膠東地區凹唇壁蜂種群調查及限制因子分析
2018-09-21 07:46:12劉麗李麗莉李超袁銳于毅門興元葉保華
中國蜂業 2018年9期
劉麗 李麗莉 李超 袁銳 于毅 門興元 葉保華
(1 山東農業大學植保學院,泰安 271018;2 山東省農業科學院植物保護研究所,濟南 250100)
凹唇壁蜂(Osmia excavataAlfken)屬于蜜蜂總科(apoidea)切葉蜂科(megachilidae),是我國北方的一個野生傳粉昆蟲,1990年被發現并用于給蘋果樹授粉[1,2]。由于單位面積用蜂量少、授粉效率高、不用飼喂、便于管理等特點,深受果農歡迎[3]。凹唇壁蜂已經在我國各果區大面積應用了25年[4,5],具有“傳粉之王”的稱號[6-8]。在果園內釋放壁蜂為蘋果授粉,可以顯著提高坐果率和果品質量,省工省時間,投資少,效益高[9]。但關于各地凹唇壁蜂種群狀況、限制因子的比較研究報道較少。為此,本文對從山東省膠東地區的蘋果主栽區棲霞、牟平收集的蘆葦蜂管、棲霞塑料管進行壁蜂種群調查,并對限制壁蜂種群的因子進行分析,以期更好的發揮凹唇壁蜂的傳粉增產功能。
1 材料與方法
1.1 研究區域概況
山東省為中國農業大省,是我國北方水果的主產區。目前山東省的凹唇壁蜂存蜂量全國最多,約5億頭,每年為近200萬畝蘋果、梨等果樹授粉,其中膠東半島在上個世紀90年代開始應用凹唇壁蜂進行授粉,是我國最早應用壁蜂授粉的地區。
1.2 調查方法
2018年1月從膠東半島的牟平、棲霞市的蘋果園收集凹唇壁蜂蜂管,開展凹唇壁蜂種群調查。在煙臺市牟平區,壁蜂主要用蘆葦管飼養,而在棲霞市,壁蜂用蘆葦管或塑料管飼養。因此,在煙臺市牟平區收集了蘆葦管蜂管(MP),在棲霞市收集了蘆葦管蜂管(QX1)和塑料管蜂管(QX2)。
從各調查點的蘋果種植戶收集凹唇壁蜂蜂管,隨機抽查100管。用小刀將蜂管從中間剖開,記錄蜂管內巢室數量、蜂繭數量、完整花粉團數量、空室數量、蜂螨危害巢室數、寄生蜂危害巢室數,然后稱量管中蜂繭的重量,用游標卡尺測量蜂繭的長度,最后用鑷子撕開蜂繭,檢查凹唇壁蜂的雌雄。
導致壁蜂死亡的作用因子按邏輯順序依次排列為:(1)卵期:敗育。凹唇壁蜂在每個巢室產的花粉團上產一粒卵,由于無法直接觀察蜂管內卵的發育情況,因此將巢室內花粉團完整的情況,表示沒有壁蜂幼蟲取食,定為卵敗育。(2)幼蟲和蛹期:螨寄生、蜂捕食、自然死亡。通過田間取樣剖查蜂管,觀察到巢室中螨寄生導致幼蟲死亡(定為“螨寄生”)和其他蜂的幼蟲取食凹唇壁蜂幼蟲(定為“蜂捕食”),剖查的巢室,花粉粒已經被取食,幼蟲、蛹已經死亡,但是沒有天敵捕食和寄生的跡象,定為由于氣候、環境等物理因子引起的“自然死亡”。
