脅迫溫度下兩株高致病性副溶血弧菌的生長差異比較分析
2018-09-20 07:27:08馬馮莉張昭寰吳佳敏孫曉紅潘迎捷
食品與生物技術學報 2018年6期
馬馮莉 , 張昭寰 , 吳佳敏 , 孫曉紅 ,2,3, 潘迎捷 ,2,3, 趙 勇 *,2,3
(1.上海海洋大學 食品學院,上海 201306;2.上海水產品加工及貯藏工程技術研究中心,上海 201306;3.農業部水產品貯藏保鮮質量安全風險評估實驗室(上海),上海 201306)
南美白對蝦是南亞和東南亞地區最為重要的水產品,常因其汁鮮肉嫩、營養豐富、價格適中,成為消費者的水產品首選,其銷售量占到全球蝦產量的90%以上,與中國對蝦、斑節對蝦并列,被稱為世界三大經濟對蝦之一[1]。然而,南美白對蝦卻攜帶很多致病菌,其中,副溶血性弧菌(Vibrio parahemolyticus,簡稱VP)被認為是首要致病菌,常引起傷口感染、敗血癥、腹瀉、頭痛和急性腸胃炎等疾病,在實際生活中,常常會因為不正當的加工操作、運輸、消費等引起副溶血性弧菌在即食蝦上滋長[2],這極大地危害了蝦的質量和公共健康,因此控制對蝦中的副溶血性弧菌對食用安全對蝦具有重大意義。VP的主要致病因子為溶血毒素,目前發現的主要有耐熱性溶血毒素(TDH);耐熱性溶血毒素相關的溶血毒素(TRH)和不耐熱溶血毒素(TLH)。而TDH是一種能消化血細胞細胞壁的酶,能在血瓊脂培養基上產生一種β-型溶血環,被稱為神奈川現象,呈現神奈川陽性 (Kanagawa phenmomenon positive,KP+),而無毒性的菌株則呈現神奈川陰性,流行病學調查研究也同樣表明副溶血性弧菌的致病力與TDH有很高的相關性。因此,TDH被認為是該致病菌的主要毒力因子[3],研究攜帶有tdh致病基因的副溶血性弧菌的生長特性在風險評估具有重大意義。
預測微生物學最初由食品預測微生物學發展而來,主要是運用微生物學、工程數學以及統計學建立數學模型,利用所建立的模型,結合計算機以及配套軟件預測以及描述微生物在不同環境條件下的生長、死亡情況,這些環境條件主要包括溫度、培養基質、水分活度、pH值以及氣體環境等等[4]。預測微生物學的主要目的是在不進行微生物檢測的條件下快速預測產品貨架期和安全性,其核心在于建立完善的數學模型。風險評估是食品安全控制的有效手段,而預測模型是進行精準評估的工具,目前,基于副溶血性弧菌的風險評估研究多集中于單一菌株在不同環境條件上建立的模型[5-6],很少有文獻探討研究關于不同來源的高致病性副溶血性弧菌的生長及其生長動力學參數是否存在差異。而作為水產品中最常見的一種致病性微生物,建立其完善的預測模型對于保障水產品質量、預測水產品貨架期具有市場意義。世界衛生組織和聯合國糧農組織在2011年發布了副溶血性弧菌在水產品中的風險評估報告,其中提到不同菌株的生長特性具有差異,即基于不同菌株建立的預測模型與風險評估的不確定性存在直接相關性,所以,建模時應當考慮菌株異質性[7]。
