谷子FT/TFL1家族的全基因組鑒定與分析
2018-09-12 02:25:30趙慶英楊致榮
山西農(nóng)業(yè)科學(xué) 2018年9期
趙慶英,楊致榮
(山西農(nóng)業(yè)大學(xué)文理學(xué)院,山西 太谷 030801)
谷子(Setaria italica)是傳統(tǒng)的優(yōu)勢(shì)作物,具有耐瘠薄、耐干旱、耐儲(chǔ)藏、營(yíng)養(yǎng)豐富、糧草兼收等特點(diǎn)[1]。針對(duì)日益嚴(yán)重的水資源短缺問(wèn)題,谷子作為重要的戰(zhàn)略?xún)?chǔ)備作物,對(duì)保障干旱、貧瘠地區(qū)的糧食安全起著非常重要的作用[2]。隨著人們消費(fèi)水平的提高以及對(duì)食品營(yíng)養(yǎng)與安全的重視,人們對(duì)小米的需求也逐年增加,但是小米的產(chǎn)量低,還不能滿足現(xiàn)代化產(chǎn)業(yè)的需求。因此,提高谷子產(chǎn)量是谷子研究的主要方向之一。
開(kāi)花是植物從營(yíng)養(yǎng)生長(zhǎng)到生殖生長(zhǎng)轉(zhuǎn)變的開(kāi)始,是植物生長(zhǎng)發(fā)育的重要階段,植物在適宜的條件和時(shí)間開(kāi)花,能夠確保其生物量和繁殖率[3]。植物開(kāi)花受到幾種開(kāi)花途徑的共同調(diào)節(jié),如光周期途徑、春化途徑、內(nèi)源激素和自主途徑等[4],F(xiàn)T和TFL1基因是這些開(kāi)花途徑下游的關(guān)鍵基因,編碼磷脂酰乙醇胺結(jié)合蛋白(PEBP)[5],在植物從營(yíng)養(yǎng)生長(zhǎng)向生殖生長(zhǎng)轉(zhuǎn)變的過(guò)程中起著重要的作用[6]。在擬南芥中,F(xiàn)T/TFL1家族有6個(gè)成員,分別為FLOWERING LOCUS T (FT),TERMINAL FLOWER 1(TFL1),TWIN SISTER OF FT (TSF),BROTHER OF FT AND TFL1 (BFT),ARABIDOPSIS THALIANA CENTRORADIALIS HOMOLOG(ATC)和 MOTHER OF FT AND TFL1(MFT),可以分為 3 個(gè)亞類(lèi),分別為FT亞類(lèi)(包括FT和TSF),MFT亞類(lèi)和TFL1亞類(lèi)(包括TFL1,BFT和ACT基因)[6-7]。
FT是一個(gè)開(kāi)花整合基因,也是誘導(dǎo)開(kāi)花的關(guān)鍵基因,它能促進(jìn)植物從營(yíng)養(yǎng)生長(zhǎng)到生殖生長(zhǎng)的轉(zhuǎn)變[7-8]。在長(zhǎng)日照條件下,F(xiàn)T基因能在長(zhǎng)日照植物(擬南芥等)的葉片維管組織伴胞中被CONSTANS(CO)基因誘導(dǎo)激活表達(dá)[9],F(xiàn)T蛋白與一種新型的內(nèi)質(zhì)網(wǎng)膜蛋白 FT-INTERACTINGPROTEIN1(FTIP1)結(jié)合后被轉(zhuǎn)運(yùn)到莖頂端分生組織,與堿性亮氨酸拉鏈結(jié)構(gòu)域(basic Leucine Zipper,bZIP)轉(zhuǎn)錄因子 FD結(jié)合,直接激活開(kāi)花基因SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1(SOC1)和 APETALA 1(AP1)基因,控制植物的成花轉(zhuǎn)變[10-11]。
