿、沙柳和檸條0—80 cm的根系層土壤水分,同時會造成80—200 cm的深層土壤含水量下降。
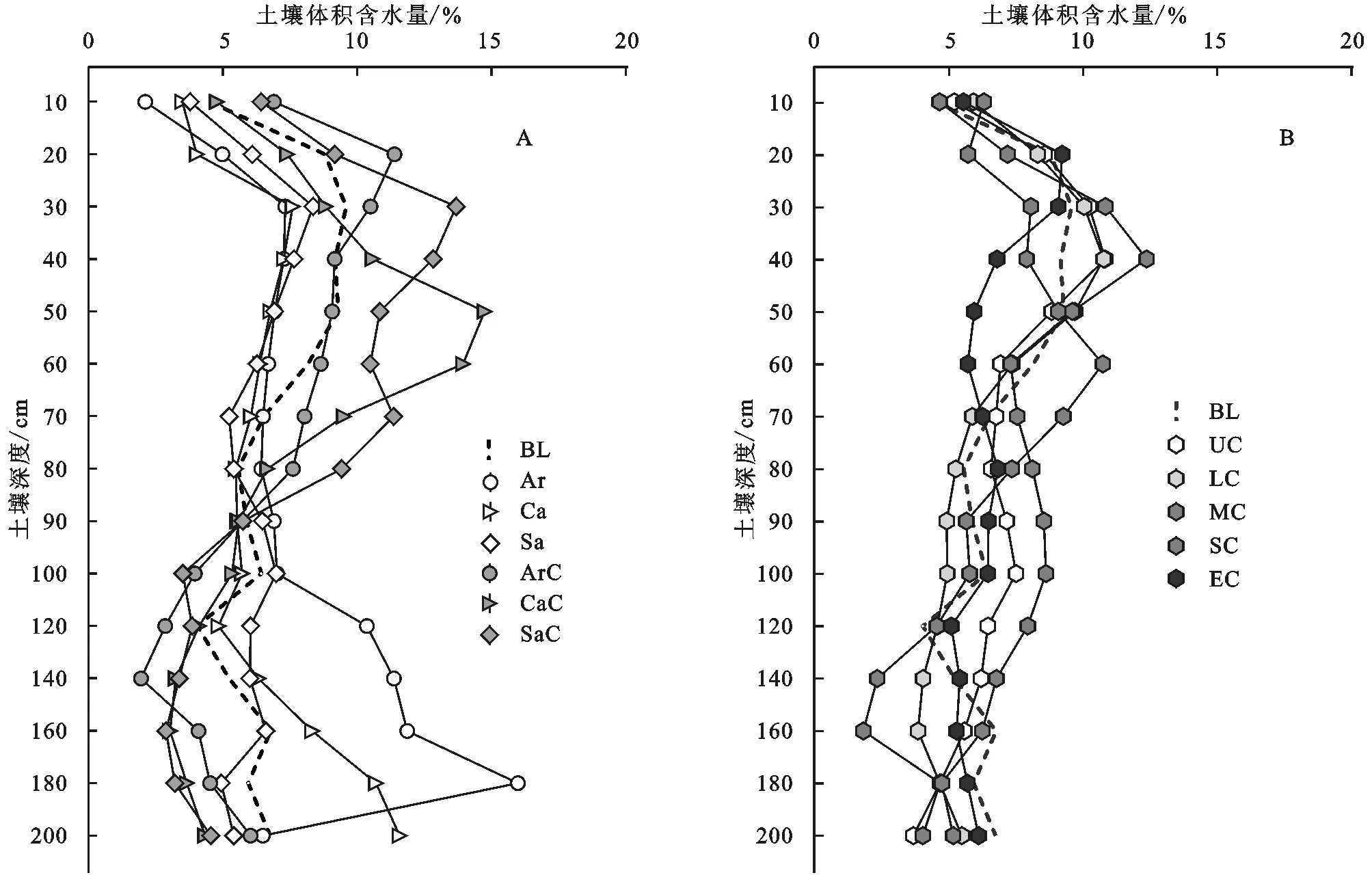
注:BL表示裸地;Ar表示沙蒿下裸地;Ca表示檸條下裸地;Sa表示沙柳下裸地;ArC表示沙蒿下蘚結皮;CaC表示檸條下蘚結皮;SaC表示沙柳下蘚結皮;UC表示無干擾蘚結皮;LC表示輕度干擾蘚結皮;MC表示中度干擾蘚結皮;SC表示重度干擾蘚結皮;EC表示劇烈干擾蘚結皮。
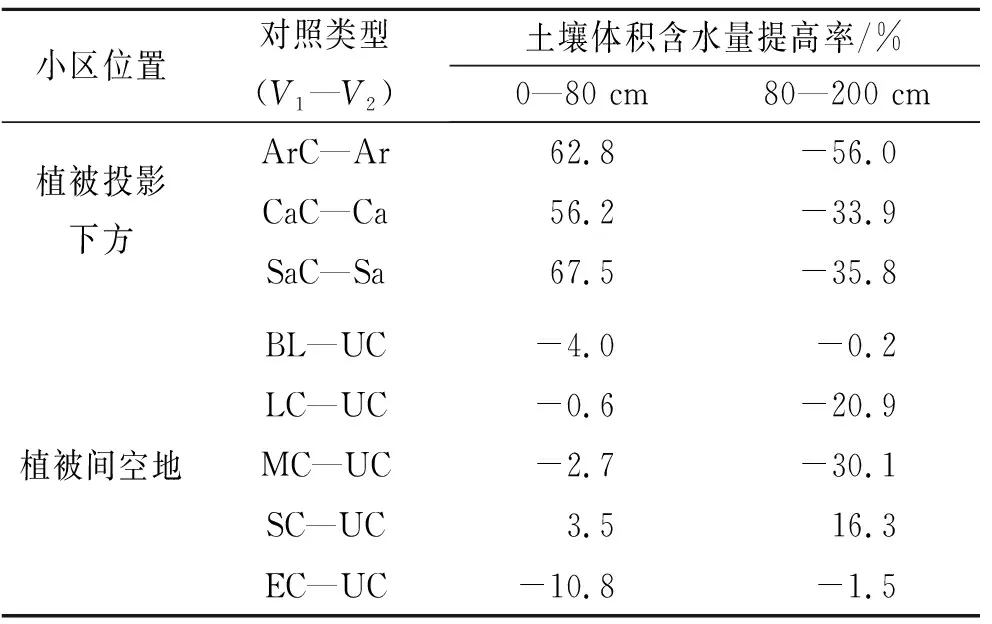
圖2 土壤水分隨深度的變化表2 土壤體積含水量提高率
注:土壤體積含水量提高率=(V1-V2)×100%/V2,V1和V2為該深度范圍內土壤體積含水量的均值。
2.1.2 植被間土壤含水量 無干擾蘚結皮在0—40 cm和70—140 cm深度范圍內的土壤含水量高于裸地(圖2B),與裸地對照相比,無干擾蘚結皮小區0—80 cm和80—200 cm深度內的土壤含水量分別提高4%和0.2%(表2)。說明植被間無干擾的蘚結皮同樣能提高土壤含水量。無干擾蘚結皮小區在80—200 cm深度內的土壤含水量高于3種植被下蘚結皮小區,但與裸地相比并未明顯增加。0—200 cm深度內,無干擾和干擾小區土壤含水量隨深度增加呈現出先升高后波動下降的趨勢。輕度、中度和重度干擾小區土壤含水量僅在幾個深度處高于無干擾小區;重度干擾小區僅在0—20 cm深度內土壤含水量低于無干擾,20—200 cm深度內土壤含水量均高于無干擾。從表2也可看出,與無干擾相比,輕度和中度干擾土壤水分含量在0—80 cm深度內略有下降,在80—200 cm深度范圍內明顯下降;重度干擾蘚結皮在0—80 cm和80—200 cm深度范圍內的平均土壤含水量分別提高3.5%和16.3%;劇烈干擾蘚結皮小區的土壤含水量在0—80 cm深度內下降10.8%,在80—200 cm深度內略有下降。