發芽糙米γ-氨基丁酸形成的谷氨酸脫羧酶活性與底物變化的相關性分析
2018-08-31 02:32:12曹晶晶顧豐穎羅其琪劉子毅
食品科學 2018年16期
曹晶晶,顧豐穎,羅其琪,劉子毅,王 鋒*
γ-氨基丁酸(γ-aminobutyric acid,GABA)是一種非蛋白質氨基酸,是動物及人體中樞神經系統中重要抑制性傳遞物質[1]。GABA通過調節中樞神經系統,具有降血壓的作用,其舒緩血管和降血壓的藥理功效已在大量的動物實驗和臨床醫學中得以證實[2-3],同時具有鎮靜神經、抑制抑郁的功效[4]。植物性來源的GABA產品因其安全性、天然性的特點備受關注,目前富含GABA食品有發芽米、發芽豆、谷物早餐產品等[5]。
在植物體內GABA主要通過谷氨酸脫羧酶(glutamic acid decarboxylase,GAD)催化谷氨酸(glutamic acid,GA)脫羧生成,其中GAD催化是其產生的主要限制性途徑[6]。GAD是一種磷酸吡哆醛(pyridoxal phosphate,PLP)-關聯酶,其活性被PLP提升[7]。GAD是一種Ca2+/鈣調蛋白依賴型的酶,具有一個鈣調蛋白結合區[8],外源添加Ca2+可提高GAD活性,促進GA轉化為GABA[9]。
發芽糙米是富含GABA植物性食品之一,也是開發GABA功能食品主要原料。發芽處理是一種有效改善谷物食用品質和營養價值的方法,可大大提高產品的生物利用率。在浸泡發芽過程中,內源性酶被生產或激活,分解淀粉及蛋白質等主要大分子,產生一些次生代謝物,如GABA和酚醛物質[10]。浸泡和發芽處理能夠顯著增加糙米中GABA含量[11],發芽過程中的環境壓力對GABA含量增加有顯著促進作用,如厭氧脅迫、低壓等離子體照射、熱激處理等[12-14]。逆境對細胞內部產生破壞,降低細胞質pH值,從而有利于GA脫羧而不利于GABA轉氨,從而顯著增加植物體內GABA含量[15]。超聲波處理是一種常用的逆境脅迫處理方式,通過一定頻率的機械振動,適當強度的超聲波作用于糙米,具有提高酶活、促進細胞生長和生物合成的作用,可促進部分營養成分含量增加[16-17]。
目前關于發芽糙米中GABA的研究主要集中在發芽條件的優化,對于GABA形成的酶及底物之間的關系研究較少。因此,本實驗主要研究糙米發芽不同階段的GA含量、GAD活性及GABA含量變化,同時考察超聲波對發芽糙米GABA含量的影響,探究發芽糙米中GA含量、GAD活性與GABA含量三者之間的關系,為提高發芽糙米的生理功效提供參考。
1 材料與方法
1.1 材料與試劑
糙米品種為五優稻4號,由黑龍江北純農產品開發有限公司提供,粳型常規水稻。2016年10月收獲,室溫密封保存。
GABA標準品、PLP 美國Sigma-Aldrich公司;β-巰基乙醇 美國Amresco公司;鹽酸(優級純),無水乙醇、乙二胺四乙酸、GA(均為分析純) 國藥集團化學試劑有限公司。
1.2 儀器與設備
DYJ-Y01糙米發芽機 中山市長益家用電器有限公司;FD-1C-80冷凍干燥機 上海比朗儀器制造有限公司;KQ-600DE超聲波清洗器 昆山市超聲儀器有限公司;TTL-DCII型氮吹儀 北京同泰聯科技發展有限公司;L-8900氨基酸分析儀 日立高新技術公司。