1.3 數據處理
應用SPSS軟件對數據進行方差分析,其中不同地點的壁蜂種群參數間的顯著性差異采用Duncan新復極差法進行分析,相關分析采用Pearson相關分析。
2 結果與分析
2.1 膠東地區壁蜂種群比較
2個采集點不同材質蜂管的壁蜂每管巢室數(F=23.551,df=2,P=0.000)、繭數(F=6.778,df=2,P=0.001)均有顯著差異,而成繭率(F=1.336,df=2,P=0.264)無顯著性差異(表1)。膠東地區壁蜂巢室數為8.71~10.71個,其中QX2的巢室數最低,為8.71個,顯著低于QX1和MP的巢室數,與QX1和MP的巢室數具有顯著性差異。QX2的每管繭數最低,為6.38個,顯著低于QX1和MP。巢室成繭率QX1最高,達75.40%,但與MP、QX2無顯著差異。
2個地點凹唇壁蜂種群的卵敗育率(F=4.182,df=2,P=0.016)、自然死亡率(F=3.559,df=2,P=0.030)均存在顯著性差異,被其他蜂捕食率(F=1.862,df=2,P=0.157)、被螨寄生率(F=2.495,df=2,P=0.084)無顯著性差異(表2)。卵敗育率在4.15%~9.70%,其中QX2的卵未孵化率顯著低于MP。被捕食率在6.05%~8.69%,棲霞和牟平之間沒有顯著性差異。QX2的被螨寄生率最高,為5.59%。MP的自然死亡率最高,達4.23%,顯著高于QX1。
2個地點凹唇壁蜂蜂繭的長度和繭重均存在顯著差異(F=6.308,df=2,P=0.003;F=17.497,df=2,P=0.000),見表3。其中,MP的繭長最大,為12.29mm,與QX具有顯著差異。MP的繭重最大,為139.29mg,QX1次之,QX2最小為90.95mg。
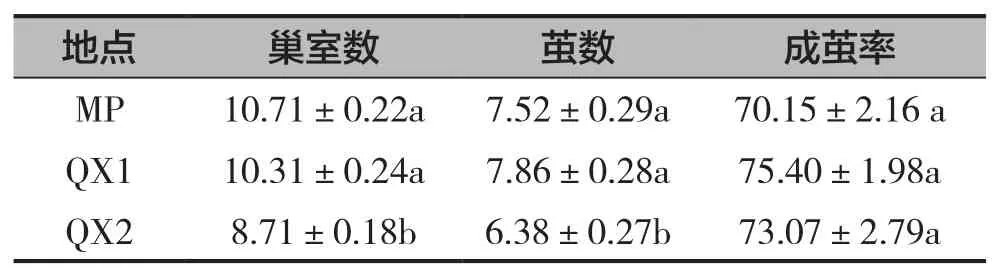
表1 膠東地區壁蜂種群參數
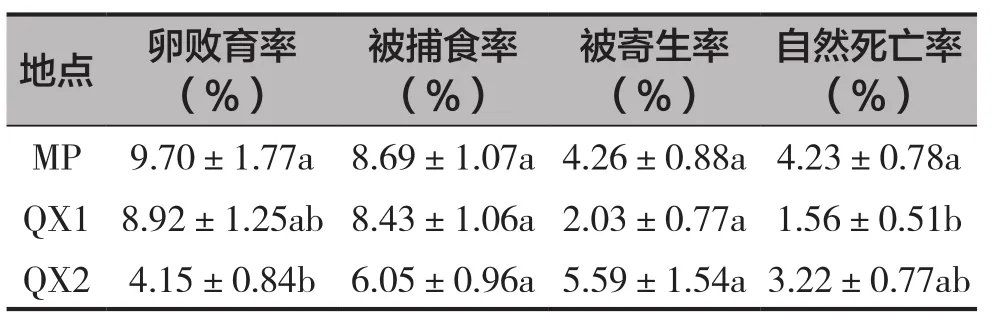
表2 作用因子對凹唇壁蜂種群的致死率
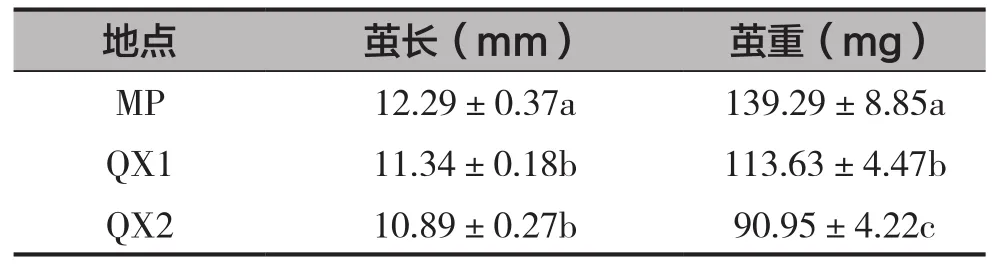