為了探討脅迫溫度下高致病性副溶血性弧菌的生長動力學參數差異,作者選用兩株相同致病性菌株ATCC 33847和F 18,建立4~30℃溫度條件下的生長模型和失活模型,運用多重數學評價指標確定最為適合副溶血性弧菌在南美白對蝦上的生長模型,在獲得模型參數基礎上進一步分析高致病性副溶血性弧菌在即食蝦上的生長特性,比較其生長動力學參數,為食品安全風險評估提供數據支撐。
1 材料與方法
1.1 實驗儀器與材料
高精度恒溫培養箱:日本Sanyan公司;BagMixer 400 VW型拍打式均質器:法國Interscience公司;離心機:德國 Eppendorf公司;ATCC 33847(tlh+/tdh+/trh-)、F 18(tlh+/tdh+/trh-):中國科學院微生物研究所購買,菌株儲存于-80℃冰箱內,甘油保存;TCBS培養基、TSA培養基、胰蛋白胨大豆肉湯(TSB)培養基:北京陸橋技術有限責任公司。
1.2 無菌蝦樣的制備
南美白對蝦購于上海農貿市場,置于均質袋中,貯藏于-80℃下。實驗前將南美白對蝦于4℃下解凍12 h,將解凍后的蝦樣品放入2.5 g/dL NaCl溶液中,煮沸20 min,置于生物安全柜中冷卻,挑選質量為10 g左右的蝦樣本用于后續實驗。另選取10只滅菌蝦樣,均質涂布于TSA培養基上,過夜培養,未發現有菌落生長,以證明蝦樣已去除本底微生物。
1.3 菌株懸浮液制備與接種
在凈化工作臺中分別取兩種副溶血性弧菌各1 mL 接種于 9 mL TSB(3 g/dL NaCl)中,37 ℃下搖床培養10~12 h,然后再各取100 μL活化菌液分別接種至 9 mL TSB(3 g/dL NaCl)中,37 ℃下搖床培養10~12 h。 將培養完成的菌液離心 10 min(3 000 g,15℃),去上清液,用PBS緩沖溶液調節吸光度,對于培養溫度分別為15、20、25、30℃的初始接種量采用3.6 lg cfu/g;培養溫度為4、7、10℃的初始接種量采用大約在 5.0 lg cfu/mL,吸取6 mL菌液加到500 mL 3 g/dL NaCl溶液中制備菌株懸浮液,搖勻。
在生物安全柜中將挑選出的南美白對蝦樣本放入上述菌株懸浮液中,振蕩40 min,使南美白對蝦與懸浮液充分接觸,進行接菌。接菌完成后,將蝦樣本放置在 4、7、10、15、20、25、30 ℃下進行恒溫貯藏。
1.4 微生物計數
在無菌條件下,將接菌后的蝦樣放入含有0.85 g/dL滅菌生理鹽水100 mL的無菌BagFilter均質袋中拍打3 min,用0.85 g/dL滅菌生理鹽水進行梯度稀釋,選用合適的稀釋度,取100 μL的稀釋液在TCBS瓊脂板上進行涂布,將平板在37℃條件下培養24 h計算菌落數。每個取樣點做兩個平行,每個稀釋梯度重復2次。
1.5 生長數據的擬合