1.3 方法
1.3.1 糙米浸泡預處理
在肖君榮等[18]的方法上作適當改動,糙米用1%次氯酸鈉浸泡10 min消毒,用去離子水沖洗3 次瀝干。糙米在發芽前進行浸泡3 h預處理。預處理條件為30 ℃營養液(2.5 mmol/L Ca2+濃度,pH 5.5)浸泡3 h。
1.3.2 制備糙米發芽
糙米預處理結束后,采用糙米發芽機30 ℃至發芽結束,每隔12 h更換一次營養液。在發芽至不同時間點進行超聲波處理。超聲波處理條件:參考張祎等[19]的方法,適當改動,將糙米置于注有營養液的器皿中,在30 ℃、40 kHz、240 W條件下超聲波處理15 min。超聲波處理不計入總發芽時長。具體處理條件如表1所示。

表1 超聲波處理方式Table 1 Conditions for ultrasonic treatment
在發芽過程中第0、12、24、36、48、60、72小時取出部分樣品,用蒸餾水沖洗3 次瀝干,冷凍干燥至水分質量分數小于10%。磨粉,過80 目篩,4 ℃貯藏待測。
1.3.3 指標測定
1.3.3.1 GABA及游離氨基酸含量測定
在Chen Huahan[14]和Komatsuzaki[20]等方法基礎上作適當調整。稱取1.0 g發芽糙米粉,加入5 mL去離子水,超聲提取30 min。8 000 r/min離心,取上清液,重復提取1 次,合并上清液。取3 mL上清液,加入7 mL無水乙醇于4 ℃過夜沉淀,離心收集上清液,氮吹至干。加入1 mL 0.02 mol/L鹽酸溶液溶解干燥物,10 000 r/min離心10 min,上清液即為GABA粗提液。過0.22 μm濾膜,采用氨基酸自動分析儀測定GABA和16 種游離氨基酸(除色氨酸和半胱氨酸)含量。其中GABA質量濃度C(mg/mL)與峰面積A之間的回歸方程為A=5.84×108C+127 176(R2=0.999 8),具有良好的相關性。
1.3.3.2 GAD活性測定
在白青云[21]和Khwanchai[22]等測定方法基礎上作適當調整。取2.00 g發芽糙米,加入10 mL的磷酸鉀緩沖液(0.05 mol/L,pH 7.2),內含2 mmol/L β-巰基乙醇、2 mmol/L乙二胺四乙酸、0.2 mmol/L PLP,4 ℃、250 r/min提取12 h。10 000 r/min冷凍離心20 min,上清液為粗酶提取液。取0.5 mL粗酶液,加入200 μL底物溶液(50 mmol/L GA)和1.8 mL磷酸鉀緩沖液(0.05 mol/L,pH 5.8),內含2 mmol/L β-巰基乙醇、2 mmol/L乙二胺四乙酸、0.2 mmol/L PLP。于37 ℃反應1 h后90 ℃水浴10 min反應滅酶,離心,取1 mL反應液氮吹干燥。加入5 mL 0.02 mol/L鹽酸溶液溶解干燥物,按照1.3.3.1節方法測定GABA含量。GAD活性定義:在37 ℃每克干質量樣品每分鐘產生的GABA量(nmol)表示。
1.4 數據處理
采用Origin 8.0軟件繪圖,SPSS 16.0軟件進行單因素方差分析(ANOVA),P值小于0.05則認為組間差異顯著。
2 結果與分析
