低鹽發酵香腸用優良菌株的分離篩選及鑒定
2018-09-23 03:27:40張一凡李昌輝李平蘭張金蘭胡錦蓉
肉類研究 2018年8期
關鍵詞:生長
張一凡,李昌輝,李平蘭,張金蘭,胡錦蓉,張 瑩*
(中國農業大學食品科學與營養工程學院,北京食品營養與人類健康高精尖創新中心,北京 100083)
發酵香腸是指將絞碎的肉(常指豬肉或牛肉)和動物脂肪同糖、鹽、發酵劑和香辛料等混合后灌進腸衣,經過微生物發酵而制成的具有穩定微生物特性和典型發酵香味,并且具有較長保質期的發酵肉制品[1]。肉制品發酵劑在發酵香腸生產過程中具有理想的代謝活性[2],能夠產生有助于風味和產品發酵成熟所需的特殊酶系[3],有利于人體消化[4],且能夠抑制腐敗微生物的生長,延長產品貨架期[5]。肉制品發酵劑的研究始于20世紀初,最早發現乳酸桿菌和過氧化氫酶陽性球菌混合發酵香腸的效果較好[6]。乳酸菌可以產生特殊酶系,有助于香腸風味的產生和發酵肉制品的成熟[7],降低香腸pH值,抑制腐敗微生物的生長[8];而葡萄球菌屬具有分解脂肪、蛋白質以及產生過氧化氫酶的活性[9],能夠豐富發酵香腸的風味。因此,乳酸菌和非致病性葡萄球菌混合菌種發酵劑的研究和應用獲得了快速發展。
近年來,肉制品發酵劑研究熱點主要集中在兩方面。一是對發酵肉制品成熟過程中風味物質形成有利的呈香菌的研究,如篩選肉制品中的優良產香葡萄球菌、戊糖片球菌等[10]。Yan Cheng等[11]研究自然風干過程中腐生葡萄球菌S25發酵的香腸與天然發酵(對照)四川香腸的脂肪和蛋白質含量變化,發現腐生葡萄球菌S25可以加速四川香腸中的蛋白質水解和脂肪分解;張曉瓊等[12]從國內外發酵肉樣品中分離篩選得到10 株性能優良的葡萄球菌菌株,并對其蛋白酶、脂肪酶、硝酸還原酶和過氧化氫酶活性以及其在高鹽和亞硝酸鹽環境中的生長情況進行測定。而另一個研究熱點是能夠提高發酵香腸衛生安全性的菌株,如產細菌素能力強、可抑制病原菌生長、提高發酵肉制品安全性的乳酸菌[13]。Kantachote等[14]以戊糖片球菌HN8和Lactobacillus namurensis NH2混合發酵劑制作泰國發酵豬肉香腸,結果表明,混合發酵劑可降低香腸中生物胺和膽固醇的含量;Ba等[15]利用從天然發酵肉中篩選出的植物乳桿菌制作發酵香腸,結果表明,接種植物乳桿菌的香腸腐敗細菌數量和脂肪氧化水平顯著降低。
食鹽(NaCl)在傳統西式香腸制品的加工中發揮著重要作用,決定著發酵香腸制品的感官質量和生物安全性[16]。食鹽不但能夠促進肌原纖維蛋白的溶解,增加蛋白質的結合特性,改善香腸質地,而且還能提高發酵香腸的微生物安全性[17]。但是傳統西式香腸制品的鹽分含量很高,約為8%[18],高于大部分肉制品,長期食用可能會對人體健康造成不利影響。近年來,隨著消費者保健意識的增強,開發低鹽西式發酵香腸將更為符合消費者的需求。但鹽量的降低可能會對發酵香腸的質量安全和感官品質產生不利影響,因此需要定向篩選適用于低鹽發酵香腸的優良菌種。
