黃土丘陵溝壑區抵抗土壤侵蝕的植物群落結構特征
2018-10-29 08:15:24焦菊英
水土保持研究 2018年6期
寇 萌, 焦菊英
(1.咸陽師范學院 資源環境與歷史文化學院, 陜西 咸陽 712000; 2.中國科學院 水利部 水土保持研究所黃土高原土壤侵蝕與旱地農業國家重點實驗室, 陜西 楊凌 712100; 3.西北農林科技大學 水土保持研究所, 陜西 楊凌 712100)
黃土丘陵溝壑區位于半濕潤半干旱向干旱荒漠地區的過渡地帶。眾所周知,由于地理位置的過渡性、地形和地貌的復雜性、土壤的易蝕性及人類對土地的過度利用,該區生態環境不斷惡化,侵蝕強烈,已成為我國水土流失與生態環境問題最嚴重的地區之一[1]。植被是連接生態系統生物與非生物環境的重要紐帶,是恢復和維持生態系統的物質基礎,對減少水土流失、改善生態環境有重要作用[2]。朱顯謨[3]指出,植物是防治水土流失的最有效和最根本措施。植物群落在垂直方向上由不同的植被層次組成,不同的植被層次具有不同的水土保持作用。如植被的林冠層、灌層能夠截留降水,改變降雨動能,起到緩沖作用;近地面的草被對于雨滴的攔截作用可以有效減緩雨滴速度,減少雨滴動能和濺蝕量,保護地表土壤免受侵害[2];貼地面的枯枝落葉層和結皮層直接覆蓋地表,可避免雨滴直接作用于土層,減少土壤濺蝕[4-6],也能有效攔截地表徑流,減弱其剝蝕能量,減少細溝或切溝侵蝕的發生[7]。
因此,植被要產生良好的水土保持作用,就必須具有一定的植被覆蓋度,良好的結構,這是植被保持水土的關鍵[8]。恢復具有合理結構與有效蓋度的植被也是控制該區水土流失、改善生態環境的根本途徑[9-10]。但目前對植被水土保持作用的評估,主要以單純的植被投影蓋度表示,并未考慮植物群落的垂直結構層次,難以反映植物群落結構差異對水土流失的影響。Wen等[11]提出了結構化植被因子指數Cs的概念及計算模型,該指數是根據植物群落的垂直結構特征及不同植被垂直層在水土保持作用方面的差異,綜合考慮了植被類型、群落的垂直結構組成和各層次蓋度多個要素的植被蓋度指標[11-12]。由于土壤表層結皮同樣具有水土保持作用,能增加土壤入滲、土壤抗沖性,減少土壤濺蝕等[4,13-14],但該指數模型并未提及。本研究將在原指數模型的基礎上,增加結皮層的水保作用系數,進一步完善該指數模型,通過對不同植物群落各垂直結構層的蓋度統計,計算群落的結構化植被因子指數,并結合前人監測的不同群落類型土壤侵蝕量數據,探明能夠抵抗土壤侵蝕的群落結構特征,為該區植被恢復和生態建設提供基礎資料與依據。
