嚙齒動物主動母性行為動態改變的神經機制*
2018-08-13 08:01:50張一帆齊星亮蔡厚德
心理科學進展 2018年8期
張一帆 齊星亮 蔡厚德,
(1南京師范大學心理學院,南京 210097) (2南京曉莊學院幼兒師范學院,南京 211171)
1 引言
主動母性行為(active maternal behavior)是雌性哺乳動物在哺乳期內有效照料幼崽的一種動機行為,涉及組織與調節一系列母性反應(maternal responsiveness)以適應幼崽發展需要的神經生物學過程(Pereira,2016; Pereira & Ferreira,2016)。哺乳動物幼崽在出生后并不能立即獨立生活,因而雌性的照料決定了幼崽的存活率和生理、行為發育狀況(Dobolyi,Grattan,& Stolzenberg,2014;Pereira & Ferreira,2016)。在人類中,母親的照料可以提供充分的刺激和訓練,使嬰兒在運動、感知、認知和社會情緒能力等方面得到良好發展(Lonstein,Lévy,& Fleming,2015)。另外,成為母親的過程也伴隨著巨大的生理改變和心理壓力,導致哺乳期女性容易出現產后抑郁(postpartum depression)等心理問題(Kim,Strathearn,& Swain,2016)。產后抑郁的母親在育幼時比較遲鈍、刻板和易怒,使嬰兒長大后傾向于淡漠和社交困難(Olazábal et al.,2013b)。因此,闡明哺乳動物主動母性行為及其神經機制,不僅可以為探究行為進化和早期發展開辟新的研究領域,也對人類母親產后抑郁等臨床問題的干預有借鑒意義。
考察主動母性行為的神經機制通常采用一些侵入性的方法(如腦組織損傷等),但這類實驗不能安全地運用于人類,因此研究者通常基于動物實驗的結果對主動母性行為的一般機制提出假設,以便采用非侵入性手段在人類中加以驗證(Lonstein et al.,2015)。在非人類哺乳動物中,實驗室大鼠(rat)和小鼠(mouse)等嚙齒動物的主動母性行為得到了最為詳盡的研究(Lonstein et al.,2015)。大鼠和小鼠的主動母性行為包括對幼崽的銜回(retrieving)、舔舐/理毛(licking/grooming)和嗅探(sniffing)等子成分(Afonso,King,Chatterjee,&Fleming,2009; Numan,2006)。在整個哺乳期(產后0~22天)內,雌鼠的主動母性行為經歷了從產后早期的發動(onset) (產后 0天)和維持(maintenance) (產后 0~11 天)階段,到產后晚期的衰退(decline) (產后 12~22天)階段的動態改變(Olazábal et al.,2013b; Pereira & Ferreira,2016)。在這一過程中,雌鼠能適應性地調整主動母性行為的表現,以滿足幼崽不同發育階段的需要(Pereira,2016)。較早的研究表明,主動母性行為的發動主要與分娩激素事件有關(Moltz & Wiener,1966); 在激素水平下降后,主動母性行為的維持轉而受到幼崽刺激的調節(Rosenblatt,1967;Reisbick,Rosenblatt,& Mayer,1975; Rosenblatt &Siegel,1981); 到了產后 12天左右,雌鼠與幼崽的接觸減少,主動母性行為開始走向衰退(Reisbick et al.,1975; Pereira & Morrell,2011)。近期,采用條件化位置偏好(conditioned place preference,CPP)任務的研究(Pereira & Morrell,2010; Wansaw,Pereira,& Morrell,2008)發現,嚙齒動物主動母性行為的動態改變與幼崽獎賞價值(incentive value)的階段性變化有關。值得關注是,還有大量研究(Cortés-Mendoza,Díaz De León-Guerrero,Pedraza- Alva,& Pérez-Martínez,2013; Numan et al.,2010; Benedetto,Pereira,Ferreira,& Torterolo,2014; Numan & Young,2016;Sabihi,Dong,Durosko,& Leuner,2014;Stolzenberg & Champagne,2016)探查了主動母性行為動態改變的神經機制。