本研究旨在從我國傳統自然發酵食品中分離出適用于低鹽發酵香腸的優良葡萄球菌與乳酸菌菌株,并通過測定葡萄球菌菌株的溶血、耐硝耐酸、硝酸鹽還原性和48 h產酸等發酵特性指標,以及產氨、蛋白酶、脂肪酶、產乙偶姻等風味指標,篩選出具有產香功能、可改善低鹽對風味產生的不良影響的優良葡萄球菌菌株;通過測定對乳酸菌菌株的產接觸酶、耐硝、48 h產酸等發酵特性指標,以及抑菌指標和產氨、產蛋白酶等風味指標,篩選出產酸快、抑菌能力強、可以在低鹽的基礎上保障香腸安全性的優良乳酸菌菌株。對篩選得到的優良葡萄球菌和乳酸菌菌株進行拮抗性實驗,得到無拮抗作用的菌株組合,作為潛在開發肉制品發酵劑的出發菌株。最終篩選出的混合發酵劑可在保障安全性的同時改善低鹽發酵香腸的風味,不但有利于低鹽發酵香腸產品的開發,而且能夠為我國自主產權發酵劑的研發提供理論依據。
1 材料與方法
1.1 材料與試劑
農家煙熏老臘肉、川味土豬臘腸 四川省閬中市蜀香坊特產臘味店;郭三酸菜 沈陽市成懿醬腌菜廠;歐力波蘭薩拉米 北京利康德利肉食品有限公司;川味臘肉衡東縣鄉間農家腌臘制品食品有限公司;廣味掛腸 北京家樂福中關村店;紫光園自制酸乳 北京紫光園飯店(西直門店);秦烹小院自制酸乳 北京秦烹小院;漁芙南湘菜館自制酸乳 北京漁芙南湘菜館;自制乳酸菌飲料 一口酸牛奶(北京鼓樓東大街店)。
MRS培養基、營養肉湯(NB)培養基、蛋白胨、牛肉膏、酵母浸膏 北京奧博星生物技術有限責任公司;MSA培養基 青島高科技工業園海博生物技術有限公司;氯化鈉、亞硝酸鈉、硝酸鉀、檸檬酸二銨、乙酸鈉、碳酸鈣(均為分析純) 西隴化工股份有限公司;細菌基因組DNA提取試劑盒、2×Taq PCR MasterMix 天根生化科技有限公司。
1.2 儀器與設備
SE602F電子天平 奧豪斯儀器(上海)有限公司;YXQ-LS-SⅡ全自動立式電熱壓力蒸汽滅菌器 上海博迅醫療生物儀器股份有限公司;MQD-S3R恒溫振蕩箱上海旻泉儀器有限公司;DK-8B電熱恒溫水槽 上海精宏實驗設備有限公司;SCL-1300垂直流潔凈工作臺 北京賽伯樂實驗儀器有限公司;MINIB-100恒溫金屬浴杭州米歐儀器有限公司。
1.3 方法
1.3.1 菌株分離與純化
無菌操作下稱取10 g絞碎的樣品,轉移至100 mL無菌生理鹽水中,振蕩(200 r/m,30 min)混勻;靜置數分鐘,再取1 mL加入到9 mL無菌生理鹽水中,即成10-2稀釋度,根據需要再依次稀釋;選取合適稀釋度的樣品溶液,倒入相應的固體培養基中,混勻后37 ℃靜置培養24~48 h;挑取單菌落反復劃線,直至獲得純的菌株。其中,乳酸菌的分離采用含有3% CaCO3的MRS固體培養基,挑取具有溶鈣圈[19]的菌株;葡萄球菌的分離采用MSA固體培養基。
1.3.2 菌株的初步篩選
對純化后的菌株進行革蘭氏染色、菌體形態觀察,選擇革蘭氏陽性菌種保存。對所分離的革蘭氏陽性菌分別進行如下篩選:
接觸酶實驗[20]:將37 ℃培養過夜后的菌液用接種環涂抹于已滴有5% H2O2的玻片上,立即觀察結果,3 min內出現氣泡者為陽性反應,反之為陰性。
耐硝性實驗[21]:以1%的接種量將菌種分別接種于不同亞硝酸鈉添加量(0、50、100、150 mg/kg)的液體培養基中,37 ℃培養48 h,于600 nm波長處測定吸光度。