1 研究區與研究方法
1.1 研究區概況
本研究以陜北黃土丘陵溝壑區的延河流域(36°23′—37°17′N,108°45′—110°28′E)為研究區(圖1)。該流域氣候屬暖溫帶大陸性半干旱季風氣候,年均溫8.8~10.2℃,年均降水約500 mm,其中7—9月的降雨量占全年降雨量的60%以上,徑流量占全年的99%以上[15]。流域從東南向西北,氣候、溫度有明顯的梯度變化特征;植被隨環境梯度的變化也很明顯,從南向北依次為森林帶、森林草原帶和草原帶[16-17]。流域南部為森林帶北緣,地貌類型以梁峁狀黃土丘陵為主,主要建群種為遼東櫟(Quercusliaotungensis)、側柏(Platycladusorientalis)、三角槭(Acerbuergerianum)等,灌木有黃刺枚(Rosaxanthina)、虎榛子(Ostryopsisdavidiana)、繡線菊(Spiraeapubescens)等,草本植物有披針葉薹草(Carexlanceolata)等,人工喬、灌木林有刺槐(Robiniapsendoacacia)、檸條(Caraganaintermedia)等[18]。流域中部為森林草原帶,是典型的梁峁狀溝壑區[19],因退耕還林草政策的實施,分布大量人工植被和自然恢復植被,喬木主要以刺槐林為主,灌叢主要是檸條、沙棘(Hippophaerhamnoides)等人工灌叢及封禁后形成的黃刺玫、狼牙刺(Sophoraviciifolia)等天然灌叢,草本植物主要是鐵桿蒿(Artemisiagmelinii)、長芒草(Stipabungeana)、白羊草(Bothriochloaischaemun)、達烏里胡枝子(Lespedezadavurica)等[20]。流域北部是草原帶,地處毛烏素沙地南緣,是黃土高原向沙地過渡的敏感區域,植被屬溫帶干旱草原型,主要物種有冷蒿(Artemisiafrigida)﹑百里香(Thymusmongolicus)等[21]。
1.2 研究方法
本研究擬采用Wen等[11]依據植物群落的垂直結構特征提出的結構化植被因子指數Cs,并結合土壤侵蝕量的侵蝕針實測數據,分析抵抗土壤侵蝕的群落結構特征。
其基本模型為[12]:
(1)
式中:ai為不同植被層的水土保持作用系數;Ci為各植被層的實際蓋度;i代表喬木冠層、灌木層、草被層、枯枝落葉層。考慮到土壤表層結皮對土壤侵蝕的影響,本研究在原模型基礎上,增加了土壤表層的結皮層。