1.5.1 一級模型擬合 在 10、15、20、25、30℃條件下,不同來源的副溶血性弧菌在即食南美白對蝦上的生長數據用修正Gompertz模型[8,10](式1)、Baranyi模型[11](式 2)、Logistic 模型[11](式 3)擬合。 模型表達式如下:
1)修正Gompertz模型:
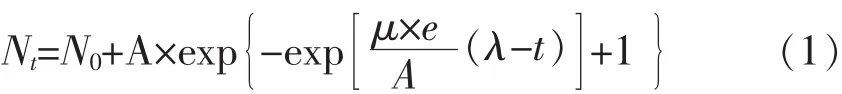
其中,N0:最初的細菌總數量,lg cfu/mL;Nt:時間為 t時的細菌總數量,lg cfu/mL;A:最大菌種量與初始菌種量之差,lg cfu/mL;λ:延滯期,h;μ:最大增長速率,lg(cfu/mL)/h。
2)修正 Baranyi模型:
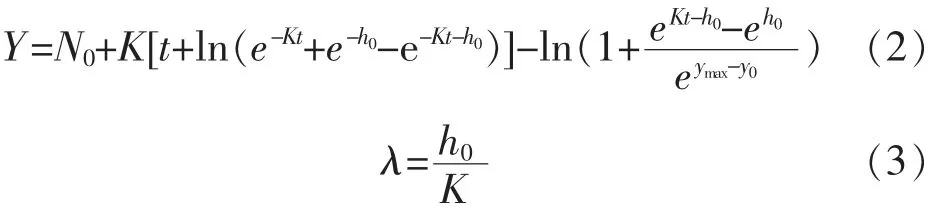
其中,N0:最初的細菌總數量,lg cfu/mL;Y:時間為 t時的細菌總數量,lg cfu/mL;h0:使模型擬合更加穩定的初始細菌量的轉換值。
3)修正Logistic模型:
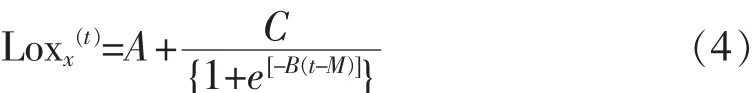
其中,x (t): 時間為 t時的細胞數量,lg cfu/mL;A:時間t降到0的漸近線數值;C:向上和向下漸近線所能得到的差值;M:絕對生長率最大的時間,h;B:當 t=M 時的相對生長率,lg(cfu/mL)/h。
1.5.2 失活模型 在4、7℃條件下,不同來源的副溶血性弧菌在即食南美白對蝦上的失活情況用Log-linear模型[10](式 5)、Weibull模型[11](式 6)和Baranyi模型(式2)進行擬合。模型表達式如下:
1)Log-linear模型:
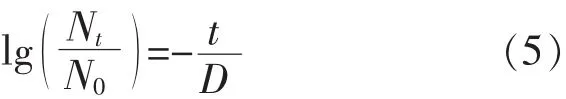
2)Weibull模型:

其中,N0:最初的細菌總數量,lg cfu/mL;Nt:時間為 t時的細菌總數量,lg cfu/mL;D:表示弧菌降低一個對數級所需要的時間,h;t:表示低溫處理時間,h;n、b:曲線形狀因子。
1.5.3 二級模型
1) Modified Ratkowsky(μmax-T) 模型
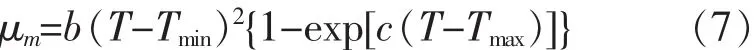
2) Non-linear Arrhenius(λ-T) 模型

其中,Tmin和Tmax分別是細菌生長/死亡的最小/最大溫度理論值,℃;A,B,C,b 和 c:線性方程回歸系數。
1.5.4 模型評價
1)相關系數R2
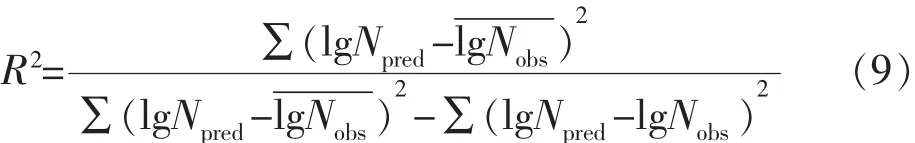
R2的值在0~1之間,且越靠近1即表明預測模型的參考價值越高[9]。
2)均方根誤差RMSE[10]
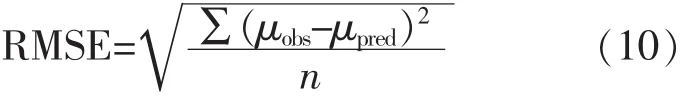
均方根誤差RMSE是說明模型預測值的離散程度的一種數值指標。