產酸能力測定[21]:以1%接種量將菌株接種于液體培養基中,37 ℃培養24 、48 h后測定發酵液的pH值。
對于所分離的陽性球菌菌株,分別進行如下額外指標的篩選:
耐酸性實驗[21]:以1%的接種量將菌種分別接種于NB液體培養基(pH值調至5.0)中,37 ℃培養48 h,于600 nm波長處測定吸光度。
溶血實驗[22]:用接種針取37 ℃培養過夜后的菌液,在血平板上劃線,37 ℃培養24、48、72 h后觀察結果,有溶血圈出現的菌株為陽性,以金黃色葡萄球菌作為陽性對照。
硝酸鹽還原酶活力測定[23]:檢查37 ℃培養過夜后菌液的硝酸鹽還原情況,在白色搪瓷比色盤中加入1 滴菌液,然后加入硝酸還原試劑A、B液各1 滴,觀察菌落周圍出現的紅圈大小和清晰度,顯色反應后,挑出紅圈直徑>1.2 cm的菌株。
1.3.3 菌株的復篩
對初步篩選所得到的葡萄球菌及乳酸菌菌株進行進一步的篩選:
蛋白酶活性檢測[21]:將37 ℃培養過夜后的新鮮菌液分別點接種在脫脂牛乳平板培養基(SM)上,培養5 d。觀察菌落周圍透明圈的大小。
氨基酸脫羧酶實驗[24]:將37 ℃培養過夜后的菌液傾注到氨基酸脫羧酶檢測培養基(分別含有0.5%酪氨酸、0.25%組氨酸、賴氨酸及精氨酸,以不添加氨基酸作為空白對照)中,37 ℃培養,變色則為陽性。
精氨酸雙水解酶實驗[25]:將37 ℃培養過夜后的菌液傾注到精氨酸雙水解酶檢測培養基(含有1% L-精氨酸鹽)中,37 ℃培養,培養基轉化為紅色者為陽性。
對于初步篩選得到的葡萄球菌菌株,分別進行如下額外指標的篩選:
脂肪酶活性檢測[21]:將37 ℃培養過夜后的新鮮菌液分別點接種在三丁酸甘油酯培養基(TB)上,培養5 d,觀察菌落周圍透明圈。
乙偶姻產生實驗[25]:接種葡萄球菌菌株于乙偶姻檢測培養基,取培養液和40% NaOH等量混合。加少許肌酸,如果培養液10 min出現紅色即為陽性反應。
對于所分離的乳酸菌菌株,進行抑菌實驗[26]作為額外的篩選指標:將乳酸菌菌株培養至約為108CFU/mL,點接種于已含有106CFU/mL的敏感指示菌(大腸桿菌、金黃色葡萄球菌、單增李斯特菌以及蠟樣芽孢桿菌)的固體平板上,37 ℃培養12 h,觀察菌落周圍是否存在抑菌圈。
1.3.4 菌株的分類學鑒定
形態學觀察:將菌株平板劃線于固體培養基上,觀察菌落形態,挑取對數生長期的菌體進行革蘭氏染色后顯微鏡下觀察菌體細胞形態。
生理生化實驗:參照《乳酸細菌現代研究實驗技術》[20]及《常見細菌系統鑒定手冊》[25]對篩選菌株的生理生化特性進行檢測,包括精氨酸雙水解酶實驗、糖醇發酵實驗等。