結果表明,幼崽獎賞價值變化引起的主動母性行為動態改變,不僅涉及分娩激素事件開啟下丘腦內側視前區(medial preoptic area,MPOA)?中腦腹側被蓋(ventral tegmental area,VTA)?伏隔核(nucleus accumbens,NA)?腹側蒼白球(ventral pallidum,VP)通路,而且需要杏仁核基底外側核(basolateral amygdala,BLA)的情緒性學習功能以及內側前額皮層(medial prefrontal cortex,MPFC)的行為選擇作用對上述通路進行實時調節。
本文首先利用CPP任務的行為學證據分析幼崽獎賞價值階段性變化與嚙齒動物主動母性行為動態改變的關系; 然后系統闡述產后早期主動母性行為的發動與維持以及晚期衰退的神經機制;最后探討未來需要研究的一些重要問題或方向。
2 CPP 任務的行為學證據
考察嚙齒動物產后不同階段的主動母性行為可以采用多種行為學方法,如母性行為測試(maternal behavior testing)、杠桿按壓(bar-pressing)和 CPP 等(Pereira & Morrell,2009; Olazábal et al.,2013a)。盡管母性行為測試能夠反映主動母性行為的變化,但難以評價幼崽對雌鼠的獎賞作用,因此需要將主動母性行為轉化為某種操作性行為,如針對幼崽的杠桿按壓或趨近行為(Olazábal et al.,2013a)。但是,雌鼠習得的按壓杠桿行為如果得不到幼崽的及時強化,繼續按壓的動機會自然減少,不便于長時間實驗觀察(Dalley,Cardinal,&Robbins,2004)。與此不同,雌鼠對幼崽的趨近行為會在不能獲得幼崽的情況下持續較長時間。因而,研究者(Pereira & Morrell,2009; Pereira &Morrell,2011)常用CPP任務,通過雌鼠趨近幼崽的行為考察幼崽作為一種具有高度獎賞價值的誘因對產后早晚期雌鼠主動母性行為的影響。
在CPP實驗中,首先訓練雌鼠將兩個具有不同知覺特征(如顏色)的場景分別與兩個刺激物建立聯結,然后移除刺激物,檢測雌鼠選擇趨向哪個場景(Tzschentke,2007)。在經典CPP任務中,兩個場景分別對應一個實驗刺激(如幼崽或可卡因)和一個中性的“控制”刺激(如生理鹽水) (Seip et al.,2008; Wansaw et al.,2008)。研究(Seip et al.,2008;Wansaw et al.,2008)發現,當對應幼崽的場景與中性場景比較時,產后早期約 80%的雌鼠選擇了對應幼崽的場景,而產后晚期 60%以上的雌鼠選擇了中性場景; 相比之下,當對應可卡因的場景與中性場景比較時,產后早期和晚期均有約 80%的雌鼠選擇對應可卡因的場景。而且,采用同步幼崽/可卡因 CPP任務(concurrent pup/cocaine choice conditioned place preference task)的研究支持以上結果。在同步CPP任務中,兩個場景分別對應幼崽和可卡因,以便在可卡因與幼崽競爭的條件下基于可卡因的劑量評價幼崽引發雌鼠主動母性行為的獎賞價值(Mattson,Williams,Rosenblatt,& Morrell,2001)。研究(Pereira & Morrell,2010,2011)發現,產后早期雌鼠對幼崽對應場景和可卡因(1.0 mg/kg)對應場景的偏好基本相同(約 40%);到了晚期,絕大多數(約 80%)雌鼠偏向可卡因對應場景,沒有雌鼠選擇幼崽對應場景。上述證據(Seip et al.,2008; Wansaw et al.,2008; Pereira &Morrell,2010; 2011)表明,產后晚期雌鼠對幼崽的偏好相對于早期明顯下降,但對可卡因的偏好一直較高。這提示產后晚期主動母性行為的衰退并不是一般性動機水平的減少所致,而是幼崽獎賞價值降低減弱了雌鼠的母性動機,因而不能有效引發主動母性行為(Mattson & Morrell,2005;Pereira & Morrell,2011)。
可見,主動母性行為動態改變與幼崽獎賞價值的階段性變化有關。問題是,從產后早期主動母性行為的發動和維持,到晚期的逐漸衰退,有哪些腦結構和神經通路參與了這一過程的加工與調節?其中涉及怎樣的神經機制?以下將梳理相關領域的研究證據加以闡述。
3 主動母性行為動態改變的神經機制