3)準確因子(Accuracy factor,Af)和偏差因子(bias factor,Bf)
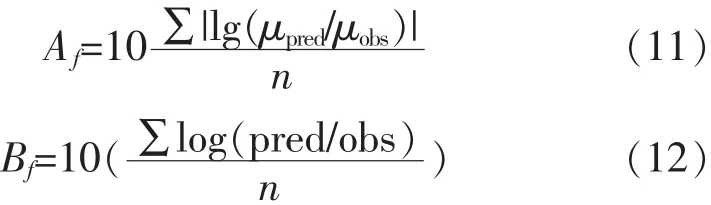
Af=1說明預測值與實驗觀測值是相等的,Af值越大表明預測模型與觀測值的平均精準度偏低;Bf主要用于判斷預測模型的偏差度,Bf=1,說明所選用模型的擬合結果與實驗觀測值相同,所選用模型的擬合效果較好[10]。式9-12中,μobs代表實驗觀測值,μpred代表模型預測值,n代表觀測值個數。
2 結果與分析
2.1 不同溫度下副溶血性弧菌的生長差異分析
如圖1所示,ATCC 33847和F 18在 20~30℃條件下屬于生長狀態,副溶血性弧菌的數量隨著培養時間延長而增長,兩株菌的生長類似,但在4~15℃,兩株菌的生長呈現明顯差異。從20℃到30℃,ATCC 33847和 F 18的最大生長量(Nmax)均為9 lg cfu/mL以上,兩者的生長速率沒有明顯差異。高溫條件下研究高致病性VP時,溫度對其生長特性的影響比其他因素要大,所以VP的生長特性相似。
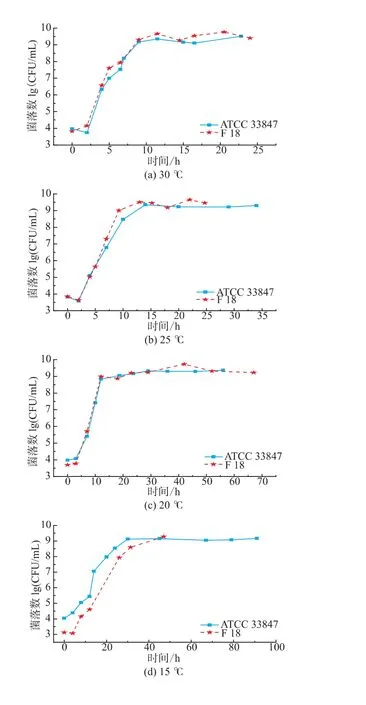
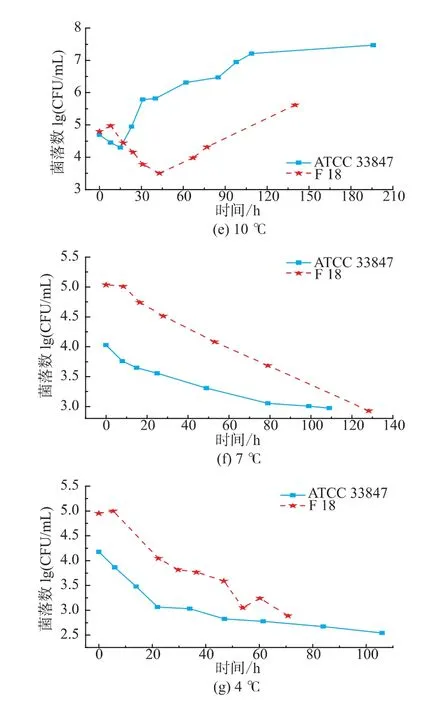
圖1 ATCC 33847和F 18在4~30℃下的生長曲線Fig.1 Growth curves of ATCC 33847 and F 18 at 4~30 ℃
在15℃時,F 18的生長比ATCC 33847的緩慢,兩株菌初現生長差異。在10℃條件下,ATCC 33847和F 18在即食南美白對蝦上的生長特性呈現了明顯的差異,ATCC 33847一直處于緩慢的生長期,Nmax約為7.4 lg cfu/mL,而F 18在前40小時處于死亡狀態,菌落數隨著時間的延長不斷減少,但是在40 h之后,菌落數隨著時間的延長而增長,在培養時間達到140 h時,菌落數大約為5.5 lg cfu/mL,超過了初始接種量4.75 lg cfu/mL,這表明,F 18在10℃時呈現了較明顯的異質性,其原因有待于進一步研究。在4~7℃條件下,副溶血性弧菌的菌落數量都在逐步減少。在7℃時,ATCC 33847和F 18最少生長量(Nmin)均降到2.9 lg cfu/mL左右,但是在培養期間,兩者的生長特性具有一定的差異。F 18的數量隨著培養時間延長,呈現緩慢減少的趨勢,但是ATCC 33847的數量下降比較快,且在培養前期(0~10 h),F 18的菌落數量基本保持不變,而ATCC 33847則下降了0.5 lg cfu/mL。在4℃,高致病性VP的生長特性相差較大,F 18菌落數減少速度在20~45 h時比較小,且隨著培養時間延長,菌落數存在一個反復現象,而ATCC 33847則一直處于減少狀態,在0~20 h期間菌落數急劇下降,之后呈現緩慢減少狀態。