16S rDNA分子生物學鑒定:細菌基因組DNA按照試劑盒說明書步驟進行提取。以提取基因組作為PCR擴增的模板,利用通用引物Lpw57(5’-AGT TTG ATC CTG GCT CAG-3’)及Lpw205(5’-CTT GTT ACG ACT TCA CCC-3’)進行PCR擴增[27]。PCR反應體系(25 μL)為:10×Buffer 2.5 μL、模板DNA 1 μL、引物各2.5 μL、dNTP 2 μL、Taq DNA聚合酶0.25 μL、dd H2O 14.25 μL,混勻5 min。PCR反應條件:94 ℃預變性5 min,94 ℃ 1 min、43 ℃ 30 s、72 ℃ 90 s,進行30 個循環,最后72 ℃延伸10 min[28]。PCR產物經瓊脂糖電泳分析后由北京諾賽基因組研究中心測序。
系統發育樹的構建:所得菌株序列根據NCBI網站(https://www.ncbi.nlm.nih.gov/)的BLAST程序在GenBank數據庫中進行比對鑒定,根據鑒定結果及相似性較高的幾株菌的16S rRNA,利用MEGA軟件進行菌株系統發育樹的構建。
1.3.5 菌株的生長特性及產酸能力測定
將菌株接種于相應的液體培養基,37 ℃培養(葡萄球菌接種在NB液體培養基中200 r/min振蕩培養,乳酸菌接種在MRS液體培養基中靜置培養),以液體培養基為空白對照,每2 h用紫外分光光度計于600 nm波長處測定吸光度,用酸度計測定菌液的pH值。
1.3.6 菌株間拮抗性測試
用含乳酸菌的接種環,在低鹽模擬肉湯固體培養基[29](NaCl質量分數2%)上劃一直線接菌。37 ℃條件下培養48 h,待形成菌落后,沿菌落邊緣(不接觸)用接種環從垂直方向接種葡萄球菌;37 ℃條件下培養24 h,觀察乳酸菌對于葡萄球菌的抑制效果。
1.4 數據處理
所有實驗均重復進行3 次,使用SPSS 17.0軟件對數據進行統計和方差分析,使用Excel軟件對數據進行分析及作圖。
2 結果與分析
2.1 優良葡萄球菌菌株初篩結果
從5 種自然發酵食品中分離出平板培養形態不同的疑似菌株共56 株,其中6 株來自川味臘肉,4 株來自歐力波蘭薩拉米,1 株來自廣味掛腸,15 株來自農家煙熏老臘肉,30 株來自川味土豬臘腸。從疑似菌株中篩選出革蘭氏陽性球菌、接觸酶陽性菌39 株,初步判定為葡萄球菌。
對所得的39 株菌株的安全性進行溶血實驗,結果表明,39 株菌均為溶血實驗陰性,說明分離的球菌均具有安全性。39 株菌株中,所有菌株均可耐受50、100 mg/kg NaNO2,僅2 株球菌對150 mg/kg NaNO2不耐受;其中34 株菌具有硝酸鹽還原酶,且其中的23 株可耐受pH 5.0的酸度,經過初篩,得到發酵特性良好的優良葡萄球菌23 株,作為下一步復篩的出發菌株。其中菌株L1、L2、L3來自川味臘肉,R1、R2、R3來自歐力波蘭薩拉米,Z7、Z9、Z10、Z12來自農家煙熏老臘肉,C6、C7、C8、C10、C11、C16、C17、C23、C24、C25、C26、C29、C30來自川味土豬臘腸。
2.2 優良葡萄球菌菌株復篩結果
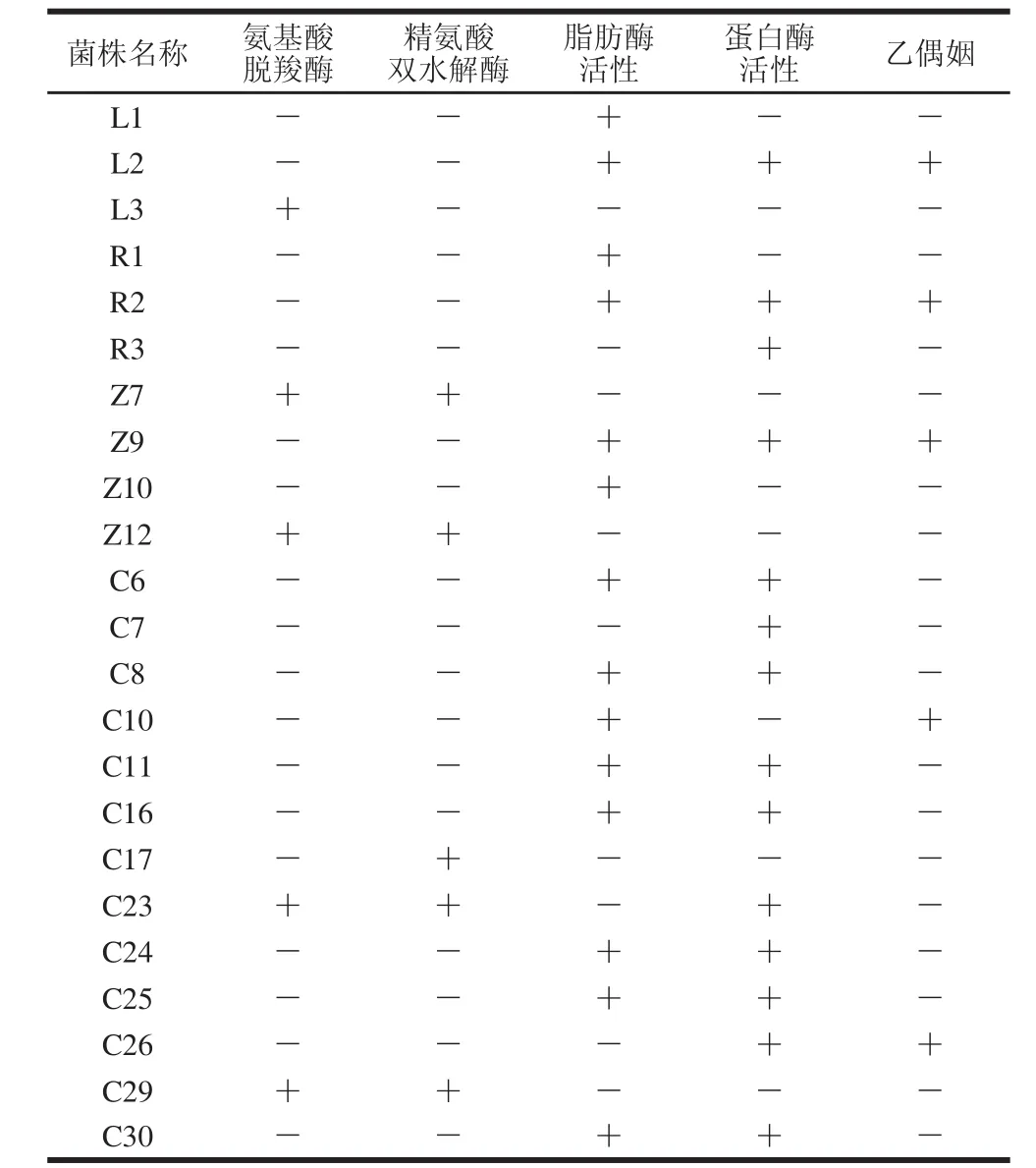
表1 葡萄球菌菌株復篩結果Table 1 Secondary screening of Staphylocooccccuuss s t raaiinnss