在擬南芥中,F(xiàn)T基因過(guò)量表達(dá)導(dǎo)致開(kāi)花時(shí)間提前[12];在水稻中,Hd3a是FT的同源基因,在短日照條件下促進(jìn)開(kāi)花[11];將日中性植物番茄中的FT同源基因SlSP3D轉(zhuǎn)到擬南芥中,能使擬南芥開(kāi)花時(shí)間提前,說(shuō)明SlSP3D是一個(gè)開(kāi)花促進(jìn)基因[11]。轉(zhuǎn)錄因子FLC和光敏色素B1可以抑制FT基因的表達(dá)[13]。
在擬南芥中,F(xiàn)T的同源基因TFL1,參與維持莖尖分生組織的狀態(tài)、延遲其向花器官的轉(zhuǎn)變[14],并與花序的形成有關(guān)[15]。TFL1蛋白抑制FT下游基因LEAFY(LFY)和 AP1的表達(dá)[16],維持花序分生組織狀態(tài),抑制花芽分化[17]。tfl1擬南芥突變體葉片減少、開(kāi)花提前,并且莖尖分生組織轉(zhuǎn)化為頂端花;TFL1過(guò)表達(dá),擬南芥的花期延長(zhǎng),次生花序數(shù)目增加[17]。
ATC和BFT過(guò)表達(dá)導(dǎo)致擬南芥晚花,與TFL1基因過(guò)表達(dá)具有相似的表型,MFT過(guò)表達(dá)導(dǎo)致開(kāi)花時(shí)間略有縮短,但這些基因的功能缺失突變并沒(méi)有顯示出任何明顯的表型[7]。MFT在種子萌發(fā)過(guò)程中也具有一定的作用[18]。TSF與FT在氨基酸水平高度相似,TSF促進(jìn)開(kāi)花,但只有在短日照條件下才有明顯的早花表型[5]。
植物成花轉(zhuǎn)變是一個(gè)復(fù)雜的過(guò)程,依賴(lài)于基因互作遺傳網(wǎng)絡(luò)的精細(xì)調(diào)控,F(xiàn)T/TFL1在該過(guò)程中起著重要的作用。人們對(duì)擬南芥和水稻等模式植物的FT/TFL1研究比較深入,但是在谷子中相關(guān)的研究較少。
本試驗(yàn)通過(guò)對(duì)谷子FT/TFL1基因家族進(jìn)行生物信息學(xué)分析,初步探究FT/TFL1基因家族與成花轉(zhuǎn)變的關(guān)系,旨在為深入研究谷子抽穗開(kāi)花機(jī)制提供理論依據(jù)。
1 材料和方法
1.1 篩選擬南芥、水稻和谷子FT/TFL1家族成員
在擬南芥中,有6個(gè)FT/TFL1家族成員已被報(bào)道,利用 NCBI(https://www.ncbi.nlm.nih.gov/)和Phytozome(https://phytozome.jgi.doe.gov/)網(wǎng)站在線搜索擬南芥FT/TFL1家族成員蛋白序列并下載,通過(guò)BLAST比對(duì),得到水稻和谷子FT/TFL1家族成員的蛋白序列和啟動(dòng)子序列。
1.2 構(gòu)建FT/TFL1家族系統(tǒng)發(fā)育樹(shù)
通過(guò)MEGA7[19]軟件對(duì)谷子、擬南芥以及水稻FT/TFL1家族成員進(jìn)行蛋白序列比對(duì),利用Neighbor-joining算法分析構(gòu)建系統(tǒng)發(fā)育樹(shù),選擇Bootstrap方法,其他參數(shù)選擇默認(rèn)值。
1.3 FT/TFL1家族保守序列分析
利用MEME v4.11.0(http://meme-suite.org/tools/meme)網(wǎng)站在線分析擬南芥、水稻和谷子的保守序列。保守位點(diǎn)的寬度設(shè)置為≥6和≤50,最大保守序列鑒定數(shù)目設(shè)定為10。