由本研究的生長特性可知,副溶血性弧菌在4~15℃條件下的生長存在較大的異質性,這表明與高溫條件相比,脅迫溫度下兩株高致病性VP的生長特性受溫度影響比較小,其他因素對其影響比溫度大,如菌株來源不同,部分菌株會呈現較大的異質性。
2.2 不同溫度條件下兩株致病性副溶血性弧菌的一級模型比較
為了探討高致病性VP在不同溫度條件下的生長差異,采用修正Gompertz模型、修正Logistic模型和Baranyi模型三個一級模型來擬合兩株致病性VP在10~30℃下的生長數據,使用Weibull模型、Baranyi模型和Log-linear模型三個一級模型來擬合兩株致病性VP在4~7℃下的死亡數據,并得到了相關參數,結果見表1。
從25~30℃,ATCC 33847和F 18在即食南美白對蝦上的生長數據均可以用所采用的三個模型進行擬合,且擬合效果良好,修正決定系數R2adj均在0.924以上。F 18在15℃和20℃條件下的生長數據無法用Logistic模型進行擬合,R2為0.800和0.223,而ATCC 33847在該溫度的擬合效果良好。而且,在10℃條件下,所選用的三個模型均無法擬合F 18的生長數據,但是ATCC 33847的擬合效果良好。同樣,Baranyi模型對ATCC 33847在4℃時的生長數據擬合效果較好,但是對F 18的擬合效果較差,R2分別為0.935和0.390。
在模型擬合中,參數RMSE的值越大表明模型的預測值的離散程度越大,Af值越大表明預測模型與觀測值的平均精準度偏低。同樣,Bf值偏離1越遠,表明擬合結果與實驗觀測值相差越遠,表明所選模型不佳。所以,綜合表1中的擬合參數,通過比較 R2adj、RMSE、Af和 Bf值, 選擇修正 Gompertz模型作為最適模型來擬合兩株致病性副溶血性弧菌ATCC 33847和F 18在15~30℃條件下的生長數據以及10℃下ATCC 33847的生長數據,選用Baranyi模型來描述7℃條件下兩株菌以及4℃條件下ATCC 33847的生長數據,但是F 18在10℃和4℃條件下的生長模型有待于進一步研究。在15~30℃時,唐等[12]考慮到初始菌量的擬合正確性,選擇Baranyi模型為不同基因型VP在12℃和35℃時的最適模型,但也指出Modified Logistic和Gompertz的擬合度高于Baranyi模型。在本研究中,綜合比較Modified Gompertz和Baranyi模型的擬合參數,作者選擇Modified Gompertz為最適模型用來描述VP在15~30℃的生長行為,這表明預測模型在選擇時具有一定的綜合性。這兩株致病菌生長數據的擬合效果同樣表明,高溫條件下,高致病性VP的生長異質性較小;而脅迫溫度下高致病性VP的異質性較大,在最適模型的選擇上存在一定的差異。即在研究致病性VP時不能籠統的選擇模型,需要經過一定的比較、分析以及對模型進行一定的修正。