表3 不同地點凹唇壁蜂繭長和繭重
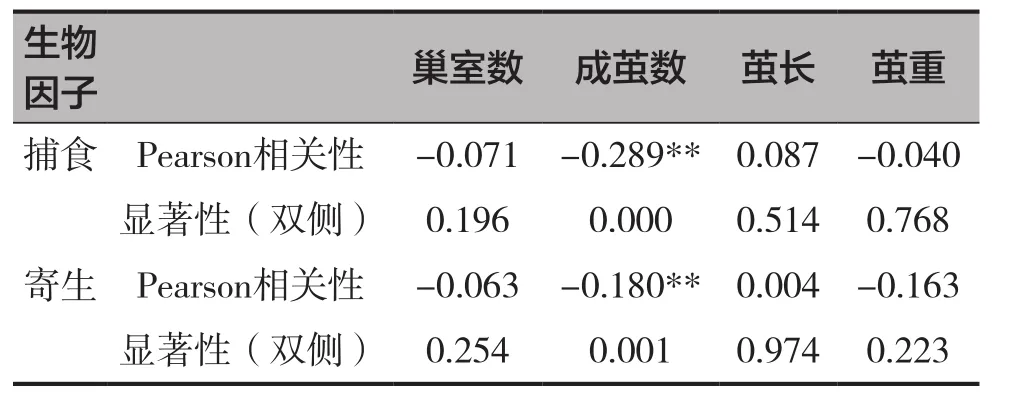
表4 被寄生率和被捕食率與巢管參數和壁蜂繭參數的相關性
2.2 蜂管天敵對壁蜂種群的影響
被螨寄生率、被捕食率均與成繭數呈顯著負相關關系(r=-0.180,P=0.001;r=-0.289,P=0.000),見表4。
2.3 導致壁蜂死亡的因子
2.3.1 寄生蜂──叉唇寡毛土蜂
叉唇寡毛土蜂(Sapyga comaYasumatsu et Sugihara)屬土蜂總科,寡毛土蜂科,寡毛土蜂屬。該蜂是1987年從日本引進角額壁蜂(Osmia cornifrons)為果樹授粉的過程中被發現,并由美國內華達大學R. W. Rust教授鑒定[10]。徐環李1993~1994年在山東調查,凹唇壁蜂被叉唇寡毛土蜂捕食率為30. 8%[11],曹衍斌等調查山東省6個地區凹唇壁蜂種群,發現壁蜂被捕食率為0.33%~6.48%,不同地區差異較大[12],而我們的結果顯示被捕食率為6.05%~8.69%。綜上所述,叉唇寡毛土蜂對凹唇壁蜂的捕食現象各地區普遍存在,不同地區的捕食率有差異,可能與其環境中叉唇寡毛土蜂的蟲源數量有關。
圖1和圖2為該雌成蜂的正腹面觀,顯示成蟲體細長,蟲體黑色,腹部有多道金色的環。雌成蜂體長10mm,觸角鞭節內側深紅色,兩觸角窩上端之間有一個黃斑,唇基兩側、中胸背板后端、中胸側板有一個黃斑。雄成蜂觸角第12節膨大,末節較短,溢縮,唇基整個為黃色[10,13]。

圖1 叉唇寡毛土蜂正面觀

圖2 叉唇寡毛土蜂腹面觀

圖3 叉唇寡毛土蜂蜂繭

圖4 凹唇壁蜂蜂繭
叉唇寡毛土蜂與壁蜂通過長期協同進化,其生活周期幾乎與寄主壁蜂生活史同步[11]。叉唇寡毛土蜂的幼蟲取食壁蜂老熟幼蟲后,在原寄主幼蟲所作的繭內化蛹并羽化為成蟲。壁蜂幼蟲被害時流出大量體液、浸蝕繭殼,變成深赤褐色,無光澤(圖3)。而正常壁蜂的繭是白色,有光澤(圖4)。因此,可以根據繭的顏色,剔除叉唇寡毛土蜂的蜂繭,減少其對凹唇壁蜂的危害。
2.3.2 捕食螨──平島氏毛爪螨
