擬南芥表皮毛突變體abt2基因克隆及功能研究
2018-09-10 07:00:46安麗君
江蘇農業(yè)科學 2018年15期
王 巖, 張 翔, 安麗君
(旱區(qū)逆境生物學國家重點實驗室/西北農林科技大學生命科學學院,陜西楊凌 712100)
表皮毛是植物地上部組織表皮向外突出形成的單細胞或多細胞結構[1-2]。表皮毛作為植物與環(huán)境接觸的最外層屏障,具有多種生理、生態(tài)功能。表皮毛的存在能抵御植食性昆蟲和病原菌對植物體的危害[3];也可以減少植株熱量的散失,提高植物對冷害的抗性[4-5];同時也能減少植物對太陽輻射的吸收,保護組織免受紫外線和極端溫度的傷害[6];增強葉片對空氣中水蒸氣的儲藏能力,提高植物的抗旱性等[7],以及吸收重金屬元素,提高植物對土壤重金屬污染的防御[8-9]。另外,表皮毛作為一種特化的表皮細胞,也是研究細胞命運決定、細胞周期調控、細胞極性生長以及細胞成熟的優(yōu)良模式系統(tǒng)[10]。
在模式植物擬南芥中,表皮毛存在于除下胚軸和子葉外的幾乎所有氣生器官上,如蓮座葉、主莖、莖生葉以及花瓣等[11]。擬南芥蓮座葉上的表皮毛為典型的單細胞結構,形態(tài)上通常為3~4個分支,其發(fā)育過程受到多種因子的調控[12]。其中KAK基因編碼1個HECT泛素蛋白連接酶,抑制表皮毛細胞從有絲分裂向核內復制過程的轉變,KAK基因功能缺失導致細胞核DNA含量升高,表皮毛分支數(shù)增加[13]。
本試驗在以擬南芥表皮毛突變體gl2-3為背景的EMS二次誘變庫中,篩選獲得了1個蓮座葉表皮毛分支數(shù)明顯增加的突變體abt2。通過回交并觀察后代的表皮毛表型,發(fā)現(xiàn)abt2是細胞核單基因控制的隱性突變體;圖位克隆以及雜交試驗證實突變基因abt2是KAK基因的一個新等位基因,abt2的突變表型是由于KAK基因第+6 881 bp處核苷酸發(fā)生突變所導致的。
1 材料與方法
1.1 試驗材料
擬南芥(Arabidopsisthaliana)野生型Columbia(Col-0)、Landsbergerecta(Ler)、gl2-3(SALK_039825)突變體純合體、KAK基因T-DNA插入功能缺失突變體SALK_037636由筆者所在實驗室保存。gl2-3背景的EMS突變體庫由筆者所在實驗室創(chuàng)制保存。
1.2 試驗方法
1.2.1 植物的培養(yǎng)與候選突變體篩選 將gl2-3背景的EMS誘變體庫按庫種植在充分吸水后的進口品氏基質泥炭土(pindstrup substrate peat moss)上,4 ℃黑暗處理3 d后移入植物培養(yǎng)間,培養(yǎng)溫度為(22±2)℃,相對濕度為70%,24 h 持續(xù)光照,光照度為100 μmol/(m2·s)。待植株生長2周后觀察蓮座葉葉表皮毛發(fā)育狀況,篩選表皮毛發(fā)育異常的植株作為候選突變體。