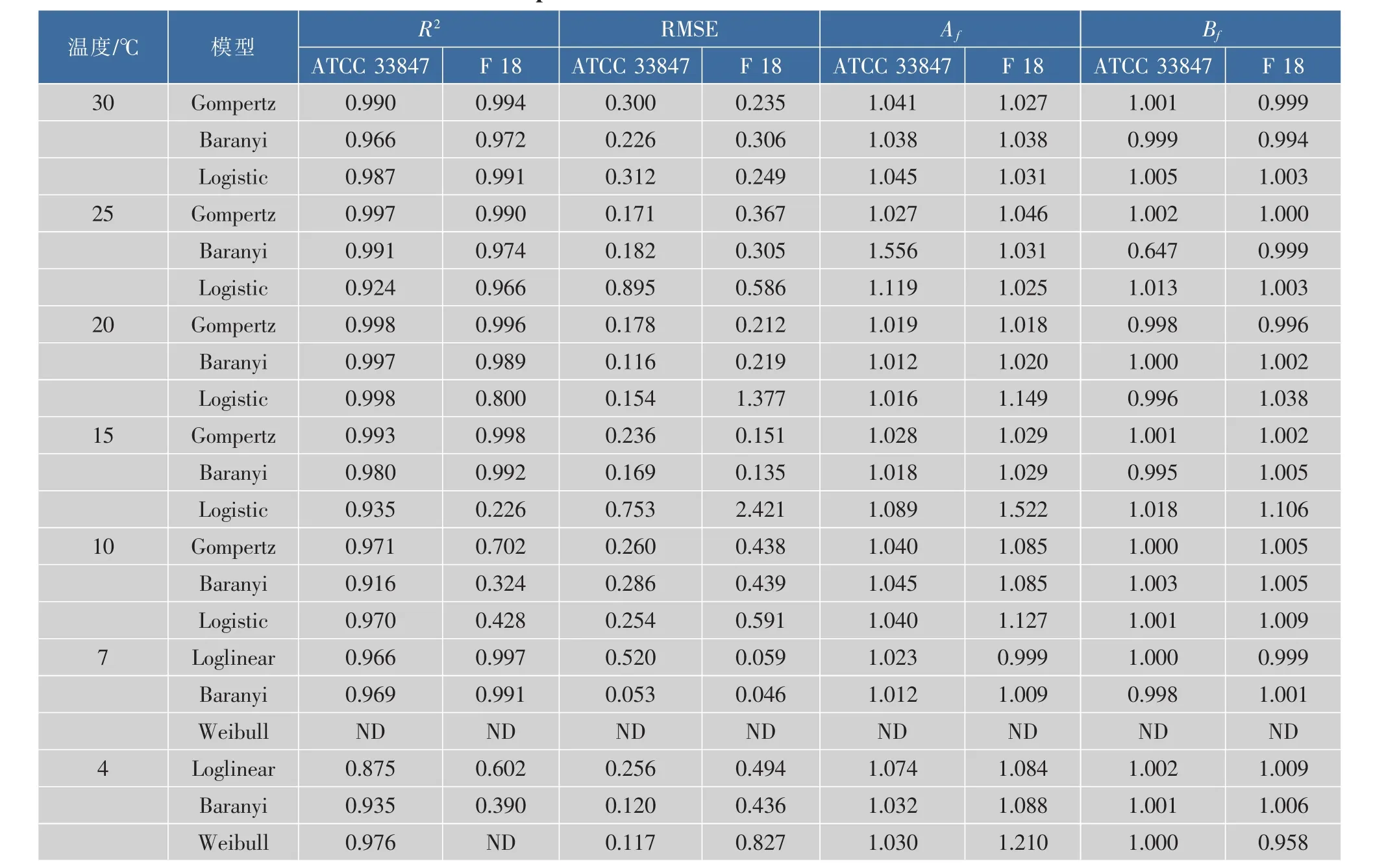
表1 ATCC 33847和F 18在4~30℃條件下的動力學生長參數Table 1 Growth parameters of ATCC 33847and F 18 at 4~30 ℃
2.3 不同溫度條件下兩株致病性副溶血性弧菌的二級模型建立及其與同類研究做比較
基于最適模型擬合得到動力學參數最大比生長速率(μmax)和延滯期(LT),根據公式 7-8 建立二級模型見圖2。隨著溫度的升高,最大比生長速率(μmax)逐漸變大,延滯期(LT)逐漸減小。從 15 ℃升至30℃過程中,F 18的μmax始終大于ATCC 33847,兩者的差值隨著溫度的升高而變大,在15℃時約為0.16 lg ((cfu/mL)/h),30 ℃時約為 0.7 lg((cfu/mL)/h);F 18的LT始終小于ACTT 33847,兩者的差值隨著溫度的升高而減小,在15℃時約為7.3 h,最小差值約為2.4 h,見表2。這表明,在生長過程中,不同溫度條件下,相同致病性VP的μmax以及LT存在一定的差異。
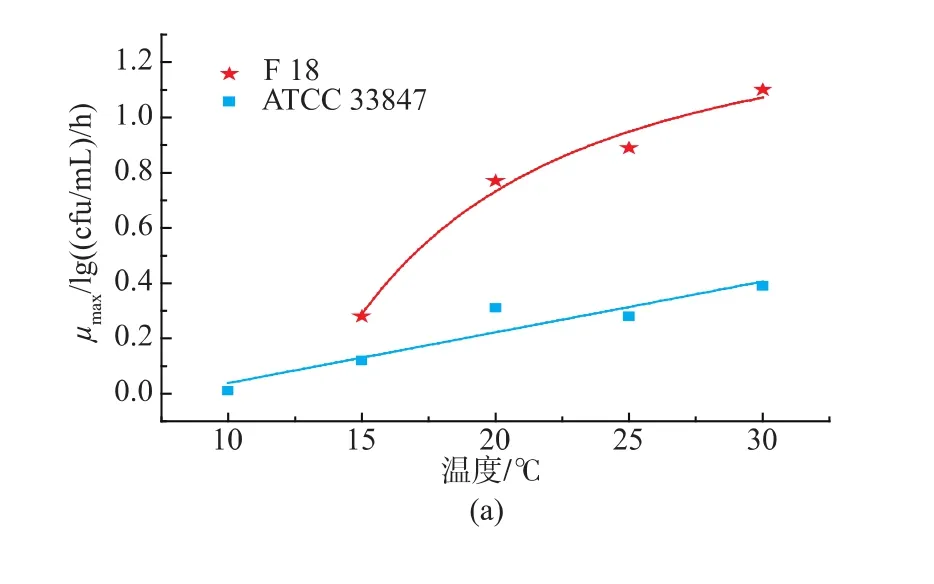

圖2 ATCC 33847和F 18的最大比生長速率(μmax)和延滯期(LT)的二級模型Fig.2 Secondary model for the effect of temperature(℃)on the specific maximum growth rate (μmax) and lagtime(LT) of ATCC 33847 and F 18
將本研究中的副溶血性弧菌在10~30℃的最大比生長速率與同類文獻建立二級模型作比較,參考文獻見表3,結果見圖3。可以看出,雖然攜帶相同的tdh致病基因,但是相較于其他三個菌株,只有ATCC 33847的生長數據可以在10℃被模型較好的擬合得到μmax值,其他三個菌株的生長數據均無法被擬合;在20~30℃,VPD43的μmax值始終保持最大,依次為 F 18,O3:K6,最小的為 ATCC 33847,而且差值隨著溫度的升高而變大,這表明VPD43在南美白對蝦上的生長最快,而ATCC 33847最慢,即相同基因型的VP在相同基質上的生長速率存在顯著性差異(P<0.05)。可能有很多原因引起這一現象,Sant等[13]在2012年研究沙門氏菌和單增李斯特菌在即食生菜上的生長表明,菌株之間的差異會引起生長差異。同樣,Carlin等[14]在2013年研究芽孢桿菌7大親緣菌體的生長時也發現了相同菌株存在生長差異。所以攜帶有相同致病基因的菌株在生長上存在異質性。