3.1 產后早期主動母性行為的發動
在分娩前后,雌鼠可以被幼崽的氣味所吸引。而且,分娩后雌鼠會立即開始對幼崽進行銜回、舔舐和嗅探(Lonstein et al.,2015)。主動母性行為的上述發動過程涉及分娩激素事件開啟MPOA-VTA-NA-VP通路,包括三個階段:(1)分娩激素事件激活 MPOA; (2) VTA-NA中多巴胺(dopamine,DA)水平的提高; (3) NA-VP的去抑制。圖1A說明主動母性行為發動的神經通路機制。
3.1.1 分娩激素事件激活MPOA
在未育雌性大鼠,來自嗅球(olfactory bulb,OB)的幼崽嗅覺信號通過杏仁核內側核(medial amygdaloid nucleus,MeA)與杏仁核皮質核(cortical amygdaloid nucleus,CA)向負責厭惡加工的下丘腦前核(anterior hypothalamic nucleus,AHN)傳遞,使雌鼠厭惡幼崽(Numan,2006)。但是,分娩激素事件能夠激活MPOA,導致嗅覺信號可以經MeA/CA傳至 MPOA,從而發動主動母性行為(Numan,2006)。雌鼠分娩時妊娠期的激素變化達到高峰,包括雌二醇(estradiol)水平上升、雌二醇/孕酮(progesterone)比例增加(Fleming,Ruble,Krieger,& Wong,1997; Lonstein et al.,2015)。證據(Numan,Rosenblatt,& Komisaruk,1977)表明,僅在妊娠后期雌鼠的MPOA中埋植雌二醇苯甲酸酯(estradiol benzoate),就可迅速激發主動母性行為。還有證據(Stolzenberg & Champagne,2016)表明,雌激素受體與雌二醇結合后可以協同共激活因子p160向基因的轉錄起始位點聚集染色質修飾酶(chromatinmodifying enzymes),打開MPOA中處于沉默狀態的多種受體基因,導致MPOA能夠對MeA/CA傳遞的嗅覺信號做出反應,進而作用于腦內獎賞系統使幼崽成為具有高獎賞價值的目標(Gagnidze,Weil,Faustino,Schaafsma,& Pfaff,2013; Numan,2006)。
3.1.2 VT A-NA中DA水平的提高
在幼崽氣味的刺激下,處于激活狀態的 MPOA可以興奮VTA的DA神經元,從而提升NA中的DA水平,使幼崽獎賞價值得以表達(Stolzenberg& Champagne,2016)。經典 CPP研究(Pereira &Morrell,2010)表明,MPOA 失活后,產后早期的雌鼠失去對幼崽的偏好,不再選擇對應幼崽的場景; 而控制組雌鼠中,有 80%的個體選擇了對應幼崽的場景。與此類似,VTA失活的雌鼠也完全失去了幼崽偏好; 而控制組有 50%選擇了對應幼崽的場景(Seip & Morrell,2009)。這些結果提示,MPOA激活后VTA釋放至NA中的DA水平可能決定了幼崽獎賞價值的高低(Seip & Morrell,2009)。研究者(Afonso et al.,2009)推測,NA中的DA活動可能是通過一種基線DA抑制(basal DA suppression)機制賦予幼崽高水平的獎賞價值。未接觸幼崽時,分娩雌鼠NA中的基線DA水平顯著低于未育雌鼠; 接觸幼崽后,分娩雌鼠 NA中的DA水平顯著超過未育雌鼠,DA水平的落差有利于雌鼠迅速將幼崽作為具有較高獎賞價值的目標進行關注(Afonso,Shams,Jin,& Fleming,2013;Olazábal et al.,2013a)。
3.1.3 NA-VP 的去抑制
NA中DA水平的快速上升,會觸發NA與控制動作程序的VP之間的去抑制機制,使DA活動轉化為主動母性行為(Numan & Young,2016)。證據(Sesack & Grace,2010)表明,NA中90%的傳出神經元是合成抑制性 γ-氨基丁酸(gamma-amino butyric acid,GABA)的中型多棘神經元(medium spiny neuron,MSN),可以通過D2受體接受DA的抑制性調節。但是,一些研究(D'Cunha,King,Fleming,& Lévy,2011; Parada,King,Li,& Fleming,2008)發現,MSN接受D2受體抑制劑處理并不明顯影響主動母性行為,接受催產素(oxytocin)受體抑制劑處理卻明顯干擾主動母性行為。