平島氏毛爪螨(Chaetodactylus hirashimai Kurosa),屬疥螨目(Sarcoptiformes)毛爪螨科(Chaetodactylidae)毛爪螨屬,是盜寄生性攜播型害螨,盜寄生,即一方從另一方處掠食,所掠食物包括另一方所殺死的、捕獲的、或準備好的貯藏食料[14]。平島氏毛爪螨通過成年蜂攜帶進入巢管中,取食巢室中的蜂糧-花粉團,快速繁殖,使壁蜂幼蟲的存活率下降[12]。雖然蜂螨不會寄生危害蜂繭和成蜂,但是其會大量附著在破繭而出的成年蜂體表,進行傳播,同時影響蜂的活動及采粉、傳粉效率[14]。該害螨經中國科學院動物研究所陳軍研究員鑒定,是我國的一個新記錄種,圖5為平島氏毛爪螨在蜂管中的為害狀,圖6為平島氏毛爪螨蟲體。
有研究表明該害螨在山東省主要蘋果產區普遍發生[12,14],各個地區的螨寄生率不同,與應用壁蜂時間有關,也與各個地點控制螨害的措施不同有關。對該害螨防治的關鍵技術是在放蜂前,徹底清除附著在壁蜂蜂繭上的螨蟲,研究顯示雙甲脒、噠螨靈、氰戊菊酯等殺蟲劑以及乳酸、次氯酸鈉、冰乙酸處理壁蜂蜂繭,都能有效除螨,同時對壁蜂相對安全[15]。
2.3.3 花斑皮蠹
我們在剝查壁蜂蜂繭的過程中,發現有少量花斑皮蠹。花斑皮蠹(Trogoderma variabileBallion)隸屬鞘翅目(Coleoptera)皮蠹科(Dermestidae)昆蟲,是一種適應性強,危害物品廣泛的儲藏物害蟲,在國內各省(市)自治區都有分布[16],幼蟲與毛皮蠹的一些種多混合發生于鳥巢和某些蜂類巢內,取食死蜂及某些動物的干死體和皮毛等[17]。花斑皮蠹幼蟲體長7.0~8.0mm,紡錘形,黃褐色,背面略隆起,腹面近扁平,胸腹部各節前半部呈黑褐色,節間腹面及足為黃白色,全身除疏生不分節的“單毛”外,在第7、8腹節背面各生1束“槍頭”狀毛。隨齡期的增加,體色加深[16],圖7為花斑皮蠹的幼蟲。有報道,花斑皮蠹也是壁蜂的自然天敵[18],花斑皮蠹如何侵入壁蜂巢室、危害壁蜂的哪個蟲態尚未見報道,還需要深入研究。根據花斑皮蠹的生物學特點,清潔壁蜂存儲環境,并用花斑皮蠹趨避劑趨避,可以減少其危害[16]。
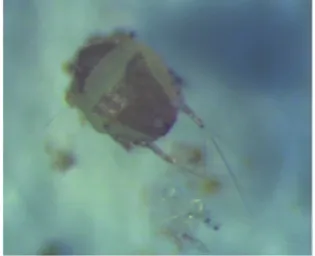
圖6 平島氏毛爪螨

圖7 花斑皮蠹幼蟲

圖8 自然死亡的凹唇壁蜂蟲體
2.3.4 自然死亡
通過田間取樣剖查蜂管,剖查的巢室中,花粉粒已經被取食,幼蟲、蛹、成蟲已經死亡,但是沒有天敵捕食和寄生的跡象,暫定為由于氣候、環境等物理因子引起的“自然死亡”。在剖管過程中,發現了身體僵硬、體色變深的凹唇壁蜂幼蟲蟲體,以蛹的狀態在蜂繭內死亡的蟲體及死亡的凹唇壁蜂成蟲,見圖8。導致壁蜂死亡的原因,還需要進一步的研究。