1.2.2 突變體遺傳背景純化及遺傳分析 先將候選突變體進行自交并觀察后代表型,選取突變表型明顯且能穩(wěn)定遺傳的候選突變體作為目標突變體;再以擬南芥Col-0為母本,目標突變體為父本進行回交,觀察F2代群體中出現(xiàn)的表型并統(tǒng)計分離比,判斷突變基因的顯隱性以及控制突變表型的基因數(shù)目;另外,通過正反交試驗判斷突變表型的遺傳方式。
1.2.3 圖位克隆 將確定的目標突變體與擬南芥Ler生態(tài)型進行雜交,在F2代植株中挑選具有突變體表型的植株作為圖位克隆群體。首先提取95株作圖群體中植株的基因組DNA,構建DNA池(bulk),利用已經公布的均勻覆蓋擬南芥5條染色體的27對分子標記為引物進行PCR,通過統(tǒng)計突變位點與分子標記的連鎖情況,判斷突變位點所在的染色體區(qū)間;確定突變位點所在區(qū)間及與其連鎖的側翼分子標記后,擴大作圖群體,設計新分子標記進行精細定位。
1.2.4 候選基因的確定 根據(jù)精細定位的區(qū)間,在擬南芥基因組數(shù)據(jù)庫(www.arabidopsis.org)中下載該區(qū)間內所覆蓋的全部基因,查閱相關資料,篩選出候選突變基因;然后針對這些候選突變基因設計引物進行PCR擴增并測序,最終確定出突變基因。
1.2.5abt2與SALK_037636突變體之間的遺傳互作分析 將abt2與SALK_037636雜交,觀察F1代植株的表型,驗證KAK基因與abt2基因之間的等位性。
1.2.6 表皮毛分支表型分析 將Col-0、abt2、SALK_037636及abt2×SALK_037636 F1代植株培養(yǎng)3周后,觀察并統(tǒng)計第3、第4張蓮座葉上具有不同分支數(shù)的表皮毛數(shù)量,并計算各種類型表皮毛所占的比例。每種基因型植株至少統(tǒng)計20株,利用GraphPad Prism 6軟件進行作圖。
2 結果與分析
2.1 abt2突變體的獲得和表型分析
為獲得參與調控擬南芥表皮毛發(fā)育的新基因,構建了以植物表皮毛發(fā)育的關鍵調控因子GL2功能缺失突變體gl2-3為背景的EMS誘變庫,并進行了大規(guī)模的遺傳篩選。在此過程中獲得了1個蓮座葉表皮毛分支數(shù)明顯增加的突變體,將其命名為abt2(aberrant trichome 2)。通過統(tǒng)計第3、第4張蓮座葉上不同類型的表皮毛數(shù)量,發(fā)現(xiàn)與Col-0相比,abt2突變體中表皮毛分支數(shù)顯著增加。在Col-0中,表皮毛主要以3個分支為主,約占表皮毛總數(shù)量的77.83%(第3張蓮座葉)、75.69%(第4張蓮座葉),僅存在極少數(shù)5個分支的表皮毛;而在abt2突變體中,表皮毛主要以4、5個分支為主,4個分支的表皮毛約占葉片表皮毛總數(shù)量的60.55%(第3張蓮座葉)、61.19%(第4張蓮座葉);5個分支表皮毛約占葉片表皮毛總數(shù)量的30.69%(第3張蓮座葉)、30.14%(第4張蓮座葉),并且能觀察到一定數(shù)量的6個分支的表皮毛(第3張蓮座葉上為 2.42%,第4張蓮座葉上為1.66%)(圖1)。