進一步研究(Romero-Fernandez,Borroto-Escuela,Agnati,&Fuxe,2013)表明,MSN中除了含有D2受體之外,還有催產素受體,兩者以異聚體(heteromer)形式并存。在幼崽吮乳刺激下,來自PVN的催產素與催產素受體相結合,可以提升DA與D2受體的親和力(Romero-Fernandez et al.,2013)。雌鼠接觸幼崽前,NA中的DA和催產素水平都較低,MSN處于活動狀態,向 VP釋放 GABA,使其保持關閉,導致主動母性行為的動作程序不能啟動。接觸幼崽后,NA中的DA和催產素水平大幅提高。在催產素的調節下,大量DA與D2受體相結合,抑制了MSN的活動,使VP去抑制,導致主動母性行為的動作程序得以啟動,最終表現出主動母性行為(Numan & Young,2016)。
3.2 產后早期主動母性行為的維持
分娩數小時后,盡管雌鼠體內的激素水平已經大幅降低(Stolzenberg & Champagne,2016),但雌鼠仍會將幼崽作為高獎賞價值的目標,并維持頻繁的主動母性行為(Olazábal et al.,2013b)。在這一階段,不僅來自幼崽的嗅覺信號可以增加 MPOA中受體基因的表達,而且BLA通過情緒性學習的強化作用以及MPFC通過調節對幼崽的行為選擇都可以影響主動母性行為的表現。圖1B說明主動母性行為維持的神經通路機制。
3.2.1 嗅覺信號增加受體基因表達的作用
在主動母性行為維持階段,激素增加 MPOA中受體基因表達的作用會迅速被幼崽的氣味等刺激引起的感覺信號所接替(Dobolyi et al.,2014;Olazábal et al.,2013b)。研究(Stolzenberg & Champagne,2016)表明,長時間接觸幼崽能夠在大鼠和小鼠MPOA中引起一系列細胞間信號的反應,包括鈣信號(calcium signaling)、細胞外調節激酶的磷酸化(phosphorylation of extracellular regulated kinase,pERK)和轉錄因子環磷腺苷反應成分結合蛋白(cyclic AMP response element binding protein,CREB)水平的改變。而且,體內ERK下游產物或CREB的缺乏,會使小鼠主動母性行為受損(Jin,Blendy,& Thomas,2005; Kuroda et al.,2007)。與雌激素受體的作用類似,上述細胞間信號也可以增加 MPOA神經元中多種受體基因的表達(Cortés-Mendoza et al.,2013; Riccio,2010),這使來自幼崽的嗅覺信號也可以激活MPOA,并提升VTA-NA中的 DA水平,進而導致雌鼠繼續將幼崽作為高獎賞價值的目標(Pe?a & Champagne,2015; Numan & Young,2016)。
3.2.2 BLA 的情緒性學習作用
但是,嗅覺信號引起的MPOA激活水平可能較低,主動母性行為的維持還需要負責情緒性學習的BLA對NA-VP去抑制機制的調節(Numan &Young,2016)。證據(Lee,Clancy,& Fleming,1999)表明,損毀BLA會破壞大鼠以幼崽為目標的杠桿按壓行為。還有證據(Lee et al.,1999; Numan et al.,2010)顯示,BLA的損毀或失活不僅干擾了動物對情緒刺激的聯結性學習,而且破壞了主動母性行為。這提示BLA可以操作性地習得關于幼崽的情緒信息,從而促進雌鼠的主動母性行為。BLA及其鄰近區域對 NA和 VP各有一條興奮性谷氨酸(glutamate)神經投射(Numan et al.,2010; Numan &Young,2016)。通往NA的投射含有突觸前的D1受體,用藥物單獨抑制D1或MSN上的D2受體對主動母性行為影響較小,而同時抑制D1和D2受體可有效破壞主動母性行為的持續表現(Parada et al.,2008; Sesack & Grace,2010),提示主動母性行為的維持依賴于D1受體與MSN上D2受體的聯合作用。Numan和 Young (2016)認為,上述谷氨酸投射能夠興奮NA中的MSN,使VP保持抑制; 當DA水平上升時,DA與突觸前的D1受體結合,使谷氨酸投射的興奮降低,這會易化 NA對VP的去抑制。而且,在VP完全去抑制的前提下,BLA及其鄰近區域通往VP的神經投射可以產生超常激活,導致每次主動母性行為都能加強 VP中神經回路的突觸聯結,進而使主動母性行為得到強化(Numan & Stolzenberg,2009; Numan &Young,2016)。
