放牧干擾下高寒草甸物種、生活型和功能群多樣性與生物量的關系
2018-08-09 08:32:06牛鈺杰楊思維王貴珍杜國禎花立民
生態學報 2018年13期
牛鈺杰,楊思維,2,王貴珍,劉 麗,杜國禎,花立民,*
1 甘肅農業大學草業學院/草業生態系統教育部重點實驗室/中-美草地畜牧業可持續發展研究中心, 蘭州 730070 2 畢節市畜牧獸醫科學研究所, 畢節 551700 3 蘭州大學生命科學學院, 蘭州 730030
青藏高原東緣約有1900種植物分布[1],且每平米物種豐富度最高可達50種[2]。高寒草甸作為青藏高原主體生態系統類型,是畜牧生產的主要資源載體,也是植物多樣性保存相對完好的地域之一[3- 4]。多樣性與生產力之間的關系是高寒草甸最主要的研究熱點之一[5- 9]。Tilman等[10]創造性地提出功能多樣性比物種多樣性更能決定生態系統生產力,進而將作為物種組合類群多少的功能多樣性引入到多樣性與生產力關系的研究之中。但Thompson等[11]和馬文靜等[8]部分學者也發現功能多樣性與生產力無關。物種多樣性和功能多樣性究竟哪個更能決定生產力的大小,一直爭議很大[6- 9]。
這種爭議一方面是由于不同研究者選擇的多樣性指數不同[12]。現今多樣性指數的種類多種多樣[12]。按組織層次不同可以將多樣性分為物種多樣性和功能多樣性,而物種和功能多樣性按其強調多樣性組分不同,又均可以分為豐富度[13]、優勢度[14]和異質性測度[15]的統計指標。王長庭等[16]選擇豐富度指數、Simpson指數和均勻度指數研究物種多樣性與生物量之間的關系,發現不同多樣性指數與生物量之間相關系數差別很大。單就功能多樣性而言,由于植物功能特征不是絕對單一的,所以對植物功能的界定就會有不同的理解和劃分方法,導致人為主觀性較強[6,17]。根據研究目的不同,研究者選用不同的功能分類方法。而以生活型態[18]和經典的林奈生物學分類法[6,8]為原始功能分類依據較為常見。王國杰等[19]使用不同的功能多樣性指標對內蒙古草原群落進行研究發現,水分生態類型多樣性比物種和生活型多樣性更能反映與生產力之間的關系,從而得出多樣性與生產力的關系受多樣性指數選擇的影響。因此,在生態學研究中,多樣性與生產力關系因研究者選擇不同功能多樣性和物種多樣性指數而異。
另一方面,研究者在野外植物調查中普遍使用地上現存量直接表示生產力,來研究其與多樣性之間的關系[4,8- 10,19]。但除了在受控試驗條件下可以用草地生物量準確折算出草地的初級生產力外[20],草地植物現存量極大地受放牧干擾的影響[19]。放牧作為所有草地類型最主要且不可避免的干擾方式[3],其對植物群落多樣性和生產力的影響取決于干擾的強度特征和時間累積效應[21]。而不同放牧強度對植物群落的影響是在空間尺度上進行比較,具有瞬時性[3]。但因為草地植物本身具有補償性生長和每年重新萌發的特性,以放牧年份為代表的時間累積效應對植物多樣性和生物量的影響就很模糊[21- 22]。導致放牧干擾下多樣性與生產力之間的關系更加不確定,鑒于放牧干擾不可避免且普遍存在,那么研究不同放牧強度長時間持續干擾下多樣性與生物量之間的關系,對多樣性與生產力之間的相互預測就具有更多的科學和現實意義。