2.1 發芽糙米GA含量測定結果
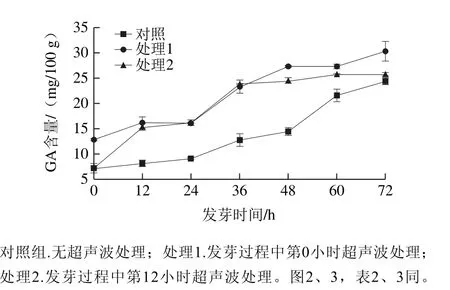
圖1 超聲波處理糙米發芽過程中GA含量變化Fig. 1 Change in GA content during brown rice germination under ultrasonic treatment
浸泡預處理后,對照組糙米中GA含量為7.20 mg/100 g。由圖1可知,隨發芽時間的延長,發芽糙米中GA含量持續增加,主要來源于糙米發芽代謝促使蛋白質水解[20]。超聲波處理組發芽糙米中GA含量明顯高于對照組。其中處理1在發芽0 h時GA含量是對照組的1.78 倍,處理2在發芽12 h時GA含量是對照組的1.88 倍,且處理組在12~24 h之間GA含量較對照組高2 倍左右,表明超聲波促進酶激活,加快了蛋白質降解速率[23],促使GA含量增加;在36 h之后,處理1較處理2發芽糙米中GA含量稍高。在發芽60 h后,GA含量增幅減緩,在后萌發階段糙米呼吸強度增大,干物質消耗量加劇,GA消耗量增加[24]。
GA含量隨著發芽時間的延長持續增加,而Xu Jianguo等[25]發現在裸燕麥富集GABA前期,GA含量隨著發芽時間的延長持續減少。分析認為,游離性GA主要來源于蛋白質分解[7]、谷氨酰胺合成酶循環、GABA轉氨酶反應[6]3 種途徑,GA的轉化途徑主要是合成GABA、合成其他氨基酸或氧化分解。因此,產生游離性GA的速率要大于GA轉化的速率。
2.2 發芽糙米中GAD活性測定結果
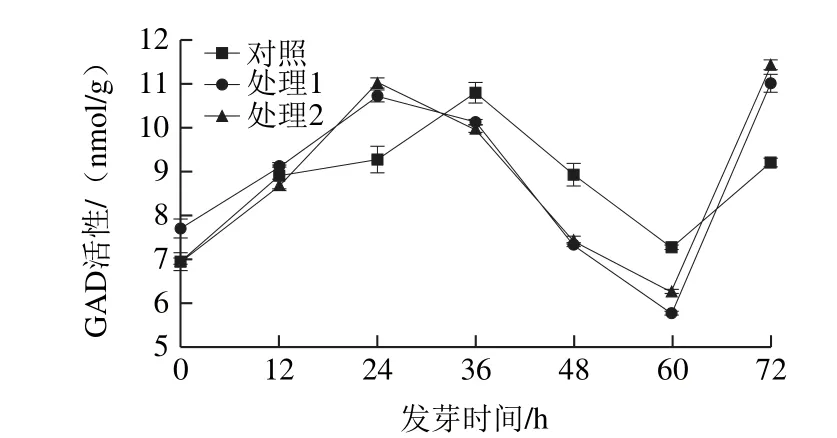
圖2 超聲波處理糙米發芽過程中GAD活性變化Fig. 2 Change in GAD activity during brown rice germination under ultrasonic treatment
糙米經浸泡處理后,對照組GAD活性為7.09 nmol/g。在發芽0~48 h之間,對照組GAD活性均值為9.25 nmol/g。糙米歷經72 h的發芽,GAD活性呈現先增后降再增趨勢。對照組在發芽第36小時達到第1個峰值11.20 nmol/g,在60 h達到谷值7.43 nmol/g,GAD活性降低34%。Xu Jianguo等[25]研究發現GAD活性有類似變化,變化量受燕麥品種影響。韓超等[26]發現米胚芽中GAD活性存在峰值,GA濃度為50~100 mmol/L時達到最大值。處理1在發芽初始GAD活性是對照組的1.11 倍,說明在發芽初期即在水分吸收階段(吸脹作用),表明超聲波處理更有助于GAD激活[16];在發芽第24小時達到第1個峰值11.13 nmol/g,在第60小時GAD活性降低至最低值5.83 nmol/g。處理2與處理1類似,GAD活性值在發芽第24小時達到峰值,在發芽第60小時達到谷值。超聲波處理將GAD活性第1個峰值提前了12 h。超聲組在60 h后,GAD活性增加幅度明顯增大,且高于第1個峰值。溫煥斌[24]發現糙米發芽至60~84 h蛋白酶活性明顯提高。糙米在發芽72 h左右已結束萌發階段,并開始進入幼苗形態建成過程[27-28]。處理1、2之間在糙米發芽過程中對GAD變化無明顯差異,但2 種超聲波處理都使GAD激活進程提前,縮短發芽時間。