1.4 FT/TFL1家族啟動(dòng)子元件分析
利用 PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)在線分析啟動(dòng)子元件。
1.5 FT/TFL1家族表達(dá)模式分析
利用R[20]軟件的heatmap函數(shù)繪制基因表達(dá)熱譜圖。
2 結(jié)果與分析
2.1 FT/TFL1家族系統(tǒng)發(fā)育樹(shù)分析
擬南芥FT/TFL1基因家族中已知的6個(gè)成員FT,TSF,MFT,TFL1,BFT 和 ATC 根據(jù)其結(jié)構(gòu)和功能分為3個(gè)亞類(lèi),分別為FT亞類(lèi)(包括FT和TSF),MFT亞類(lèi)和 TFL1亞類(lèi)(包括 TFL1,BFT和ACT 基因)[5,7,21]。
為了分析谷子FT/TFL1家族成員的系統(tǒng)進(jìn)化關(guān)系,利用Mega 7.0軟件構(gòu)建了擬南芥、水稻和谷子的系統(tǒng)進(jìn)化樹(shù)。從圖1可以看出,谷子FT/TFL1家族可以分為3個(gè)亞類(lèi),即FT,MFT和TFL1亞類(lèi),F(xiàn)T和MFT亞類(lèi)對(duì)開(kāi)花起促進(jìn)作用,TFL1亞類(lèi)抑制開(kāi)花,同一亞類(lèi)的基因具有相同或相似的功能,在植物開(kāi)花過(guò)程中共同作用,調(diào)控植物開(kāi)花。

2.2 FT/TFL1家族保守序列分析
利用MEME在線軟件對(duì)擬南芥、水稻和谷子的FT,MFT和TFL1亞類(lèi)的蛋白保守位點(diǎn)進(jìn)行了分析,結(jié)果如圖2所示,3個(gè)亞家族成員均有一段含有50個(gè)氨基酸殘基的保守序列。FT亞家族的保守基序?yàn)閄2DPDXPXPSXPX3EXLHWXVTDIPX7GXEX3YE XPXPX2GXH,MFT亞家族的保守基序?yàn)镸TDPDAP SPSXPXMREX2HWXVX2IPGGTX4GX5YMXPRPXVG IH,TFL1亞家族的保守基序?yàn)镋X2GXDXRSFFTLX MXDPDXPXPSX5EXLHWXVTXIPGTTDX2FGXE。FT和TFL均有一個(gè)決定其功能的關(guān)鍵氨基酸殘基,F(xiàn)T蛋白的第85位是一個(gè)酪氨酸殘基,對(duì)控制開(kāi)花時(shí)間具有重要的作用[22],TFL1蛋白第88位的組氨酸對(duì)于維持頂端分生組織的生長(zhǎng)和延遲開(kāi)花起主要作用,MFT蛋白的色氨酸也能促進(jìn)擬南芥開(kāi)花,但是相對(duì)于組氨酸和酪氨酸作用較弱,這幾個(gè)主要的氨基酸殘基與其他殘基共同作用,調(diào)控?cái)M南芥成花轉(zhuǎn)變[23]。
2.3 FT/TFL1家族啟動(dòng)子序列分析
利用PlantCARE軟件在線分析谷子FT/TFL1基因啟動(dòng)子區(qū)順式作用元件,結(jié)果發(fā)現(xiàn),除了啟動(dòng)子和增強(qiáng)子區(qū)常見(jiàn)的順式作用元件CAAT-box和核心啟動(dòng)子元件TATA-box外,還具有光響應(yīng)元件sp1和G-box,胚乳表達(dá)順式作用元件Skn-1_motif以及與逆境脅迫相關(guān)的順式作用元件,如脫落酸響應(yīng)元件ABRE、低溫響應(yīng)元件LTR和厭氧誘導(dǎo)響應(yīng)元件ARE等(表1)。說(shuō)明FT/TFL1基因家族成員在胚乳表達(dá)、響應(yīng)光信號(hào)和抵抗逆境過(guò)程中起重要作用。