對初篩所得的23 株葡萄球菌進行復篩,由表1可知,其中有17 株菌既不產氨基酸脫羧酶,也不產精氨酸雙水解酶,這17 株菌中有15 株菌具有脂肪酶活性,14 株菌具有蛋白酶活性,而既產蛋白酶又產脂肪酶的菌株有12 株,其中有3 株產乙偶姻。因此,最終篩選出3 株具有優良風味特性的葡萄球菌,它們分別為篩選自川味臘肉的L2、篩選自歐力波蘭薩拉米的R2以及篩選自農家煙熏老臘肉的Z9。
2.3 優良乳酸菌菌株初篩結果
從6 種自然發酵食品中分離出的平板培養形態不同的疑似菌共52 株,其中25 株來自郭三酸菜,7 株來自歐力波蘭薩拉米,5 株來自北京紫光園自制酸乳,4 株來自秦烹小院自制酸乳,7 株來自漁夫南湘菜館自制酸乳,3 株來自一口酸牛奶(鼓樓東大街店)的自制乳酸菌飲料。從疑似菌株中篩選出具溶鈣圈的革蘭氏陽性、接觸酶陰性乳酸菌30 株。
所得30 株乳酸菌菌株中,所有菌株均可耐受50、100 mg/kg NaNO2,其中有28 株菌可耐受150 mg/kg亞硝酸鹽。不同菌株的產酸情況差異較大,部分產酸較快菌株在MRS培養基中培養24 h,pH值可達3.67,其中21 株菌的pH值可在24 h內降至3.70~3.90,產酸效果較好。經過初篩,得到發酵特性良好的優良乳酸菌21 株,作為下一步復篩的出發菌株。其中菌株P1、P2、P4、P5、P6、P8、P9、P12、P15、P19、P20、P21來自于郭三酸菜,S1來自于歐力波蘭薩拉米,SN1-1、SN1-2、SN1-3、SN1-5來自于北京紫光園自制酸乳,SN2-6、SN2-7、SN2-8來自于秦烹小院自制酸乳,X來自于一口酸牛奶自制乳酸菌飲料。
2.4 優良乳酸菌菌株復篩結果
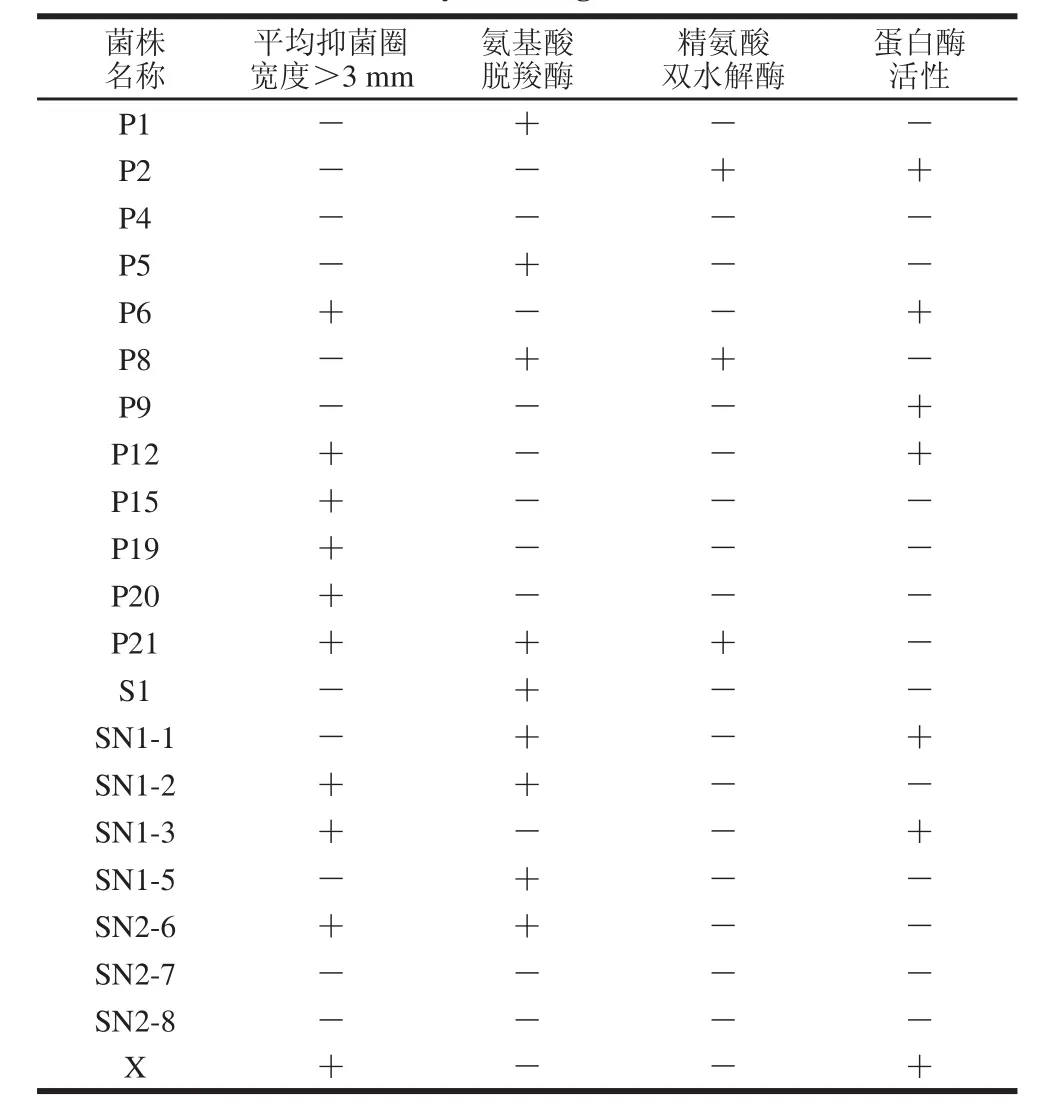
表2 乳酸菌菌株復篩結果Table 2 Secondary screening of lactic acid bacteria
對初篩所得的21 株乳酸菌進行復篩,經過抑菌實驗,所有菌株均對大腸埃希氏菌、金黃色葡萄球菌、單增李斯特菌、蠟樣芽胞桿菌有抑制作用。由表2可知,根據抑菌圈的大小,其中10 株乳酸菌的抑菌作用最強(抑菌圈平均寬度大于3 mm)。在進行復篩的21 株菌中,氨基酸脫羧酶陰性的菌株有12 株,精氨酸雙水解酶陰性的菌株有18 株,具有蛋白酶活性有7 株。因此,最終篩選出4 株兼具多種優良特性的乳酸菌菌株,分別為篩選自郭三酸菜的P6與P12、篩選自北京紫光園自制酸乳的SN1-3以及篩選自一口酸牛奶自制乳酸菌飲料的X。
2.5 菌株的分類學鑒定結果
2.5.1 菌株形態學鑒定結果
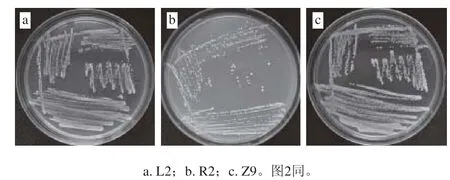
圖1 葡萄球菌菌落形態特征Fig. 1 Colony characteristics of Staphylococcus
由圖1可知,葡萄球菌菌株L2、R2及Z9在NB固體培養基上37 ℃培養24 h,菌落直徑約1~2 mm,菌落形態均呈白色圓形,不透明,形狀規則,其中L2、R2菌落光滑、濕潤、呈凸起狀,Z9菌落邊緣粗糙,質地干燥且菌落比較扁平。

圖2 葡萄球菌菌體形態學特征(×1 000)Fig. 2 Morphological characteristics of Staphylococcus (×1 000)