Scott-Taggart等[29]的研究表明,GAD活性受GA調節。Dennison等[30]發現南芥體中GA除可自身激活GAD外,還可通過改變細胞質中Ca2+濃度而影響GAD活性。結合圖1可知,在24~36 h間GA增加量較大,GAD活性較高,在48~60 h間處理組GA含量基本保持不變,GAD活性降低。表明發芽糙米中GAD活性受GA含量調節。
2.3 發芽糙米中GABA含量測定結果

圖3 超聲波處理糙米發芽過程中GABA含量變化Fig. 3 Change in GABA content during brown rice germination under ultrasonic treatment
糙米浸泡預處理后,對照組GABA含量為6.84 mg/100 g。由圖3可知,隨發芽時間延長,發芽糙米中GABA含量持續增加,在發芽第72小時,達到68.75 mg/100 g,且在發芽第36小時后,增長速度明顯加快。結合圖1,發芽糙米中GA含量在12~24 h增長幅度緩慢,產物GABA含量在24~36 h時增幅減緩,GABA增加量受GA含量調節,時間稍有延后。結合圖2,在0~36 h間,GAD活性升高到1.60 倍,GABA含量增加到3.56 倍;在36~60 h間GAD活性下降明顯,GABA增加到1.69 倍,GABA增加量受GAD活性調節。
GABA含量在超聲波處理后較對照組低,GABA是游離氨基酸,超聲波處理加速游離氨基酸溶出[31]。與對照組相比,發芽糙米24~48 h之間GABA含量明顯增加,超聲波能夠促進GAD激活[32],進而促進GABA含量的增加。但在48~60 h GABA無明顯增加,GABA是三羧酸循環支路的重要中間產物[7]。植物體中GABA由GAD催化GA脫羧得到[6]。在GABA支路中,GABA通過轉氨基作用轉化為琥珀酸半醛,并進一步轉化成琥珀酸,從而進入三羧酸循環[33]。結合圖2,GAD活性在48~60 h階段較低,直接限制了GABA含量增加,使GABA增加量不明顯。
在糙米發芽后期GABA含量增長幅度明顯增大,因超聲波加快了糙米發芽進程,此時糙米提前結束萌芽,進入幼苗形態建成過程,游離氨基酸等小分子物質增加[27-28]。Zhang Yungang等[34]發現超聲波能量作用于酶分子,可使酶分子的構象發生改變,進而改變酶的生物活性。同時超聲波促進植物細胞壁周圍物質降解,細胞內Ca2+和H+濃度增加,由于GAD是鈣調素蛋白,可以有效激活GAD活性,促進GABA在發芽過程中的積累。超聲波處理在發芽第36小時之前較對照組能增加GABA含量,在發芽12 h處理較0 h進行超聲波處理效果較好;超聲波處理只對發芽前期有作用,在中后期無顯著差異。糙米發芽過程中GABA含量變化主要取決于糙米自身發芽代謝。
2.4 發芽糙米中游離氨基酸含量變化
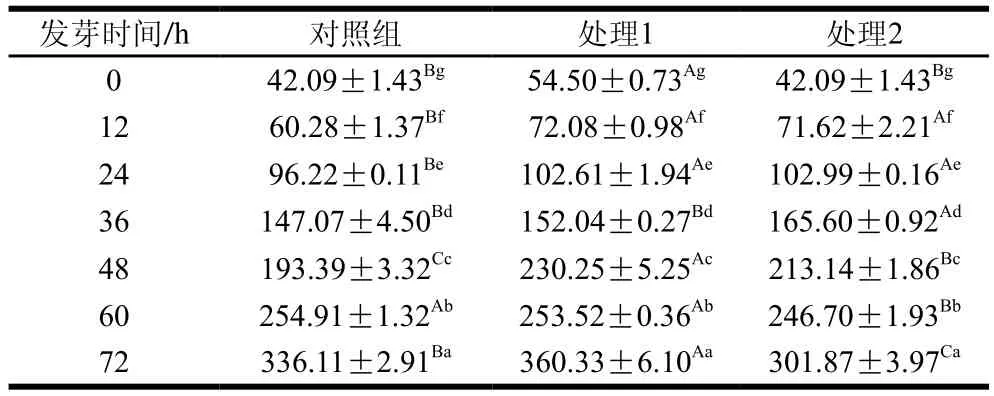
表2 超聲波處理發芽過程中游離氨基酸總含量的動態變化Table 2 Change in free amino acids contents during brown rice germination under ultrasonic treatment mg/100 g