3.2.3 MPFC 的行為選擇作用
MPFC是腦內整合情緒信息的高級中樞,可以利用從 BLA傳來的情緒信號對刺激的獎賞價值進行評估,從而制定行為決策(劉飛,蔡厚德,2010; Dilgen,Tejeda,& O’Donnell,2013)。MPFC包含邊緣下區(infralimbic region,IL)和邊緣前區(prelimbic region,PrL)等子區域(Pereira & Morrell,2011)。在操作性條件學習的一般過程中,IL負責按照已習得的關于刺激的獎賞價值進行反應,PrL則依據當前的獎賞價值進行反應(Dalley et al.,2004)。采用杠桿按壓范式的研究(Balleine &Dickinson,1998; Killcross & Coutureau,2003)表明,PrL失活(IL單獨起作用)的大鼠在獎賞不穩定的條件下仍然表現出穩定的條件化反應,失去了對當前獎賞價值的敏感性; 而IL失活(PrL單獨起作用)的大鼠反之,如果不能得到及時的獎賞,條件化反應立即減少。換句話說,IL以習慣模式起作用,PrL則以目標導向模式起作用(Dalley et al.,2004)。IL和PrL在產后早期主動母性行為中的調節作用與上述模式一致。同步CPP研究表明,PrL失活(IL單獨起作用)消除了雌鼠的可卡因偏好,使 70%雌鼠選擇了對應幼崽的場景; IL失活(PrL單獨起作用)消除了雌鼠的幼崽偏好,使80%雌鼠選擇了對應可卡因的場景; 而控制組中選擇兩類場景的雌鼠各占40% (Pereira & Morrell,2011)。由此來看,在主動母性行為的維持中,IL可能基于BLA已習得的情緒性經驗選擇幼崽,表現為習慣模式; PrL則可能基于BLA提供的當前的情緒性經驗選擇幼崽,表現為目標導向模式(Dalley et al.,2004; Olazábal et al.,2013b)。
研究顯示,MPFC的兩個子區域調節主動母性行為的神經通路有所不同,這可能與它們的上述功能特性有關。IL主要通過興奮性投射影響MPOA的激活(Olazábal et al.,2013b),這使主動母性行為傾向于以簡單和刻板的方式表現。相比而言,PrL參與主動母性行為調節的機制則比較復雜。PrL中存在至少兩組與主動母性行為有關的神經元。第一組是向MPOA投射的興奮性神經元(Olazábal et al.,2013b),含有催產素受體,接受來自PVN的催產素投射(Sabihi et al.,2014)。證據(Febo,Numan,& Ferris,2005; Sabihi et al.,2014)表明,幼崽的吮乳行為會引起PVN分泌催產素和MPFC的激活,且用催產素受體拮抗劑處理 PrL會損害雌鼠的主動母性行為,提示來自幼崽的感覺刺激可以通過催產素影響 PrL中的興奮性神經元,進而提高MPOA的激活水平。第二組是向NA中MSN投射的興奮性谷氨酸神經元,含有突觸前的D2受體(Sesack & Grace,2010)。當NA中DA水平上升時,DA與 D2受體結合會削弱 PrL對MSN的興奮性影響,進而易化 VP的去抑制(Olazábal et al.,2013b; Sesack & Grace,2010)。可見,PrL既可以通過MPOA調控DA的釋放,又可以通過 VP去抑制參與主動母性行為的調節,從而使雌鼠的主動母性行為表現更為靈活。
3.3 產后晚期主動母性行為的衰退
與產后7天相比,產后13天左右雌鼠舔舐幼崽的頻率從每分鐘1次下降到0.5次,而初次銜回的潛伏期從約5秒上升到88秒; 產后22天前后幼崽斷奶時,雌鼠的主動母性行為完全終止(Pereira & Morrell,2009; 2011)。產后晚期雌鼠主動母性行為的上述衰退過程與幼崽的獨立性需求不斷提高有關(Pereira & Ferreira,2016)。證據(Pereira & Morrell,2009; Olazábal et al.,2013b)表明,產后晚期幼崽的視覺、聽覺和運動能力明顯提升,傾向于獨自離巢覓食,并且會抗拒雌鼠的銜回。研究者(Olazábal et al.,2013b)認為,隨著幼崽獨立性的提高,雌鼠接受到的幼崽感覺刺激會逐漸減少,在主動母性行為中的操作性情緒學習效率也不斷降低。伴隨這一過程,BLA難以繼續習得來自幼崽反應的情緒性信號,導致 VP的去抑制和主動母性行為受到的強化作用減弱,使產后晚期的主動母性行為逐漸減少(Pereira &Ferreira,2016; Olazábal et al.,2013b)。