由圖2可知,經革蘭氏染色后在顯微鏡下觀察,葡萄球菌菌株L2、R2及Z9的菌體細胞均為圓球形,革蘭氏陽性,其中菌株L2部分成對生長,菌株R2單個生長,菌株Z9呈不規則堆狀排列。
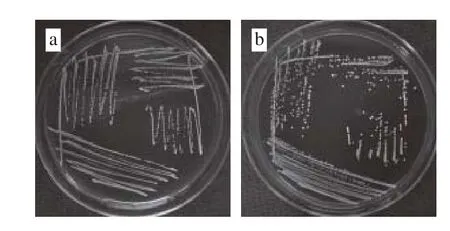
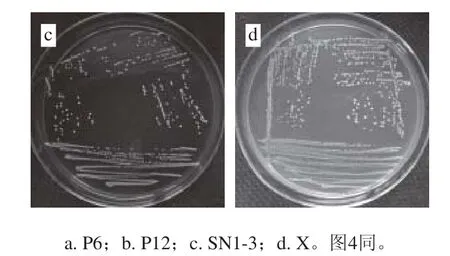
圖3 乳酸菌菌落形態特征Fig. 3 Colony characteristics of lactic acid bacteria
由圖3可知,乳酸菌菌株P6、P12、SN1-3及X在MRS固體培養基上37 ℃培養24 h,菌落直徑約1~2 mm,菌落形態均為白色圓形,菌落表面光滑,濕潤凸起,邊緣規則,其中X菌落呈半透明狀,P6、P12、SN1-3菌落均不透明。
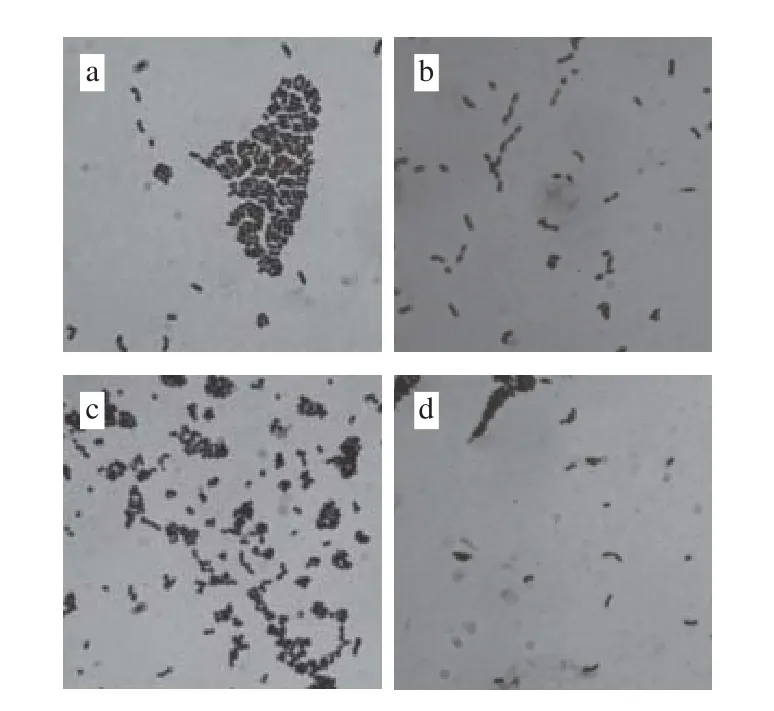
圖4 乳酸菌菌體形態學特征(×1 000)Fig. 4 Morphological characteristics of lactic acid bacteria (×1 000)
由圖4可知,經革蘭氏染色后在顯微鏡下觀察,乳酸菌菌株P6細胞成對或成短鏈,革蘭氏陽性,粗短桿狀;菌株P12成對或成短鏈,為革蘭氏陽性短桿菌;菌株SN1-3細胞部分以直二個方向形成四聯排列,為革蘭氏陽性球菌;菌株X成對或成短鏈,為革蘭氏陽性桿菌。
2.5.2 菌株生理生化實驗鑒定結果

表3 葡萄球?菌菌株生理生化實驗結果Table 3 Physiological and biochemical characteristics of Staphylocooccccuuss
由表3可知,參考《常見細菌系統鑒定手冊》[25],根據葡萄球菌生理生化鑒定結果,初步推斷葡萄球菌菌株L2、Z9為腐生葡萄球菌,R2為肉葡萄球菌。
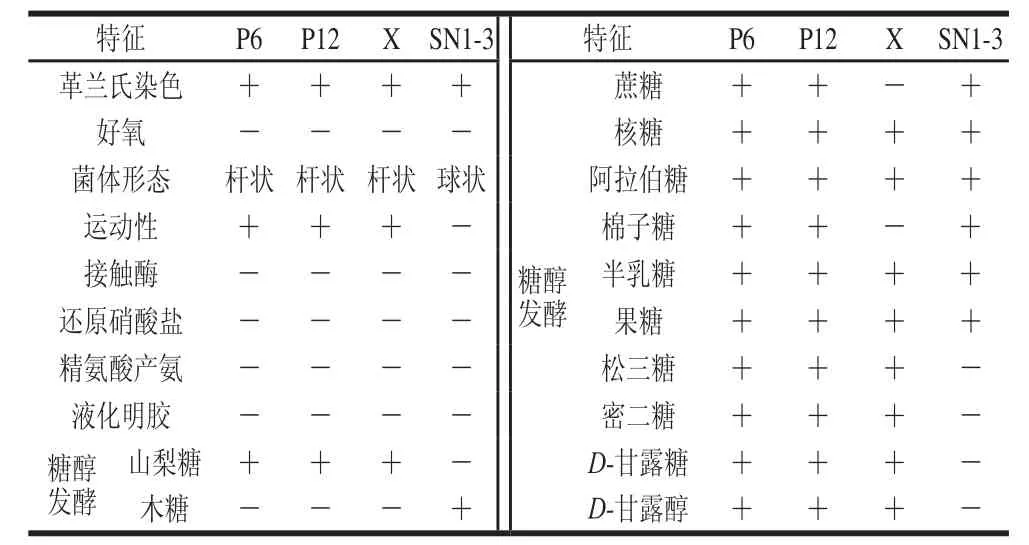
表4 乳酸菌菌株生理生化實驗結果Table 4 Physiological and biochemical characteristics of lactic acid bacteria
由表4可知,參考《常見細菌系統鑒定手冊》[25]和《乳酸細菌現代研究實驗技術》[20],根據乳酸菌菌株的生理生化鑒定結果,初步推斷乳酸菌菌株P6、P12為植物乳桿菌或鼠李糖乳桿菌、X為干酪乳桿菌、SN1-3為戊糖片球菌。
2.5.3 菌株的16S rDNA分子生物學鑒定結果
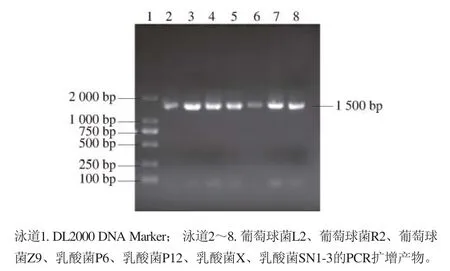
圖5 菌株16S rDNA的PCR擴增產物電泳圖Fig. 5 Electrophoresis of PCR products 16S rDNA gene from strains
由圖5可知,對菌株的16S rDNA序列采用通用引物進行擴增后,在1 000~2 000 bp之間可見到清晰明亮的條帶,大小約為1 500 bp。