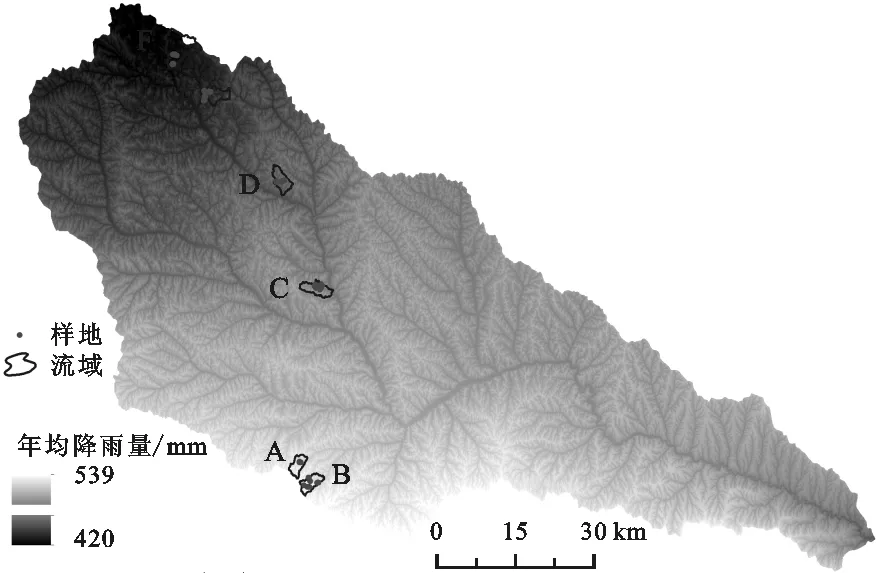
注:A為尚合年流域;B為毛堡則流域;C為陳家坬流域;D為張家河流域;E為石子灣流域;F為高家溝流域。
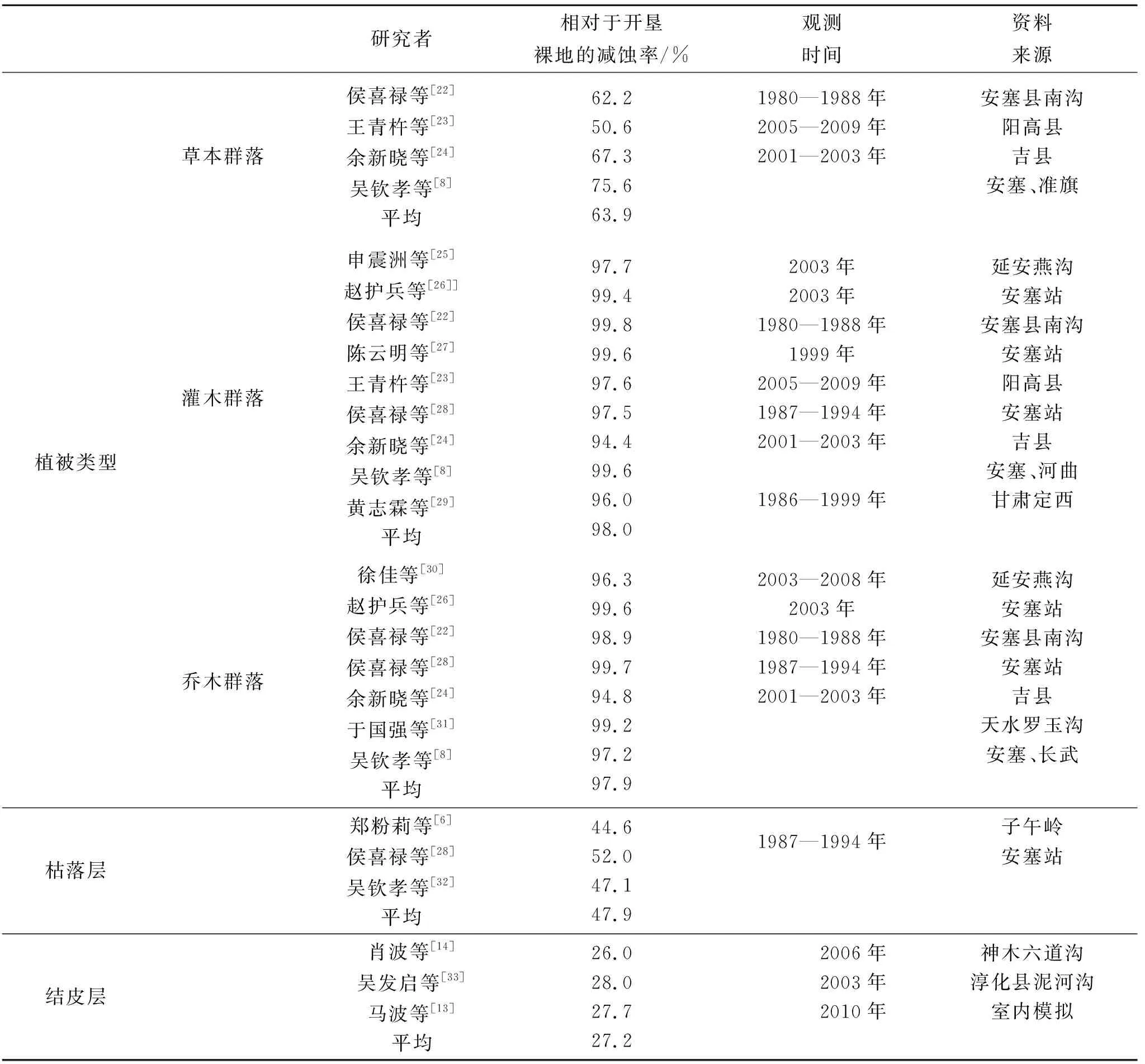
圖1 延河流域和典型小流域位置及樣地分布表1 主要植被類型減蝕率
由于各覆蓋層次在空間上分布及其控制水土流失機理的差異導致每個層次在群落控制水土流失效益中的作用有所差別。本研究通過搜集陜北黃土高原地區大量徑流小區的觀測資料,主要包括不同植被類型及枯落層、結皮層相對于開墾裸地的減蝕率(表1),分別確定喬木冠層、灌木層、草被層、枯枝落葉層、結皮層的水土保持作用系數,建立結構化植被因子指數Cs模型。
于2013年、2014年在延河流域從南到北3個植被帶各選擇2個典型小流域共6個小流域(圖1中A,B,C,D,E,F)為研究區域,選取了廣泛分布且是自然演替各階段的代表性植物群落以及該區主要分布的人工喬木林和灌木叢,共12種植物群落類型[34]。植被調查:樣方大小為喬木10 m×10 m、灌木5 m×5 m、草本2 m×2 m,至少3人以目測法目估樣方中各覆蓋層的蓋度,取平均值為該覆蓋層蓋度;采用侵蝕針法測定12種不同群落類型的土壤侵蝕模數。
侵蝕針法的具體測定如下:于雨季前(4月中旬),在每個群落內從左至右設置3個侵蝕針監測小區(小區大小:2 m×2 m),在每個監測小區內按50 cm×50 cm的網格布設侵蝕測針,每個小區9根侵蝕測針,布設侵蝕針時采用手壓或鐵錘敲擊方式,將鐵釘沿坡面垂直方向潛入土中,以鐵釘頂部與坡面平行為準,在每根侵蝕針右側5 cm處插一根紅色木簽,以標記侵蝕針的位置,在布設時盡可能不干擾監測小區內的土壤表層狀況,布設侵蝕測針的同時,采用環刀法采集土樣,測定各監測小區的土壤容重;雨季結束后(10月下旬),對每支侵蝕針上、下、左、右4個方向的侵蝕深或淤積深進行測量,取其平均值作為該測定點的侵蝕深或淤積深,侵蝕針出露地表代表被測定點在整個試驗期間為侵蝕點;侵蝕針被覆蓋代表被測定點為沉積點;侵蝕針的侵蝕深為0時代表被測定點在降雨過程中侵蝕與沉積平衡[34]。
采用侵蝕針法計算監測小區的土壤侵蝕量,計算公式如下[35]:
A=d×ρs×103
(2)
式中:A為土壤侵蝕量(t/km2);d為土壤侵蝕深(mm);ρs為土壤容重(g/cm3)。