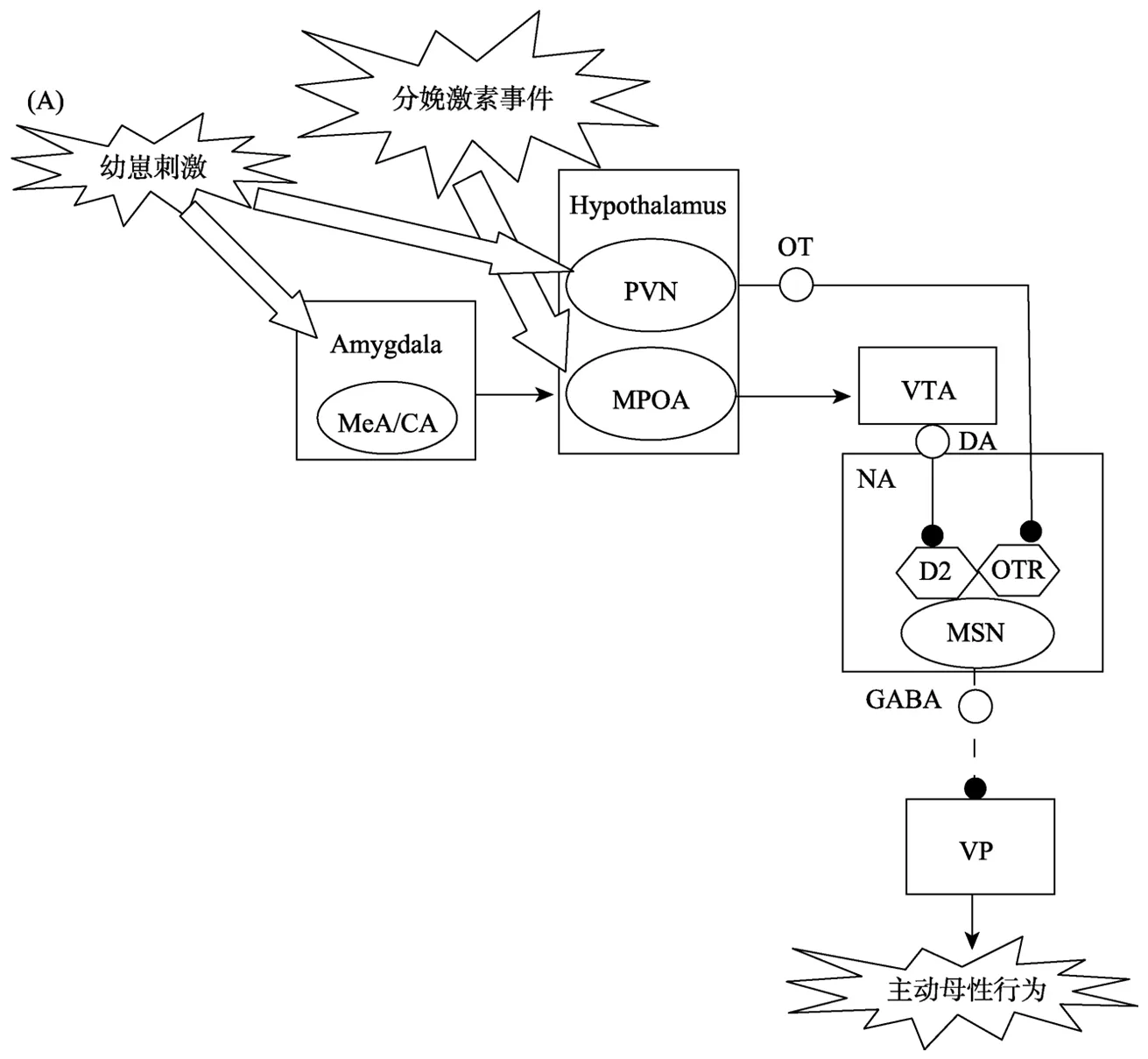
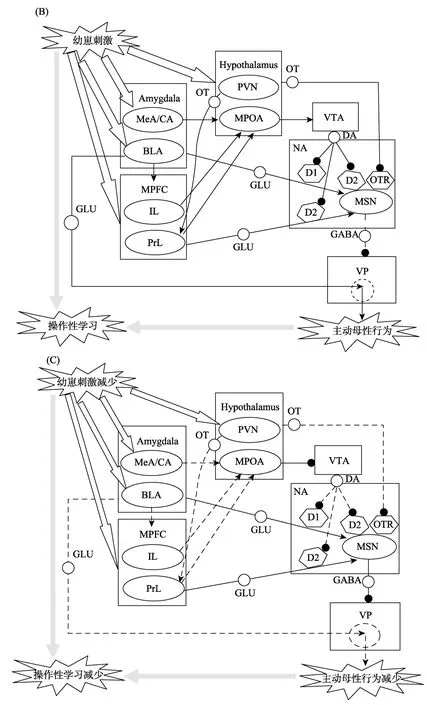
圖1 主動母性行為動態改變的神經通路機制
MPOA的功能在產后晚期也發生了重大改變(Olazábal et al.,2013b)。研究(Pereira & Morrell,2009)發現,產后晚期 MPOA 失活使雌鼠對幼崽的銜回、舔舐和嗅探等行為的頻率大幅提升,提示MPOA在產后晚期可能對主動母性行為起抑制作用。黑色素聚集素(melanin-concentrating hormone,MCH)參與了 MPOA對產后晚期主動母性行為的調節。研究(Benedetto et al.,2014)發現,向MPOA灌注 MCH能夠損害主動母性行為,而產后晚期MPOA中 MCH 的水平不斷上升,在斷奶時達到頂峰,這與主動母性行為減少的趨勢相一致。Pereira和Ferreira (2011)認為,產后晚期MPOA可能會抑制VTA中DA神經元的活動,從而降低幼崽的獎賞價值,也會使主動母性行為逐漸減少。
另外,MPFC的兩個子區域在產后晚期對主動母性行為的調節作用與維持階段有明顯不同。證據表明,產后晚期IL對MPOA的興奮性投射明顯減弱(Olazábal et al.,2013b),IL的失活對主動母性行為不產生明顯影響(Pereira & Morrell,2011;Olazábal et al.,2013b),提示IL可能不再參與主動母性行為的調節。證據(Pereira & Morrell,2009;2011)還表明,產后晚期PrL的失活會損害雌鼠主動母性行為的表現,而 MPOA失活(PrL完好)的雌鼠主動母性行為的表現則大幅提升,在受到幼崽抗拒時也不能對主動母性行為進行及時調整,提示PrL在產后晚期只以一種刻板的方式維系一定的主動母性行為。
可見,產后晚期主動母性行為的衰退是雌鼠對幼崽發展需要變化的一種響應。在這一過程中,BLA的情緒性學習效率下降對產后產晚期主動母性行為的衰退可能起主導作用。有研究者(Olazábal et al.,2013b)推測,產后晚期MPFC的功能改變可能與來自 BLA的情緒性學習信號減少有關,而MPFC對MPOA的興奮性投射減弱又可能導致后者表現出抑制主動母性行為的作用。圖1C說明主動母性行為衰退的神經通路機制。
4 總結與展望
綜上所述,哺乳期嚙齒動物的主動母性行為會經歷從產后早期的發動和維持到晚期衰退的動態改變,涉及雌鼠調整自身的母性反應以適應幼崽獎賞價值階段性變化的一系列復雜神經機制,與多條神經通路、多種心理成分和多個節點調節的神經加工有關。首先,MPOA-VTA-NA-VP是一條控制主動母性行為的基本通路。在主動母性行為發動階段,分娩激素事件可以激活 MPOA,進而開啟這一基本通路; 在維持和衰退階段,來自BLA和MPFC的調控通路可以對基本通路進行實時調節,進而影響主動母性行為的表現。其次,以上通路機制包含多種成分的心理加工。在基本通路中,MPOA是加工母性動機和幼崽獎賞價值的中樞,VTA-NA-VP則是控制動機行為程序的一般通路。在調控通路中,BLA可以操作性地學習有關幼崽的情緒信息,使主動母性行為得以強化;MPFC則可以利用來自BLA的情緒性經驗,通過獎賞評估調節雌鼠對幼崽的選擇偏好,使主動母性行為在產后早晚期表現出不同的模式。最后,來自BLA和MPFC的調控通路可以通過多條神經投射對基本通路的多個節點進行調節。MPOA是基本通路的始端節點,其激活可以控制VTA釋放DA,使雌鼠能夠將幼崽作為高獎賞價值的目標;NA-VP則是基本通路的末端節點,NA中DA水平的提高可以使 VP去抑制,導致主動母性行為得以表現。在主動母性行為的維持階段,BLA既可以通過對 NA的興奮性投射易化通路末端 VP的去抑制,又可以直接向 VP發放興奮性神經投射,這兩條投射在基本通路末端的共同作用可以使主動母性行為得到有效強化; MPFC中的IL只興奮基本通路始端的MPOA,而PrL則既可以興奮始端的MPOA,也可以促進末端VP的去抑制,IL與PrL的協同作用使主動母性行為不僅具有穩定性,又有靈活性。在主動母性行為的衰退階段,BLA對通路末端的去抑制和強化作用明顯減退,通路始端 MPOA功能的轉變使幼崽獎賞價值降低,MPFC對通路的調節作用也逐漸撤出,最終導致主動母性行為逐漸關閉。