利用NCBI網站的BLAST程序對菌株16S rDNA測序結果在GenBank已知序列數據庫中進行對比分析。結果表明:葡萄球菌菌株L2、Z9與腐生葡萄球菌(Staphylococcus saprophyticus)具有最高同源性,葡萄球菌菌株R2與肉葡萄球菌(Staphylococcus carnosus)具有最高同源性;乳酸菌菌株P6、P12與植物乳桿菌(Lactobacillus plantarum)具有最高同源性,乳酸菌菌株X與干酪乳桿菌(Lactobacillus casei)具有最高同源性,乳酸菌菌株SN1-3與戊糖片球菌(Pediococcus pentosaceus)具有最高同源性。選取與GenBank中相似性較高菌株的序列,利用MEGA 6.0軟件進行系統發育樹的構建,結果如圖6~8所示。

圖6 葡萄球菌菌株L2、R2、Z9的系統發育樹Fig. 6 Neighbour-joining tree of Staphylococcus L2, R2 and Z9


圖8 乳酸菌菌株SN1-3的系統發育樹Fig. 8 Neighbour-joining tree of Lactobacillus SN1-3
綜合生理生化及分子生物學鑒定,結果表明,葡萄球菌菌株L2、Z9為腐生葡萄球菌,葡萄球菌菌株R2為肉葡萄球菌;乳酸菌菌株P6、P12為植物乳桿菌,菌株X為干酪乳桿菌,SN1-3為戊糖片球菌。
2.6 菌株的生長特性及產酸能力
本研究測定了篩選出的3 株葡萄球菌及4株乳酸菌的在培養過程中的生長情況及培養基的pH值變化,并繪制曲線。

圖9 不同葡萄球菌的生長曲線及產酸能力( =3)Fig. 9 Growth curve and acid production of Staphylococcus (n = 3)
由圖9可知,葡萄球菌L2、Z9和R2均能在NB液體培養基中生長且長勢良好,這3 株菌的生長速率和生長曲線的趨勢非常相似,生長調整期均較短,12 h后進入生長穩定期,菌數達到最大值。此外,這3 株菌的pH值變化也非常相似,均隨培養時間的增加呈上升趨勢,在24 h后趨勢均逐漸平緩,與生長曲線趨勢大體一致。
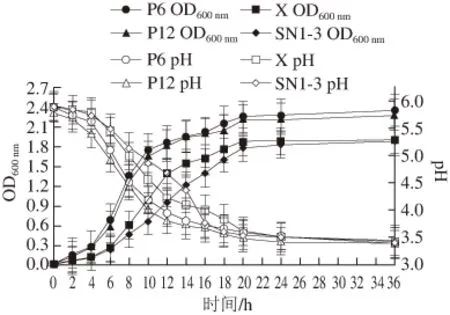
圖10 不同乳酸菌的生長曲線及產酸能力(n==44)Fig. 10 Growth curve and acid production of lactic acid bacteria (n = 4)
由圖10可知,乳酸菌P6、P12、X及SN1-3均能在MRS液體培養基中生長且長勢良好,但生長速率存在差異。乳酸菌P6、P12的生長速率較快,生長調整期較短,約為0~2 h,16 h后進入生長穩定期,菌數達到最大值。而菌株X與SN1-3生長速率較慢,生長調整期略長,約為0~4 h,20 h后才達到生長穩定期。此外,這4 株菌產酸較快,0~12 h內pH值下降迅速,36 h后均達到3.5以下,產酸能力較強。但4 株菌的產酸速率存在差異,菌株P6、P12產酸最快,而菌株SN1-3相對較慢。
2.7 菌株間的拮抗作用