因布設侵蝕針的樣地有部分受到人為干擾與破壞,導致部分監測小區未獲得監測數據,并剔除測量誤差較大的小區,最終獲得有效觀測數據的小區共有111個,包括遼東櫟+紫丁香(Syringapekinensis)+大披針薹草(Carexlanceolata)群落10個,三角槭+紫丁香+大披針薹草群落9個,刺槐+鐵桿蒿群落17個,紫丁香+虎榛子+鐵桿蒿+大披針薹草群落6個,狼牙刺+鐵桿蒿6個,檸條+鐵桿蒿6個,沙棘+鐵桿蒿14個,白羊草群落9個,鐵桿蒿群落9個,長芒草群落15個,達烏里胡枝子群落5個,豬毛蒿(Salsolaruthenica)+狗尾草(Setariaviridis)群落5個。
2 結果與分析
2.1 群落不同覆蓋層水土保持作用系數的確定
植物群落各覆蓋層水土保持作用系數的確定,是建立結構化植被因子指數的關鍵。由于各覆蓋層次在空間上分布及其控制水土流失機理的差異導致每個層次在群落控制水土流失效益中的作用有所差別。
本研究以文獻資料[6,8,13-14,22-33](表1)中對主要植被類型數年的徑流小區(以開墾裸地為對照)監測資料為依據,確定喬、灌、草等各層作用系數。根據表1中的數據,草本群落的平均減蝕率為63.9%,枯落層平均減蝕率為47.9%,結皮層平均減蝕率為27.2%。灌木林的平均減蝕率為98.0%,灌木林由灌木層和草本層構成,灌木層減蝕率用灌木林減蝕率減去草本層減蝕率,為34.0%。喬木林的平均減蝕率為97.9%,喬木林小區一般為刺槐林小區,由喬木層、草本層、枯落層組成,但林下草本一般較為稀疏,本研究以草本層蓋度占50%計算林下草本層減蝕率,草本層減蝕率為32.0%;林下枯落層覆蓋度和厚度均較大,本研究以覆蓋度100%計算林下枯落層減蝕率,枯落層減蝕率為47.9%;因此,喬木層減蝕率為18.1%。對結構完整的喬灌草群落,喬木層、灌木層、草本層、枯枝落葉層、結皮層的減蝕比例為18.1∶34.0∶63.9∶47.9∶27.2,各層作用系數分別為0.094 5,0.177 9,0.334 5,0.250 6,0.142 4;對缺少灌木層的喬木群落,喬木層、草本層、枯枝落葉層、結皮層的減蝕比例為18.1∶63.9∶47.9∶27.2,各層作用系數分別為0.115 0,0.406 9,0.304 8,0.173 2;對灌草群落,灌木層、草本層、枯枝落葉層、結皮層的減蝕比例為34.0∶63.9∶47.9∶27.2,各層作用系數分別為0.196 5,0.369 4,0.276 8,0.157 3;對草本群落,草本層、枯枝落葉層、結皮層的減蝕比例為63.9∶47.9∶27.2,各層作用系數分別為0.459 8,0.344 5,0.195 7(表2)。