在VP微生物風險評估研究中建立預測微生物模型時,因選用的菌株來源不同,會使得VP的生長呈現較大的生長差異;不同研究中所選用的一級模型,同樣會對最大生長量、初始接菌量、生長速率等參數造成一定的偏差。通過本研究可以看出,高溫條件下,相同高致病性VP在即食蝦上的生長特性具有相似性,但是生長參數存在差異(P<0.05);在脅迫溫度下,相同致病性VP存在較大的生長特性差異,其生長參數存在顯著性差異(P<0.05)。因此,在研究致病性VP的機理以及進行風險評估時,需要考慮菌株異質性所導致的影響,某一菌株所建立的模型應用于另一菌株時,需要進行修正或者直接建立新的模型,以便更準確的預測菌株生長的風險性。
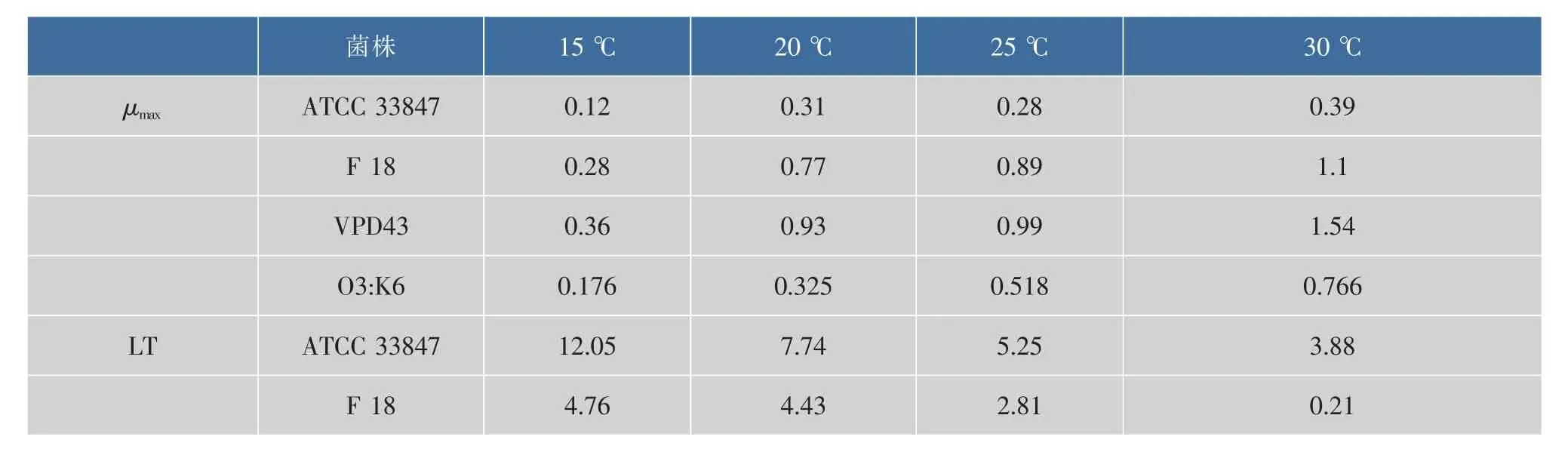
表2 副溶血性弧菌在不同溫度下的 μmax(lg(cfu/mL)/h)和 LT(h)值Table 2 μmax(Log10(cfu/mL)/h)and LT(h) values of Vibrio parahaemolyticus at different temperatures
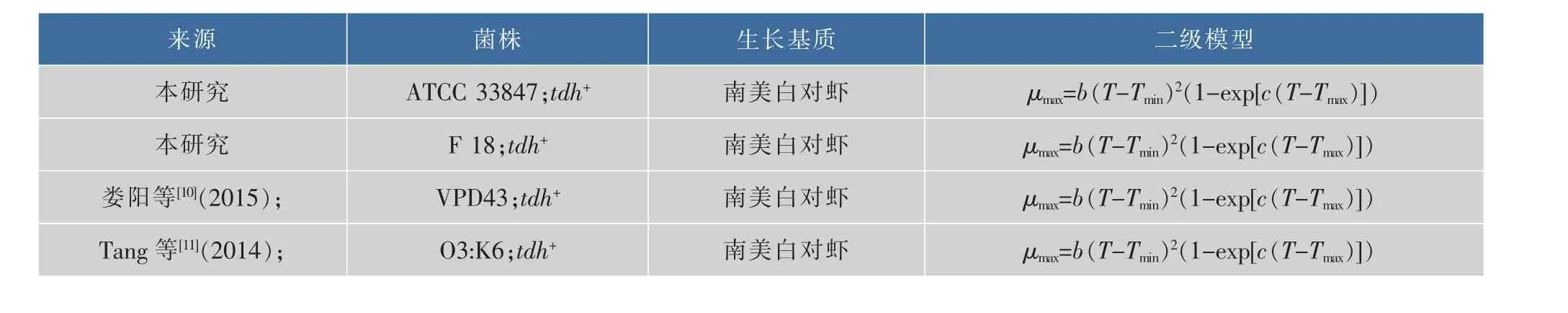
表3 本研究與同類研究副溶血性弧菌的生長差異比較Table 3 Comparison of growth parameters of V.parahaemolyticus in the present study and the similar researches
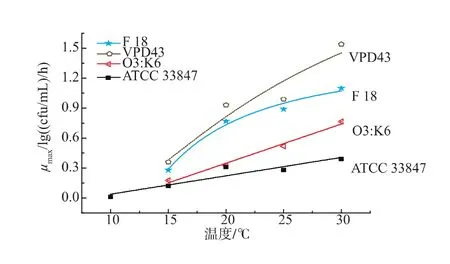
圖3 本研究中致病性副溶血性弧菌在10~30℃的μmax與同類文獻做比較Fig.3 A μmaxcomparison of published and observed V.parahaemolyticus from 10℃to 30℃
3 結 語
預測微生物學模型是食品安全領域中的一個重要研究主題,為了能方便、準確預測食品的貨架期、控制食品的品質以及對食品進行準確的風險預警、風險評估,目前,國內外的學者針對不同條件不同致病微生物和腐敗微生物開展了大量的研究工作,這些工作使得預測微生物學的數據基礎得以逐步健全。如王璐華等[15]采用Gompertz和Linear方程擬合副溶血性弧菌在4~40℃條件下的生長曲線,結果采用Gompertz模型作為VP的最適模型;Ting等[16]研究了單增李斯特菌和背景微生物在甜瓜上的生長情況,結果表明3-parameter logistic model可以較好的擬合單增李斯特菌在背景微生物下的生長情況;賀群等[17]采用修正Gompertz模型、修正Logistic模型和Baranyi模型擬合面包蝦中副溶血性弧菌在10~37℃條件下的生長情況,結果表明Gompertz模型擬合效果最佳。Yoon等采用修正Gompertz模型致病性和非致病性副溶血性弧菌擬合在肉湯和牡蠣勻漿中的生長動力學,并用Davey和平方根模型等二級模型擬合延滯期和最大比生長速率隨溫度的變化曲線,結果表明建立的模型可有效安全地預測副溶血性弧菌的生長。Tang等[10]比較了不同致病性的副溶血性弧菌在南美白對蝦以及培養基上的生長差異,證明不同致病性的副溶血性弧菌的生長動力學參數存在一定的偏差。