2.2 abt2突變基因的定位
2.2.1abt2突變位點的遺傳分析 由于abt2突變體表皮毛發(fā)育缺陷表型非常明顯,所以期望通過圖位克隆分離突變基因,以進一步研究其調控表皮毛發(fā)育的機制。在進行圖位克隆之前,首先對abt2突變位點進行遺傳分析。通過將Col-0與abt2突變體進行正反交并觀察表皮毛表型,發(fā)現(xiàn)所獲得的F1代植株均表現(xiàn)野生型表皮毛表型,表明abt2突變體是細胞核基因控制的隱性突變體;F2代植株中野生型表型與突變體表型個體的分離比約為3 ∶1,符合孟德爾隱性單基因遺傳定律,從而說明abt2突變體的發(fā)育表型是由單基因控制的。
2.2.2abt2突變位點的粗定位 在確定了abt2突變體的遺傳特性之后,對abt2的突變位點進行粗定位。首先將Ler生態(tài)型植株與abt2突變體進行雜交,在F2代中挑選具有abt2表型的植株作為作圖群體,利用實驗室已有的均勻分布在擬南芥5條染色體的27個分子標記(圖2)和由95株作圖群體植株的基因組DNA構成的DNA池進行PCR擴增,根據(jù)與突變位點連鎖的分子標記所擴增出條帶的遺傳背景偏向突變體來源的Col-0,而用于雜交的另一親本Ler條帶不能擴增或擴增量弱于對照的原理,發(fā)現(xiàn)突變位點與第4條染色體上分子標記F20D10#1(17933942)連鎖較為緊密,說明突變位點可能位于該分子標記附近(圖2)。
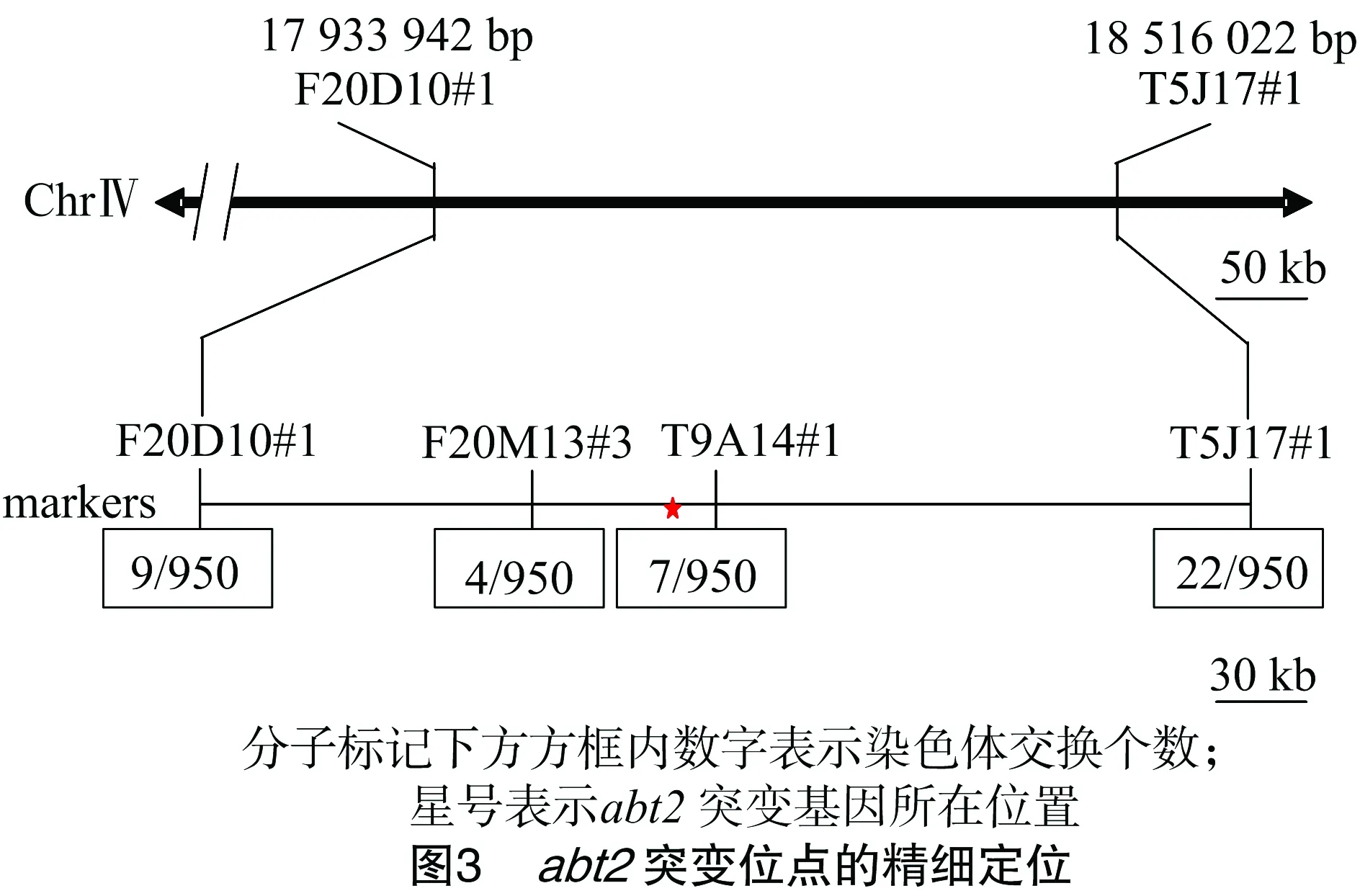
2.2.3 突變位點的精細定位 確定突變位點與染色體的連鎖關系后,為了進一步確定突變位點在染色體上所在的區(qū)間,以95個突變體DNA為模板,以F20D10#1(17933942)及在F20D10#1下方增加的新的分子標記T5J17#1(18516022)為引物進行PCR擴增,結果(表1)顯示突變位點位于分子標記F20D10#1和T5J17#1之間,所以將分子標記F20D10#1和T5J17#1作為后續(xù)定位的側翼分子標記(flanking marker)(圖3)。接著通過在Flanking marker區(qū)間內增加新的分子標記對abt2突變位點進行精細定位。利用1個新的dCAPS引物F20M13#3(18010126)和1個新的插入/刪除(In/Del)分子標記T9A14#1(18053432)(表1)對F2作圖群體中475株突變體表型植株的基因型進行分析,最終將突變基因定位在分子標記F20M13#3和T9A14#1的區(qū)間內,該區(qū)間的物理距離為43 306 bp(圖3)。


表1 精細定位所用分子標記