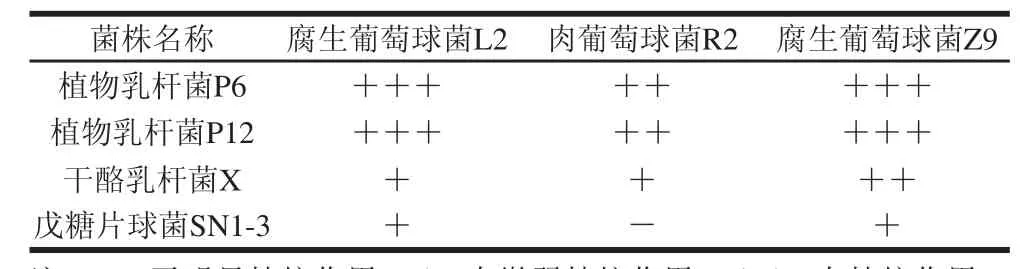
表5 菌株間拮抗性測試結果Table 5 Compatibility test among antagonistic strains
由表5可知,戊糖片球菌SN1-3對于3 種葡萄球菌的拮抗作用最微弱,對肉葡萄球菌R2無明顯拮抗作用;而植物乳桿菌P6、P12對于3 種葡萄球菌的拮抗作用最強,對腐生葡萄球菌Z9具有非常強的抑制作用,若復配使用非常不利于葡萄球菌的生長。因此,經菌株間拮抗實驗的結果,選擇戊糖片球菌SN1-3與肉葡萄球菌R2作為最終制作低鹽發酵香腸的復配菌株。
3 結 論
國內肉類科技界在產品加工過程中在質量與特性變化、加工用發酵劑菌種選育、配制等方面開展了大量工作,但是關于低鹽發酵香腸的工藝研究和菌株選育方面的研究較少。因此,定向從我國自然發酵制品中篩選利于低鹽西式發酵香腸制品生產與開發的適應性優良菌種,不但可以利用我國豐富的菌種資源,還能夠為我國低鹽發酵香腸產品的研發提供重要的理論依據。
本研究從臘肉、四川香腸、薩拉米等5 種自然發酵制品中分離、純化得到56 株過氧化氫酶陽性球菌菌株。通過菌株耐受性、發酵特性、風味特性等指標對所分離的56 株過氧化氫酶陽性菌株進行篩選,最后篩選得到腐生葡萄球菌L2、肉葡萄球菌R2以及腐生葡萄球菌Z9為適用于低鹽發酵香腸、具有優良產香特性的葡萄球菌菌株。其中L2來自川味臘肉,R2來自薩拉米,Z9來自農家煙熏老臘肉,它們均可以耐受150 mg/kg的NaNO2和較低的酸度,并且具有硝酸鹽還原能力。菌株均不產生氨基酸脫羧酶和精氨酸雙水解酶,具有一定的蛋白酶和脂肪酶活性,還可以產生風味物質乙偶姻,具有優良的產香特性。
本研究還通過菌株耐受性、發酵特性、抑菌特性、風味特性等指標對分離自泡菜、自釀酸乳、乳酸菌飲料等6 種自然發酵制品中的52 株乳酸菌菌株進行篩選,最終篩選得到植物乳桿菌P6、植物乳桿菌P12、干酪乳桿菌X以及戊糖片球菌SN1-3 4 株適用于低鹽發酵香腸的優良乳酸菌菌株。其中,菌株P6、P12來自酸菜,SN1-3來自酸乳,X來自乳酸菌飲料。4 株菌均可耐受150 mg/kg的NaNO2,具有很好的發酵特性,產酸速度快,產酸能力強,不含有氨基酸脫羧酶和精氨酸雙水解酶,可以抑制金黃色葡萄球菌、大腸桿菌、單增李斯特菌、蠟樣芽胞桿菌等食源性致病菌,并且具有較好的蛋白酶活性,有利于保障低鹽發酵香腸的安全性及風味。
葡萄球菌與乳酸菌復配作為香腸發酵劑,可以在產香的同時保證低鹽發酵香腸的安全性,但是乳酸菌產生的細菌素以及乳酸會對葡萄球菌的生長造成不利影響,因此混合發酵劑的篩選必須考慮到菌株間的拮抗作用。本研究經過葡萄球菌與乳酸菌的拮抗實驗,發現戊糖片球菌SN1-3和肉葡萄球菌R2之間無明顯的拮抗作用,可以用于香腸混合發酵劑的開發。因此,后續實驗也將進一步利用SN1-3+R2復配組合制作低鹽發酵香腸,考察其混合發酵的效果,為我國自主產權低鹽發酵香腸混合發酵劑的研究和開發提供參考。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