表2 不同植被類型的結構化植被因子指數模型
注:C1為喬木冠層,C2為灌層,C3為草被層,C4為枯落層,C5為結皮層。
2.2 植物群落結構化植被因子指數的確定
2013年延河流域遭遇“百年一遇”的暴雨侵襲,出現了全流域有氣象記錄(1945年)以來范圍最廣、強度最大、持續時間長、月雨量最多的強降雨過程。根據陜西省水利廳網站的數據,安塞站5—10月總降雨量達1 071.8 mm,侵蝕性降雨量(≥12 mm的降雨,下同)836.3 mm;寶塔站5—10月總降雨量達1 406.8 mm,侵蝕性降雨量1 220.4 mm。2014年降雨量仍然遠高于延河流域多年平均降雨量,根據陜西省水利廳網站的數據,安塞站5—10月總降雨量達942.5 mm,侵蝕性降雨量752.9 mm;寶塔站5—10月總降雨量達1 119.6 mm,侵蝕性降雨量894.8 mm。相比于2013年、2014年總降雨量和侵蝕性降雨量均減小。
通過對12個不同植物群落Cs值的分析,并結合2013年和2014年不同群落土壤侵蝕模數數據發現,所有群落類型都呈現出隨著Cs增大,侵蝕強度減小的趨勢(圖2)。在2013年,自然演替早期群落及人工喬、灌群落的土壤侵蝕強度都不同程度的大于1 000 t/(km2·a)(黃土高原地區的允許土壤流失量[36]);自然演替中、后期群落及自然喬、灌群落隨著Cs增大,土壤侵蝕強度小于1 000 t/(km2·a)。2014年監測的不同群落土壤侵蝕強度小于2013年,但演替早期的豬毛蒿+狗尾草群落和人工喬、灌群落的侵蝕強度依然大于1 000 t/(km2·a);自然演替中、后期群落及自然喬、灌群落表現出與2013年一致的規律,但土壤侵蝕強度小于1 000 t/(km2·a)時的群落Cs值低于2013年。
在2013年特大暴雨年份,豬毛蒿+狗尾草群落、達烏里胡枝子群落、鐵桿蒿群落、人工檸條林、人工刺槐林的減蝕能力較弱,但當以上群落的Cs值分別大于23.8%,15.0%,31.7%,35.7%,30.8%,且近地面的草本層或枯落層蓋度也高時(附表),土壤侵蝕強度小于2 500 t/(km2·a),為輕度侵蝕[36]。