不同干擾強度對應的結皮蓋度與0—80 cm,80—200 cm深度內土壤含水量均值的Person相關系數r分別為0.31和-0.29,統計上不顯著,表明干擾強度與0—80 cm,80—200 cm深度內的土壤含水量非線性相關,土壤水分隨干擾強度變化似乎無規律可循。但從土壤水分含量上看,重度干擾蘚結皮小區0—80 cm和80—200 cm深度內的土壤含水量與無干擾相比分別提高3.5%和16.3%,其余干擾強度的土壤水分含量僅在幾個深度處高于無干擾小區。生物結皮對土壤蒸發有一定的抑制作用[31],干擾使完整的生物結皮破碎,在促進入滲的同時也會影響蒸發。輕度和中度干擾對結皮蓋度的影響并不能明顯提高土壤含水量,劇烈干擾對蘚結皮造成過度破壞,可能加劇土壤蒸發,導致0—80 cm深度內的土壤含水量下降。相比之下重度干擾有利于提高0—80 cm的根系層和80—200 cm的深層土壤水分含量。
2.2 蘚結皮對風蝕的影響
2.2.1 植被下土壤風蝕量 裸地小區9—11月發生輕微風蝕,11—3月風積量逐漸增大,3—7月風積逐漸減少,5—7月風蝕量達到最大,7—9月出現少量風積(圖3A)。植被下裸地和植被下蘚結皮小區風蝕主要發生在5—9月,5月之前以風積為主。沙蒿下裸地風蝕頻率最高,其余小區風蝕和風積頻率相當(表3)。植被下裸地小區觀測期內的風蝕總量排序為沙蒿下裸地>沙柳下裸地>檸條下裸地。蘚結皮的存在能夠提高植被下方沙土的抗風蝕能力。與植被下裸地相比,沙蒿、檸條和沙柳下蘚結皮小區的風蝕量分別減少了92.8%,11.3%和104.2%。絕大多數研究都證實生物結皮具有減輕風蝕的作用[8]。許多蘚類植物的莖葉體和假根共同參與錨固土壤的過程,呈分枝狀的苔蘚原絲體在整個結皮層中散布,這些植物體結構與土壤顆粒緊密結合,提高了表層土壤的穩定性[2]。