3 討論與結論
壁蜂(Osmia)廣泛分布于世界各地,具有活動早、耐低溫、飛行快、授粉效率高、活動范圍小的特點,被廣泛應用于我國北方蘋果、梨、桃等果樹的傳粉。1990年在棲霞觀東西山果園釋放日本引進角額壁蜂傳粉時,設巢引誘回收到一個野生傳粉優勢蜂種,經中國農科院吳燕如教授鑒定為凹唇壁蜂[19]。凹唇壁蜂具有“傳粉之王”的稱號,是我國重要的資源昆蟲。凹唇壁蜂一年發生1代,以卵、幼蟲、蛹、成蟲在管狀巢內越夏越冬,初孵幼蟲取食一個花粉團完成從幼蟲到成蜂的整個過程,不再補充營養。翌年的春季壁蜂破繭而出,營巢、采集花粉和產卵,巢外活動時間40~50天,壁蜂的生活史和生活習性非常便于果農飼養和利用。
我們系統調查了棲霞市和牟平地區的凹唇壁蜂種群,發現兩地凹唇壁蜂的成繭率均在70%以上,顯著高于曹衍斌等2015年在膠東地區的調查結果(15%~44%)[12],這可能與不同年份氣候以及蜜粉源的質量有關。我們的研究結果還顯示,蜂管的材質會影響凹唇壁蜂的營巢數量,也會影響結繭數及蜂繭質量。總的來看,蘆葦管更有利于凹唇壁蜂營巢、結繭。凹唇壁蜂在塑料管的巢室數、形成的蜂繭數均低于蘆葦管的巢室數,推測壁蜂更喜歡在蘆葦管中筑巢、結繭。從壁蜂規模化飼養角度看,雖然塑料管對壁蜂種群略有影響,但是由于塑料蜂管的成本更低、剝繭更方便,同時可以重復使用,因此塑料蜂管在生產中被普遍應用。
自然天敵是威脅壁蜂種群的重要因子,我們調查進一步明確了凹唇壁蜂的主要自然天敵是叉唇寡毛土蜂和平島氏毛爪螨,2種自然天敵普遍發生,限制壁蜂成繭。我們對2種天敵的生物學和生態學了解的還不夠,需要進一步的深入研究。我們發現除了自然天敵外,還有一些引起壁蜂死亡的“自然因子”。與2015年在山東蘋果園壁蜂種群的制約因子調查數據相比較,發現2017年壁蜂種群的自然死亡率(1.56%~4.23%)明顯低于2015年的調查結果(9.69%~79.44%),其原因還需要進一步研究明確。我們發現蘆葦管中凹唇壁蜂的蜂繭長度和質量均大于塑料管,猜測因為蘆葦管管壁較厚,凹唇壁蜂在其中受外界環境脅迫小,而塑料管管壁較薄,壁蜂在其中易受外界環境影響,消耗能量大,重量輕。
總之,凹唇壁蜂是一種優秀的本土傳粉昆蟲資源,在我國北方水果生產中發揮著重要的傳粉增產作用,我們對其種群生物學和種群限制因子的研究還有待進一步加強。