人工沙棘林及長芒草和白羊草群落的減蝕能力強于其他草本群落,當Cs分別大于34.1%,31.4%和26.2%時,土壤侵蝕強度小于2 500 t/(km2·a);當Cs分別大于41.9%,38.3%和38.6%,且草本層和枯落層蓋度較大時,土壤侵蝕強度小于1 000 t/(km2·a)。自然喬、灌群落的減蝕能力最強,遼東櫟+紫丁香+大披針苔草和紫丁香+虎榛子+鐵桿蒿+大披針苔草群落的土壤侵蝕強度小于2 500 t/(km2·a),當Cs分別大于31.0%和48.8%,且灌木層和草本層蓋度較大時,土壤侵蝕強度小于1 000 t/(km2·a);三角槭+紫丁香+大披針苔草和狼牙刺+鐵桿蒿群落Cs分別大于38.3%和38.6%時,土壤侵蝕強度小于2 500 t/(km2·a),當Cs分別大于36.6%和42.2%,且灌木層蓋度較大時,土壤侵蝕強度小于1 000 t/(km2·a)。
在2014年豐水年份,豬毛蒿+狗尾草群落、人工檸條林、人工刺槐林的減蝕能力依然較弱,但當這些群落的Cs值分別大于22.1%,35.7%,26.4%,且草本層和枯落層蓋度較大時,土壤侵蝕強度小于2 500 t/(km2·a),為輕度侵蝕[36]。人工沙棘林減蝕能力強于其他人工林和早期群落,當Cs>32.1%時,土壤侵蝕強度小于2 500 t/(km2·a);Cs>34.4%,且草本層或枯落層蓋度較大時,土壤侵蝕強度小于1 000 t/(km2·a)。演替中、后期及自然喬、灌群落的減蝕能力較強,土壤侵蝕強度均小于2 500 t/(km2·a),當達烏里胡枝子、長芒草、鐵桿蒿、白羊草、狼牙刺+鐵桿蒿、紫丁香+虎榛子+鐵桿蒿+大披針苔草、遼東櫟+紫丁香+大披針苔草、三角槭+紫丁香+大披針苔草群落Cs分別大于19.2%,31.4%,34.6%,36.6%,36.1%,42.2%,21.4%,36.0%,且灌木層或草本層蓋度較大時,土壤侵蝕強度小于1 000 t/(km2·a)。
2.3 結構化植被因子指數與土壤侵蝕的關系
Cs與土壤侵蝕之間有極顯著的負相關關系(p<0.000 1),對不同群落Cs值與土壤侵蝕模數的關系進行擬合,發現兩者之間有明顯的指數關系、對數關系和二次多項式關系(表3),其中指數函數的擬合程度最高,圖3顯示了Cs與土壤侵蝕強度的指數關系。
3 討 論