浸泡預處理后糙米中游離氨基酸含量為42.09 mg/100 g。超聲波處理后游離氨基酸總量明顯增加,表明超聲促進糙米中蛋白質水解。隨發芽時間延長,大部分游離氨基酸(除天冬氨酸、絲氨酸外)含量持續增加,游離氨基酸總量由42.09 mg/100 g增加至336.11 mg/100 g。但2 種超聲波處理對發芽后期游離氨基酸含量增加無顯著促進作用。
2.5 GABA含量相關性分析

表3 超聲波處理糙米發芽過程中GABA、GAD活性和GA相關性分析Table 3 Correlation analysis between GABA, GAD activity, and GA during brown rice germination under ultrasonic treatment
為更清楚探明在浸泡和發芽過程中GABA、GAD及GA之間的相互關系,進行相關性分析見表3。3 種不同處理方式下的糙米在發芽過程中GABA與GA之間呈顯著正相關(P<0.05);但GAD活性與GABA和GA的相關性較低(P≥0.05)。GABA和GA呈顯著正相關是因為隨著時間延長兩者都是呈增加趨勢。
通常情況下,GABA含量隨GAD活性增加而增加,反之亦然[35]。本研究中GABA與GAD沒有顯著相關性,但Liu Liu等[36]發現GABA含量與GAD活性之間呈顯著正相關。Komatsuzaki等[37]發現外源添加GA可以調控GAD活性和GABA含量。Li Yan等[38]發現,在植物組織中GABA含量不僅受GAD活性的影響,還受到其他因素如GA含量、GABA轉氨酶和琥珀酸半醛脫氫酶影響。此外,Su Guoxing等[39]發現大豆中的GABA積累與多胺的氧化有關,受二胺氧化酶的活性增加影響。所以發芽糙米中GABA含量和GAD活性之間的關系還需進一步研究。
3 結 論
本研究借助超聲波逆境脅迫,采用浸泡發芽增加糙米中GABA含量,分析GA含量、GAD活性與GABA含量之間的關系。隨著發芽時間延長,GAD活性呈先增加后降低再增加的趨勢;在發芽0~48 h之間,對照組GAD活性均值為9.25 nmol/g,GABA含量增加到6 倍以上;在48~60 h之間,GABA含量增加量不明顯,GABA含量受GAD活性調節。在發芽過程中GABA和GA含量隨著發芽時間延長兩者均呈遞增趨勢,且二者之間呈顯著正相關(P<0.05)。
2 種超聲波處理方式在糙米發芽12 h后顯著增加了GABA含量,加快了GAD激活進程;顯著提高GA含量,達到對照組的2 倍以上;在發芽前期對GABA含量有顯著增加,對發芽后期GABA影響不明顯。在糙米發芽前期超聲波對其他種游離氨基酸含量有顯著促進作用;但2 種不同超聲波處理時間點之間的差異不顯著,糙米發芽過程中GABA含量變化主要取決于糙米自身發芽代謝。
綜上,發芽糙米中GABA形成受GAD活性調節,并與GA含量呈顯著正相關,超聲波處理對糙米發芽過程中GAD活性、GA含量有顯著影響,并有助于增加發芽糙米中GABA含量。然而,GA、GABA含量與GAD活性之間復雜的代謝途徑的關系還不清楚,仍需更進一步研究。