然而,盡管調控哺乳期嚙齒動物主動母性行為動態改變的神經機制已初見端倪,但仍有一些重要問題或方向亟待深入探究。
第一,MPOA中抑制性神經元的作用。MPOA包含背側區(dorsal MPOA,MPOAd)和腹外側區(ventrolateral MPOA,MPOAvl) (Olazábal et al.,2013b)。證據(Olazábal et al.,2013b)表明,MPOAd中分布著向腦內厭惡系統投射的抑制性神經元,MPOAvl中則分布著向 MPOAd投射的抑制性神經元。還有證據顯示,前一類是協同表達甘丙肽(Galanin)的 GABA神經元,可以抑制雌鼠的主動回避幼崽行為(Wu,Autry,Bergan,Watabe-Uchida,& Dulac,2014; Zha & Xu,2015); 后一類則可能是協同表達MCH的GABA中間神經元,可以抑制MPOA對VTA的興奮性傳出,與產后晚期MPOA轉變為抑制性角色有關(Benedetto et al.,2014;Olazábal et al.,2013b)。可見,這兩類抑制性神經元可能通過不同的功能參與了產后早晚期主動母性行為的調節。進一步研究需要關注如下問題:(1) MPOAd中的GABA傳出神經元如何與興奮性傳出神經元相互協同以調節產后早期主動母性行為?(2)產后晚期MPOAvl中的GABA中間神經元是否會通過抑制MPOAd中的GABA傳出神經元使雌鼠主動回避幼崽?這在主動母性行為衰退的過程中又扮演怎樣的角色?(3)產后不同階段MPFC的功能改變是否也會影響MPOA中抑制性神經元的作用?
此外,腦內的五羥色胺(serotonin,5-HT)系統對MPOA的投射也可能參與主動母性行為的調控(陳磊磊等,2017),這是否會通過MPOA中的兩類GABA 神經元起作用,尚待研究。
第二,NA不同子區域的作用。NA分為殼(shell)區和核(core)區。證據表明,電損毀殼區可破壞產后早期主動母性行為(Li & Fleming,2003),但采用布比卡因(bupivacaine)將 NA 整體失活卻沒有明顯影響(Pereira & Morrell,2011),這提示NA 的殼區和核區的功能及其相互關系可能比較復雜。證據(Sesack & Grace,2010; Root,Melendez,Zaborszky,& Napier,2015)還表明,殼區和核區的MSN分別有投射到 VP的腹內側區(ventromedial VP,VPvm)和背外側區(dorsolateral VP,VPdl),其中,殼區到 VPvm的投射是主動母性行為去抑制的通路,而核區到VPdl的投射是調節自主運動去抑制的通路。NA整體失活導致的殼區受損可能使主動母性行為去抑制出現困難,而核區受損則會導致自主運動過度釋放(Li & Fleming,2003;Pereira & Morrell,2011)。研究者(Nicola,2007; Pereira& Morrell,2011; Olazábal et al.,2013a)推測,較高的自主運動水平有利于主動母性行為的表現,從而在一定程度上彌補了殼區受損帶來的損害,使行為的輸出結果可能保持不變。值得注意的是,NA殼區受損妨礙主動母性行為的機制還不清楚。證據(Numan,2006)顯示,NA殼區除了有一條對VPvm 的去抑制通路之外,還有一條表達腦啡肽(enkephalin)的神經通路可以易化 BLA的谷氨酸投射對 VP的直接激活作用,進而增進對主動母性行為的強化。未來研究可以通過 NA核區的單獨損毀或失活探查自主運動如何參與產后早晚期主動母性行為的表現,還可以考察NA殼區向VP投射的兩條通路如何在產后早晚期相互作用及其對主動母性行為動態改變的影響。
第三,MPFC中前扣帶回(anterior cingulate,ACg)的調節功能。研究表明,在妊娠前損毀雌鼠MPFC中包含ACg在內的廣泛區域,會使分娩后雌鼠銜回和舔舐幼崽的頻率降低、秩序性受損(Afonso,Sison,Lovic,& Fleming,2007); 而單獨失活 ACg主要損害主動母性行為的秩序性(Pereira & Morrell,2011)。可見,與PrL和IL不同,ACg可能主要通過調節多種行為成分的時間秩序來組織母性反應。有證據(Kesner,2000)表明,損毀ACg破壞了雌鼠對按時間順序排列的空間線索記憶。而且,ACg與負責母性動機加工的MPOA和情緒性學習的BLA均有神經聯系(Olazábal et al.,2013b)。今后的研究可以關注雌鼠在復雜的育幼環境中如何通過ACg的上述神經聯系計劃和組織適當的主動母性行為。