植物群落的結構化植被因子指數Cs是根據植被的不同垂直層在水土保持作用方面的差異提出的概念,更多考慮了群落結構差異對土壤侵蝕的影響[12]。當群落結構趨于復雜時,群落的減蝕能力就愈明顯。在本研究建立的Cs模型中,林冠層的水土保持作用系數最小,草本層和枯落層最高,其次為灌木層,這與Wen[11]等和雷婉寧[12]建立的Cs模型中不同垂直層次的水土保持作用系數變化趨勢一致。在黃土高原,土壤侵蝕主要在暴雨期,林冠層的防蝕貢獻較小,森林的防蝕能力主要取決于林地枯枝落葉層的數量和質量[37]。喬木層蓋度再高,但要是林下結構不完整將會導致喬木群落的水土保持功能急劇下降,“遠看綠油油,近看黃水流”的現象正是有些植被蓋度較高而水土流失依然嚴重的表現,如本研究中人工刺槐林減蝕能力較弱,就算有再高的Cs值,土壤侵蝕強度也高于黃土高原地區的允許土壤流失量。植被的垂直分層結構對于減輕土壤侵蝕有重要作用,當植物群落的垂直層次結構越完備,其減少土壤侵蝕的作用將更加明顯[38],只有結構完整的植物群落才具有最好的水土保持作用,結構化植被因子指數正是綜合考慮了群落的垂直結構組成和各層次蓋度,才更能準確地表達植被的水土保持作用。
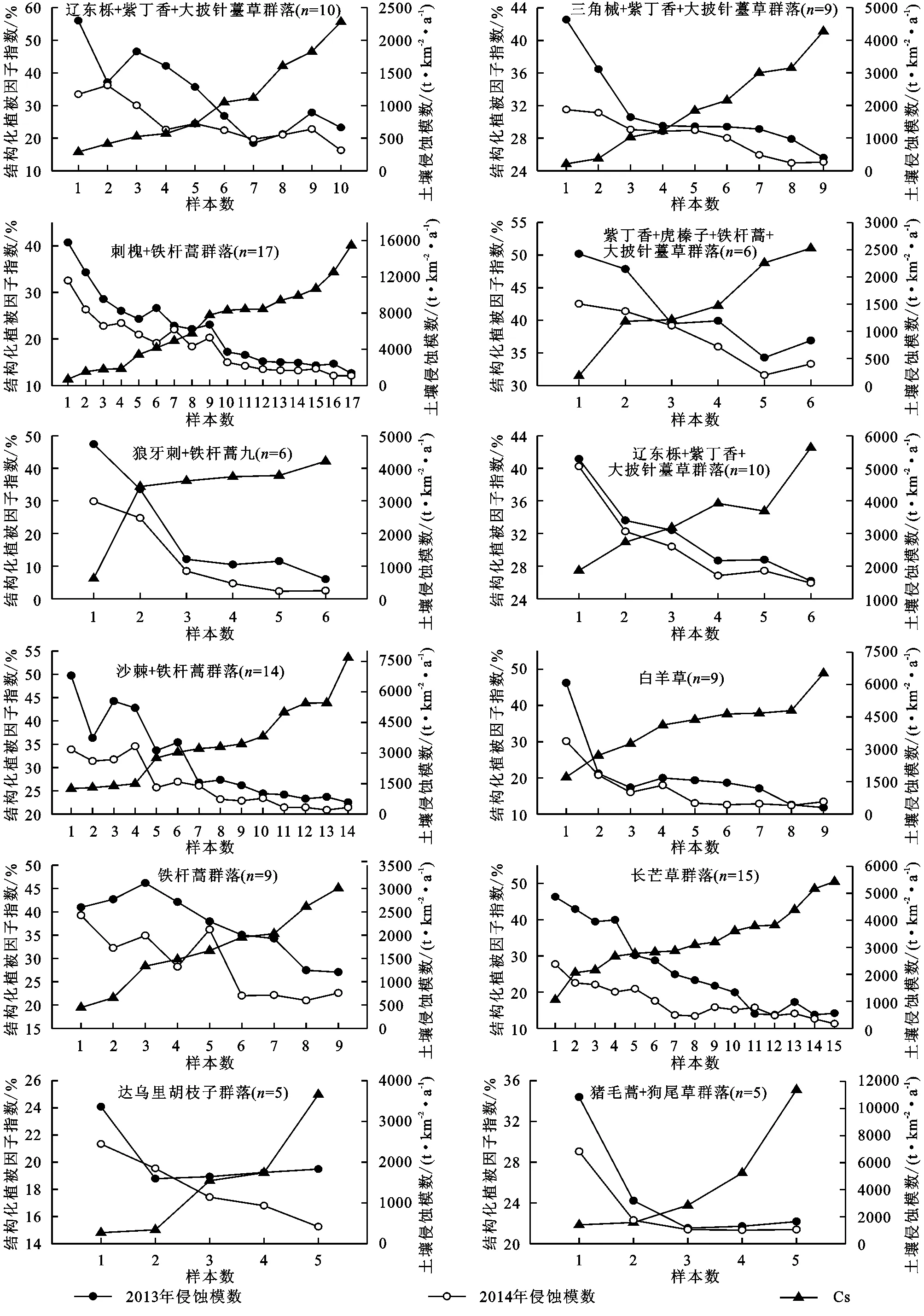
圖2 不同群落的結構化植被因子指數Cs與2013年、2014年土壤侵蝕模數

圖3 群落的結構化植被因子指數Cs與侵蝕強度的關系
不同群落類型減少土壤侵蝕的能力不同,有研究發現演替中、后期草本群落的土壤抗蝕性優于人工林[39],對于自然植被恢復而言,隨植被恢復的推進,植被對地表的防護功能、土壤的持水性能和滲透性能不斷加強[40]。本研究中,演替不同階段的草本群落,人工林,自然恢復的喬、灌群落有各自能有效減少土壤侵蝕強度的適宜Cs值。演替早期群落及人工林群落垂直結構簡單,不能有效減少土壤侵蝕。這是由于演替早期群落物種組成簡單,本身植被覆蓋較低,也不能有效改善土壤結構;而人工林則是由于缺乏林下植被層,特別是貼近地面層[41-42],因此這些群落減蝕能力較差。
演替中、后期的達烏里胡枝子和鐵桿蒿群落隨著Cs增大,能有效減少土壤侵蝕,但在暴雨年份土壤侵蝕強度高于黃土高原地區的允許土壤流失量;而演替中、后期的長芒草、白羊草群落能有效控制土壤侵蝕,隨著Cs增大,在暴雨年份土壤侵蝕強度也小于黃土高原地區的允許土壤流失量,這與禾本科植物根系有密切聯系,禾本科植物屬須根系植物,須根系多分布在土壤淺層,大量細小的毛根交織形成的網對根周土體具有加筋作用,可發揮其對土壤的網絡固持作用,有效提高土壤抗蝕性[43-44]。