結皮中的藍藻、真菌分泌的多糖和有機膠體能粘結土壤顆粒,產生更大的土壤團聚體,從而提高啟動風速,減輕土壤風蝕[8]。我們的研究結果表明,研究樣地的風蝕主要發生在5—9月,蘚結皮等存在能夠提高植被下方沙土的抗風蝕能力,與植被下裸地相比,沙蒿、檸條和沙柳下蘚結皮小區的風蝕量分別減少了92.8%,11.3%和104.2%。不同植被類型下的小區和不同植被與蘚結皮組合小區的風蝕量也都存在差異。董治寶等[32]研究表明,植物是通過改變氣流對地表物質的作用效果來影響風蝕,大約在植被高度以下,風速變化明顯受植被個體影響。沙蒿、檸條和沙柳這3種植被在冠層結構、植物體形態、莖葉疏密程度等方面的差異可能是引起地表風蝕差異的主要原因。蘚結皮的蓋度差異以及植被與蘚結皮組合類型也可能影響風蝕量。
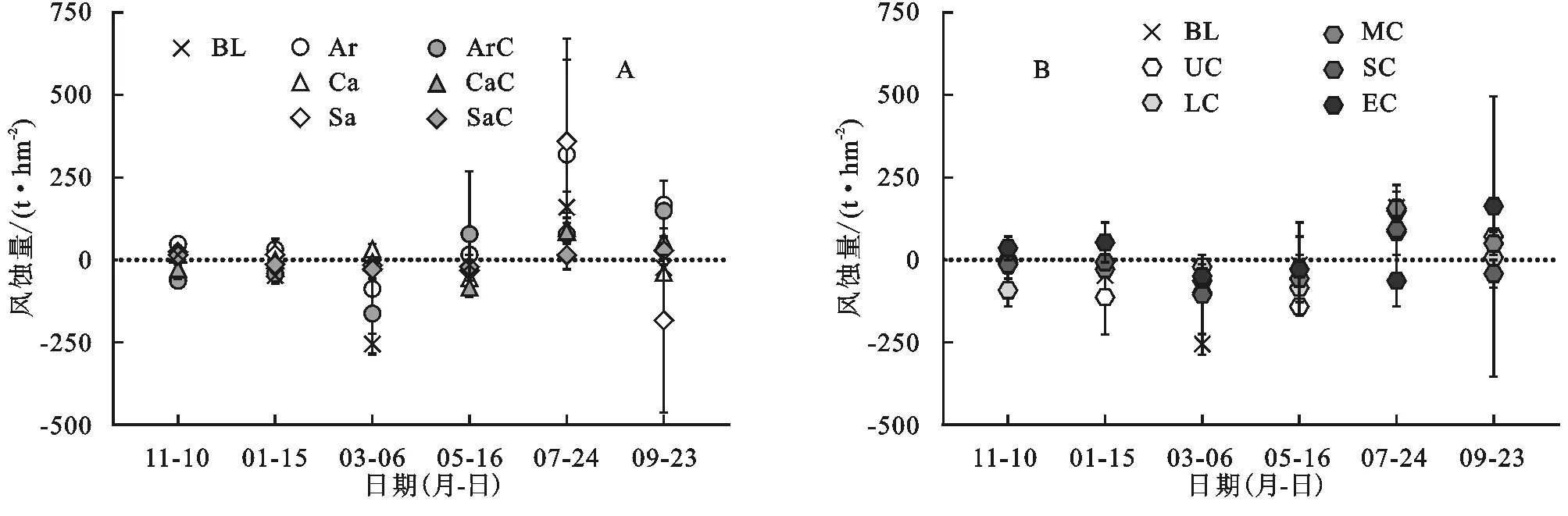
圖3 各小區風蝕量
2.2.2 植被間土壤風蝕量 觀測期內,植被間小區5月之前以風積為主,劇烈干擾小區風蝕量極大值(162.9 t/hm2)出現在7—9月,其余小區的風蝕量極大值集中出現在5—7月,排序為裸地(159.7 t/hm2)>中度干擾(155.9 t/hm2)>輕度干擾(148.8 t/hm2)>重度干擾(92.1 t/hm2)>無干擾(85.0 t/hm2)。在5—7月,裸地的風蝕量高出無干擾近1倍,中度和輕度干擾風蝕量明顯高于重度干擾和無干擾。裸地小區的風積頻率為風蝕頻率的2倍,風積總量大于風蝕總量,最終產生風積(表3)。無干擾蘚結皮小區僅在5—9月出現風蝕,整個觀測期內的風蝕總量低于風積總量,最終出現風積。說明在5—9月風蝕集中時期,無干擾的蘚結皮下層沙土并未發生風蝕,出現的風蝕量是蘚結皮表面風積沙土在風力作用下被重新搬運的結果。無干擾小區和裸地小區的風積總量基本相等。