第四,幼崽聲音信號識別與主動母性行為的動態改變。研究(Banerjee & Liu,2013; Marlin,Mitre,D’Amour,Chao,& Froemke,2015)表明,幼崽聲音信號的改變不僅會引起雌鼠聽覺皮層神經元電活動的可塑性變化,也使MPOA的激活水平發生了相應改變,提示聽覺皮層對幼崽聲音信號的識別可能參與了主動母性行為的調節。需要注意的是,來自幼崽的聲音信號是一種間接的或遠距離的刺激,雌鼠對其意義的識別可能涉及更復雜的認知加工過程,但幼崽聲音信號調節主動母性行為動態改變的神經機制還不清楚。研究者(Banerjee & Liu,2013)推測,聽覺皮層可以通過杏仁核(amygdala)和海馬腹側下托(ventral subiculum;vSub)與 MPOA 相互作用,完成對幼崽聲音信號的識別及其情緒性和情境意義的學習與記憶。在此基礎上,聽覺皮層的神經信號不僅可以直接通過MPOA影響VTA-NA中的DA水平,還可以經MPFC到達 NA,調節主動母性行為的去抑制過程。未來研究可以建立產后早晚期的幼崽聲學模型(acoustic model) (Banerjee & Liu,2013),系統考察主動母性行為從發動、維持到衰退的過程中聽覺皮層神經可塑性變化與上述兩條神經通路功能聯結的形成與消退之間的關系,也可以比較幼崽聽覺信號與其他感覺信號的識別在調節雌鼠主動母性行為動態改變中是否扮演不同角色。
最后,哺乳期人類母親親子互動行為的神經機制。盡管哺乳期的人類母親并不表現出與嚙齒動物主動母性行為成分(銜回、舔舐和嗅探等)完全對應的行為表現,但她們在照料嬰兒過程中會利用多種感覺通道的信息和主動的行為反應與嬰兒互動(包括相互凝視、身體接觸和聲音交流),以回應嬰兒的需求(Strathearn,2011; Kim et al.,2016)。一些功能性磁共振成像(functional magnetic resonance imaging,fMRI)研究(Swain et al.,2008;Seifritz et al.,2003; Laurent & Ablow,2012)檢查了人類母親聽到自己嬰兒哭泣時的神經功能反應,發現紋狀體(striatum)、下丘腦(hypothalamus)、杏仁核、MPFC和前扣帶皮層(anterior cingulate cortex)等腦區的激活明顯增強。這提示,母親識別自己嬰兒聲音信號所激活的腦區與嚙齒動物主動母性行為所涉及的腦區十分相似,也包含了負責動機、情緒和行為組織等多種心理加工成分的神經結構。但是,人類母親與嬰兒的親子互動還可能涉及高級的認知執行功能。例如,親子互動質量高的母親在觀看自己的嬰兒玩耍視頻時,與自我覺知和認知靈活性有關的額中回(middle frontal gyrus)和額下回(inferior frontal gyrus)的激活水平更高,且紋狀體的 NA與這兩個腦區的功能聯結增強(Atzil,Hendler,& Feldman,2011; Lonstein et al.,2015),提示人類母親的親子互動依賴于皮層通過一系列高級功能與皮層下結構相互影響。另外,產后抑郁或有抑郁傾向的母親上述腦區的神經活動與正常母親有明顯差異,如聽到自己嬰兒聲音引起的NA激活低于正常母親,而左側MPFC的激活又高于正常母親(Swain et al.,2008; Laurent& Ablow,2012),這也許可以為產后抑郁母親的腦功能定向臨床干預提供借鑒。然而,在 fMRI的實驗中被試需要固定身體姿勢,因此不可能考查真實的親子互動過程中人類母親的腦功能活動情況(Lonstein et al.,2015)。未來研究有必要引入可以允許母親在親子互動中完成某些身體動作的腦功能技術,如腦電圖(electroencephalography,EEG)和功能性近紅外光學成像(functional near-infrared spectroscopy,fNIRS)等,并將來自EEG、fNIRS和fMRI的研究證據相互參照,進一步探究哺乳期人類母親親子互動行為的神經機制(Kim et al.,2016)。