演替中、后期的草本群落物種組成復雜,不同植物在群落中占有各自適宜的空間資源,群落具有垂直結構層,而且由于貼近地表,可以有效攔截雨滴,減少雨滴數量、滴濺數量和減蝕量;另外,經過長時間的植被恢復,土壤表層結構得到改善,對降雨的入滲和通透能力顯著提高[37,45]。自然喬、灌群落垂直結構復雜,Cs值也較高,能有效控制土壤侵蝕,主要是因為天然林發育時間長,已經形成了比較完整且相對穩定的群落結構,群落各垂直層次可分別起到截留、蓄積滯留降水及促進入滲的作用,而且喬、灌群落根系發達,在土壤團聚體形成過程中根系的影響極為突出,根系的作用使土壤團聚體的整體性增強,提高了土壤的抗侵蝕性[40,46]。
擬合Cs與土壤侵蝕模數的關系發現,兩者有明顯的指數關系,同時對數函數和二次多項式的擬合也很顯著,這與許多學者的研究結果一致,如植被蓋度與土壤流失量之間存在明顯的指數關系,隨著群落蓋度的增大,土壤侵蝕量呈指數下降[47],林地侵蝕量和林地覆蓋度之間存在二次多項式關系[28]。但也有研究發現林草地覆蓋度與侵蝕量之間呈倒數關系[48],在小流域尺度上森林植被覆蓋度與土壤侵蝕量之間存在冪函數關系[49],而土壤侵蝕量與植被覆蓋度之間的指數關系似乎更為多數學者認可[50-51]。
4 結 論
本研究依據文獻資料的數據分析,推算出植物群落各覆蓋層水土保持作用系數,建立了不同植被類型的結構化植被因子指數Cs模型。在建立的模型中,喬木冠層的水保作用系數最小,貼近地面的草本層和枯落層水保作用系數最大。在不同水文年中,演替早期群落及人工林均未表現出較強的減蝕能力,演替中、后期群落及自然喬、灌群落則能有效減少土壤侵蝕。
在特大暴雨年份,當長芒草、白羊草群落Cs>38.3%,且草本層和枯落層蓋度較大;自然喬、灌群落Cs分別大于31.0%~36.6%,42.2%~48.8%,且灌木層和草本層蓋度較大時,土壤侵蝕強度小于1 000 t/(km2·a)。在豐水年份,當演替中后期群落和自然喬、灌群落Cs分別大于19.2%~36.6%和36.14%~42.2%,21.4%~36.0%,且灌木層或草本層蓋度較大時,土壤侵蝕強度小于1 000 t/(km2·a)。因此,不同群落其垂直結構完整,且近地面的灌木、草本層及貼地面的枯落層中至少有一層蓋度較大時,土壤侵蝕強度小于黃土高原地區的允許土壤流失量。
通過該模型計算出不同群落的Cs值,分析不同群落Cs值及土壤侵蝕強度數據發現,Cs與土壤侵蝕強度之間存在極顯著的負相關關系,對兩者進行擬合,發現兩者之間有明顯的指數、對數和二次多項式關系。