表1 谷子FT/TFL1基因家族啟動(dòng)子順式元件預(yù)測(cè)
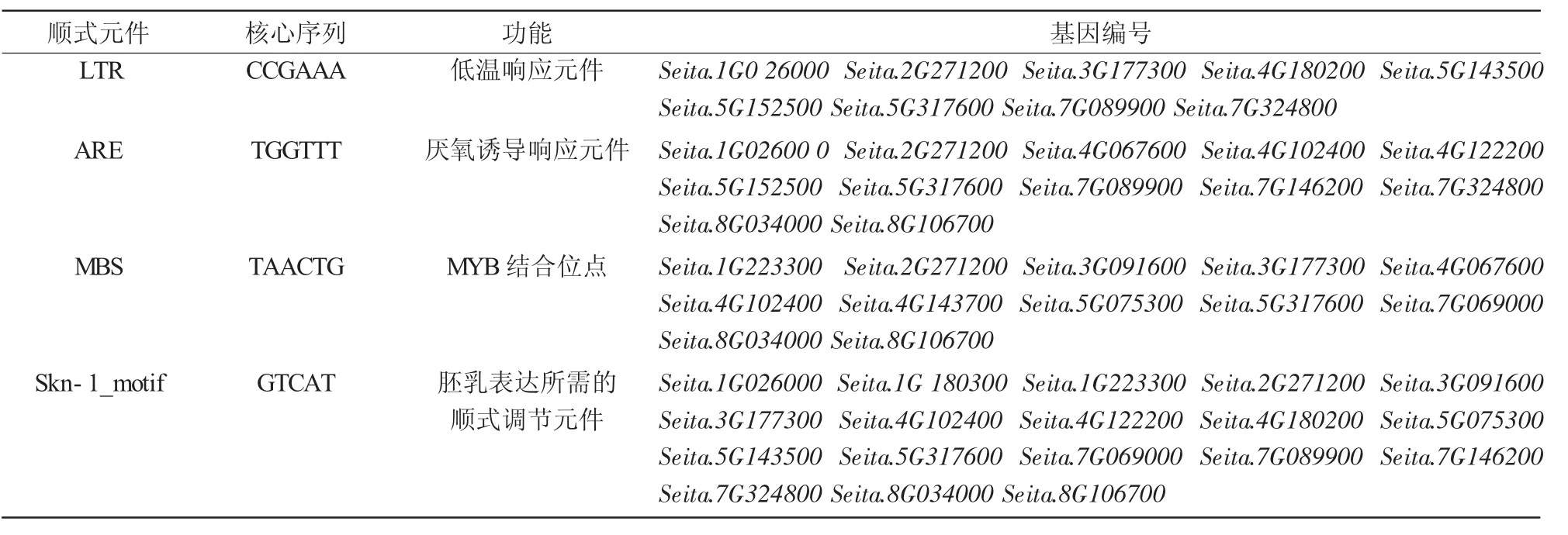
續(xù)表1
2.4 FT/TFL1家族表達(dá)模式分析
通過(guò)NCBI和Phytozome網(wǎng)站檢索到29個(gè)谷子FT/TFL1基因家族成員,通過(guò)Phytozome網(wǎng)站,統(tǒng)計(jì)了FT/TFL1基因家族成員在各個(gè)組織中的表達(dá)量,通過(guò)R語(yǔ)言中的heatmap函數(shù)繪制熱圖,結(jié)果發(fā)現(xiàn),Seita.4G180200基因在穗中的表達(dá)量最高,Seita.4G122200基因在穗中的表達(dá)量也較高,Seita.4G122200基因在根中的表達(dá)量最高;TFL1亞家族的Seita.1G180300和Seita.7G324800基因在根中的表達(dá)量較高,Seita.4G122200的在葉中和苗中的表達(dá)量也較高。FT/TFL1家族不同成員在谷子不同組織中的表達(dá)量存在顯著差異(圖3)。

3 結(jié)論與討論
植物FT/TFL1基因是植物由營(yíng)養(yǎng)生長(zhǎng)向生殖生長(zhǎng)轉(zhuǎn)變的關(guān)鍵基因,通過(guò)對(duì)FT/TFL1基因家族進(jìn)行生物信息學(xué)分析,為深入研究谷子FT/TFL1基因家族對(duì)成花轉(zhuǎn)變的調(diào)控奠定了基礎(chǔ)。
通過(guò)生物信息學(xué)分析發(fā)現(xiàn),谷子FT/TFL1家族可以分為3個(gè)亞類(lèi),分別為FT亞類(lèi),MFT亞類(lèi)和TFL1亞類(lèi),其中,F(xiàn)T亞類(lèi)和MFT亞類(lèi)對(duì)開(kāi)花起促進(jìn)作用,TFL1亞類(lèi)抑制開(kāi)花,3個(gè)亞類(lèi)的基因相互協(xié)調(diào),共同調(diào)控谷子開(kāi)花。
谷子、水稻和擬南芥的FT,MFT和TFL1亞類(lèi)的蛋白序列高度保守,均具有一段含有50個(gè)氨基酸殘基的保守序列,F(xiàn)T蛋白第85位的酪氨酸殘基、TFL1蛋白第88位的組氨酸殘基分別對(duì)調(diào)控?cái)M南芥開(kāi)花和維持頂端分生組織的生長(zhǎng)具有重要作用。
通過(guò)對(duì)谷子FT/TFL1基因家族起始密碼子上游約1 500 bp左右的啟動(dòng)子順式作用元件分析發(fā)現(xiàn),它主要有光響應(yīng)元件sp1、胚乳表達(dá)順式作用元件Skn-1_motif以及與逆境相關(guān)的元件,如低溫響應(yīng)元件LTR等。說(shuō)明FT/TFL1基因家族成員在胚乳表達(dá)、響應(yīng)光信號(hào)和抵抗逆境過(guò)程中起重要作用。
通過(guò)對(duì)谷子表達(dá)模式分析發(fā)現(xiàn),Seita.4G122200基因在根、葉和苗中的表達(dá)量都最高,Seita.4G180200基因在穗中的表達(dá)量最高,F(xiàn)T/TFL1家族在不同組織中的表達(dá)量存在顯著差異。
綜上所述,本研究通過(guò)對(duì)谷子FT/TFL1家族的生物信息學(xué)分析,初步探究了谷子FT/TFL1家族在谷子開(kāi)花途徑中的作用,結(jié)果為以后研究FT/TFL1家族成員控制谷子開(kāi)花的具體調(diào)控機(jī)制提供了理論依據(jù)。