在風蝕較弱的部分時段(9—翌年1月,3—5月),隨著干擾強度增大,風積量有減少趨勢,風蝕量有增加趨勢。但在風蝕集中的5—9月,這一趨勢并不明顯。不同干擾強度對應的結皮蓋度與平均風蝕量之間的Person相關系數r為-0.77,統計不顯著,表明干擾強度與風蝕量同樣無線性相關關系。輕度和重度干擾蘚結皮小區發生風積,重度干擾的風積總量高于輕度干擾;中度和劇烈干擾蘚結皮小區最終產生風蝕,劇烈干擾風蝕總量(109.8 t/hm2)高于中度干擾(14.2 t/hm2)。從風蝕和風積頻率來看,無干擾、輕度和中度干擾風積頻率高于風蝕頻率;重度干擾風蝕頻率最低,風積頻率最高;劇烈干擾風蝕頻率最高,風積頻率最低。放牧、踩踏等人類活動是干旱生態系統常見的干擾類型,小尺度、低強度的干擾有助于生物結皮和其他植物的生長,而超過環境承載力的強烈干擾必然會威脅生態系統穩定[15],尤其是在生態環境脆弱地區,過度的人為干擾會破壞地表穩定性,影響生物結皮固碳、固氮等功能,加劇水土流失和養分損失[33]。在研究地,風蝕集中的5—7月,輕度和中度干擾小區的風蝕量接近裸地,重度干擾風蝕量相對較低,劇烈干擾具有較高的風蝕頻率和最高的風蝕總量,且出現了風蝕量極大值。因此,蘚結皮受到劇烈及以上強度的干擾會使土壤更易遭受風蝕。重度干擾發生風蝕的頻率最低,風蝕嚴重期風蝕量較小。
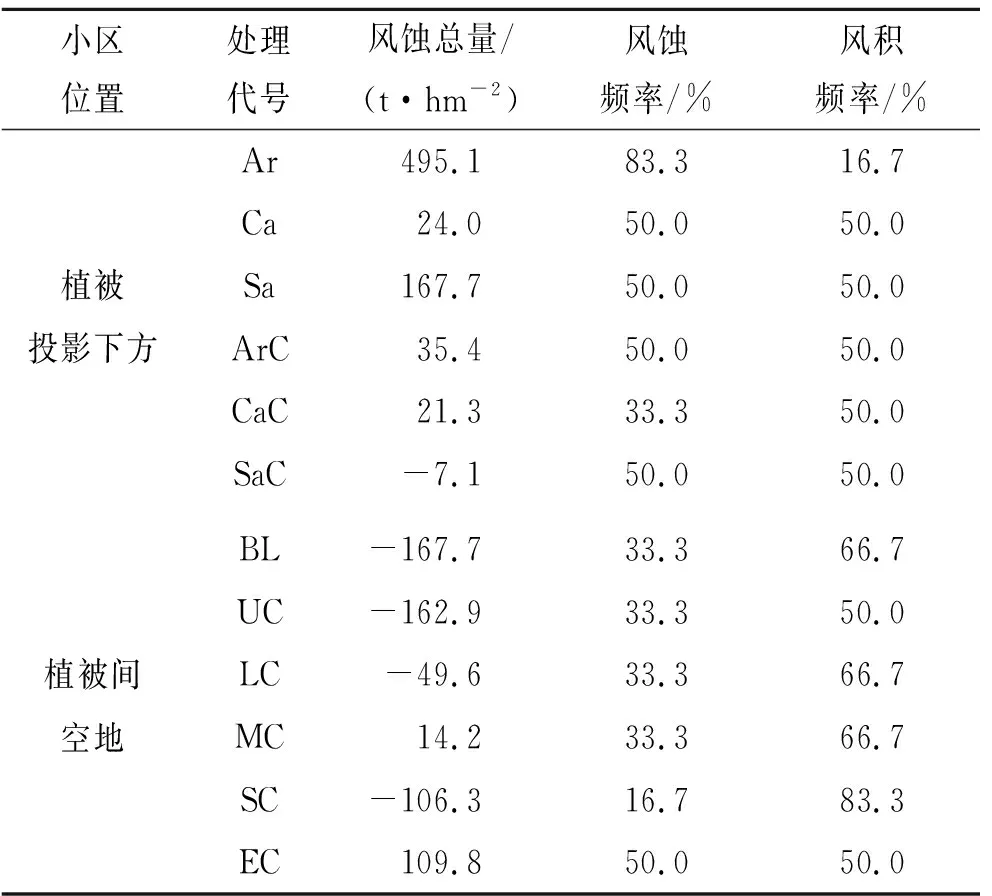
表3 風蝕總量與風蝕、風積頻率
注:風蝕總量正值表示風蝕,負值表示風積;風蝕或風積頻率為風蝕或風積出現次數與觀測總次數(6次)的百分比。
綜合上述土壤水分和風蝕的結果來看,重度干擾能夠在不加劇侵蝕的情況下提高土壤水分含量,這證實了適度干擾的可行性。關于不同干擾類型對生物結皮風蝕的影響已有大量報道[14-15],但目前尚缺乏有關干擾強度對生物結皮風蝕影響機理的研究。我們的試驗結果也局限于小尺度地塊,還需要在更多地區和更大尺度上加以驗證。