由此可見,副溶血性弧菌的預測模型研究更多的集中在致病性、非致病性菌株在不同條件下、不同基質中的生長特性以及預測微生物模型的建立,這些研究為預測食品貨架期、風險評估提供了一定的數據基礎,但是因菌株異質性所帶來的生長差異分析有待于研究完善。從本研究結果可知,4~15℃下,兩株高致病性VP在即食南美白對蝦中的生長特性及生長參數存在顯著性差異(P<0.05);高溫條件下,兩株高致病性VP的生長參數存在差異(P<0.05);在擬合最適模型時,F 18在10℃時的生長數據用所選用的模型無法擬合,而ATCC 33847的生長數據擬合效果良好;建立二級模型時,F 18的最大比生長速率始終大于ATCC 33847,最大差值可達到 0.7 lg((cfu/mL)/h);F 18 的延滯期始終小于ATCC 33847,最大差值可達到7.3 h。而與其他文獻相比較發現,攜帶有相同致病基因的VP的最大比生長速率以及延滯期值等動力學生長參數存在顯著性差異(P<0.05),這證明相同致病性菌株的生長特性存在異質性,而這會對風險評估的準確性帶來一定的影響。
在進行微生物風險評估時,現有的大多數研究所建立的預測模型并未根據菌株異質性對所選用的模型進行修正。而從模型的角度出發考慮,研究越貼近菌株的真實情況,所建立的預測模型更具有參考價值,以不切合現實情況或遠離現實情況為基礎所建立的模型存在很大的不確定性,不利于相關管理部門直接使用風險評估的結果對該食品做出管理措施,也不利于消費者的健康消費。所以,在研究副溶血性弧菌的或者其他水產品時,應將菌株異質性作為影響因素之一,在之后應詳細研究不同來源、不同致病性、不同血清型的副溶血性弧菌的生長、死亡等生長特征,建立不同影響因素下或者混合副溶血性弧菌的預測模型,為風險評估提供更可靠、更全面的數據基礎。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
音樂探索(2022年2期)2022-05-30 21:01:37
小讀者(2021年2期)2021-03-29 05:03:48
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
華人時刊(2019年13期)2019-11-17 14:59:54
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
文苑(2018年22期)2018-11-19 02:54:14
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
光學精密工程(2016年6期)2016-11-07 09:07:19