2.2.4 候選基因的篩選與確定 確定了abt2突變位點所在的區(qū)間后,利用擬南芥基因組數(shù)據(jù)庫對該區(qū)間內所覆蓋的全部基因進行分析,重點關注與表皮毛發(fā)育相關的基因。通過查閱文獻發(fā)現(xiàn),在該區(qū)間內基因At4g38600被報道與表皮毛發(fā)育有關。At4g38600基因也被稱為KAKTUS/UbiquitinProteinLigase3(KAK/UPL3),編碼E3泛素蛋白連接酶,KAK基因的功能缺失會導致擬南芥蓮座葉上的表皮毛分支數(shù)顯著增加,與abt2突變體的表皮毛發(fā)育缺陷表型相似[14-15]。因此推測abt2突變體的表型很可能是由于KAK基因的功能缺失所導致的。為了驗證abt2是否為KAK的新等位基因,首先對abt2突變體背景中的KAK基因進行測序,發(fā)現(xiàn)在abt2突變體中KAK基因的第6 881 bp處發(fā)生了單堿基突變,由鳥嘌呤(G)突變?yōu)橄汆堰?A)(圖4), 這表明abt2 突變表型的產生很可能是由KAK基因的單堿基突變造成的,KAK可能就是abt2的突變基因。

2.2.5abt2和KAK基因間的遺傳互作分析 為了進一步確定KAK基因的突變是造成abt2突變表型產生的原因,利用KAK基因的功能缺失突變體SALK_037636對abt2和KAK基因之間的等位性進行分析。SALK_037636的突變表型是由于在KAK基因的第+1 241 bp處插入了T-DNA,導致KAK的表達量降低所造成的(圖5)。
將abt2與SALK_037636進行雜交并獲得了F1代植株,通過觀察F1代植株第3、第4張蓮座葉上的表皮毛發(fā)育表型,發(fā)現(xiàn)其與abt2和SALK_037636的表型相似,表皮毛分支數(shù)明顯增加(圖6),這表明abt2基因突變與KAK基因突變不能互補彼此的突變表型,abt2的突變基因就是KAK,abt2是KAK的新等位基因。
3 討論與結論
特色遺傳材料的獲得是研究基因功能的強有力手段。為了獲得新的參與調控植物表皮毛發(fā)育的新因子,本試驗利用EMS高效誘變擬南芥gl2-3所建立的二次突變體庫進行了大規(guī)模的遺傳篩選。在此過程中獲得了1個蓮座葉上表皮毛分支數(shù)明顯增加的突變體,并將其命名為abt2。通過圖位克隆及遺傳互作分析,表明abt2的突變表型是由KAK基因的+6 881 bp處發(fā)生由鳥嘌呤到腺嘌呤的單堿基突變所造成的,abt2基因是KAK基因的一個新等位基因。同時也發(fā)現(xiàn),盡管在abt2突變體背景中KAK基因發(fā)生了單核苷酸突變,但是該突變并未造成所編碼的相關氨基酸發(fā)生改變,因此為無義突變。該無義突變如何造成了abt2突變體表現(xiàn)出了顯著的表皮毛發(fā)育缺陷表型將是下一步研究的重點。另外,KAK是表皮毛核內復制的抑制基因,KAK功能缺失突變體的蓮座葉表皮毛呈現(xiàn)過度分支的表型,并伴隨表皮毛細胞核DNA含量增加[16],本試驗所獲得的abt2突變體為進一步研究KAK基因與表皮毛調控之間的關系提供了新的遺傳材料,也為深入研究KAK參與調控植物表皮毛發(fā)育的可能機制奠定了基礎。