3 結 論
(1) 蘚結皮的存在能夠提高沙蒿、沙柳和檸條0—80 cm的根系層土壤水分,同時也使80—200 cm的深層土壤含水量下降。干擾強度與土壤水分含量非線性相關。植被間無干擾的蘚結皮同樣能提高土壤含水量。重度干擾有利于提高0—80 cm的根系層和80—200 cm的深層土壤水分含量。
(2) 觀測期內,風蝕主要發生在5—9月。蘚結皮的存在有利于減輕植被下方沙土風蝕,沙蒿、檸條和沙柳下蘚結皮小區的風蝕量與植被下裸地相比分別減少了92.8%,11.3%和104.2%。干擾強度也與風蝕量非線性相關。蘚結皮受到劇烈及以上強度的干擾會使土壤更易遭受風蝕,重度干擾風蝕發生頻率最低,風蝕嚴重期的風蝕量也較小。
(3) 從植被間小區的土壤水分和風蝕量綜合來看,重度干擾蘚結皮小區在不加劇風蝕的情況下,有利于提高0—200 cm深度內的土壤水分,肯定了適度干擾的可行性。
猜你喜歡
中學生數理化·七年級數學人教版(2022年6期)2022-06-05 06:50:50
快樂學習報·教育周刊(2022年16期)2022-05-01 21:25:05
中學生數理化·七年級數學人教版(2020年11期)2020-12-14 06:59:52
藝術品鑒證.中國藝術金融(2018年8期)2019-01-14 01:14:28
藝術品鑒證.中國藝術金融(2018年10期)2019-01-08 02:44:26
藝術品鑒證.中國藝術金融(2018年6期)2019-01-08 02:43:04
藝術品鑒證.中國藝術金融(2018年12期)2018-08-26 06:03:48
新聞傳播(2016年10期)2016-09-26 12:14:59
新聞傳播(2015年10期)2015-07-18 11:05:40
交通建設與管理(2015年15期)2015-03-20 15:18:57