基于上述分析,本研究在青藏高原東北緣高寒草甸選擇6塊不同放牧強度樣地,連續4 a進行物種、生活型和功能群多樣性調查與生物量監測,研究不同放牧強度作用下多樣性與生物量之間的關系。以期明晰不同層次的多樣性對放牧干擾在空間強度特征與時間尺度累積作用下的敏感性以及多樣性和生物量之間的相互關系對放牧干擾的響應,最終為有效評估高寒草甸在空間和時間尺度上的植物多樣性變化和建立合理的放牧制度提供科學依據。
1 材料與方法
1.1 研究區概況
研究區位于青藏高原東北緣,植被類型為高寒嵩草草甸。行政上隸屬甘肅省天祝縣抓喜秀龍鄉(37°11′—37°14′ N,102°40′—102°47′ E),海拔2958 m。年均氣溫-0.1℃,>0℃的年積溫為1380℃。年均降水量416 mm,主要集中在7—9月,占全年降雨量的76%。無絕對無霜期,植物生長期120—140 d。土壤類型為高山草甸土,土層厚度40—80 cm。優勢物種為垂穗披堿草(Elymusnutans)、冷地早熟禾(Poacrymophila)、西北針茅(Stipakrylovii)、矮生嵩草(Kobresiahumilis)和陰山扁蓿豆(Melissitusruthenica)等。
1.2 試驗設計
在系統考察試驗地植被情況和調查當地實際載畜量的基礎上,選擇地勢平坦、草地類型一致、優勢種為垂穗披堿草和大針茅的總面積共10 hm2樣地,此樣地在圍欄放牧試驗之前以3.69羊單位/hm2進行輕度的自由放牧管理。根據當地的草地生物量計算草地理論承載的羊單位頭數為4.5—5.0羊單位/hm2,進而設置6個放牧強度區(放牧強度=羊單位數×放牧時間放牧面積)開展控制放牧試驗(表1),選擇體重相近,約為45 kg的甘肅高山細毛羊為放牧家畜。以全年每天連續放牧的方式進行放牧管理。放牧作用于基礎條件一致的草地,消除了地形和氣候因素的干擾。以當地平均放牧強度作為基準,上下浮動設置放牧強度梯度,并定義高于當地平均放牧強度(中等放牧強度)的樣地4、樣地5和樣地6為高放牧強度樣地,低于當地平均放牧強度的樣地1、樣地2和樣地3為低放牧強度樣地。于2012年8月,開始圍欄放牧,并連續4 a對樣地植物群落進行定點調查。

表1 研究樣地信息
不同小寫字母表示不同放牧強度樣地各項指標差異顯著(P<0.05)
1.3 實地取樣
1.3.1 取樣規則
2012—2015年,每年8月15號左右進行樣地植物群落調查。采用系統隨機取樣法,將每個樣地規則地分為9個小區。在每個小區內隨機地設置一個樣方,取樣時避開樣地群落邊緣,即每個樣地重復取樣9次。4年共調查樣方216個。在2012年首次取樣后將取樣點固定。之后每年在固定取樣點調查取樣。
1.3.2 調查和計算方法
群落內每一物種蓋度:用0.25 m2樣方,針刺100次;物種高度:每一種植物隨機測量10次;物種頻度:每樣方附近隨機拋樣圓20次,記錄每一物種頻度。用最大值標準化法計算每一物種高度、蓋度和頻度的相對值。物種重要值計算公式為:IV=(A+B+C)/3。式中:A為相對蓋度;B為相對頻度;C為相對高度。用各生活型和功能群植物的重要值之和表示生活型和功能群重要值。根據植物生活形態和功能性劃分方法分別將物種劃分為6個生活型(叢生型、根莖型、蓮座型、匍匐型、一年生、直立型)[23]和4個功能群(禾本科、莎草科、豆科、雜類草)[6,8,24]。在選擇生活型時,因為根莖型植物的耐牧性和一年生植物對放牧作用的敏感性,因而將其優先從植物中篩選出來,其余4種生活型根據莖的形態(叢生、蓮座、匍匐、直立)也可進行完全劃分。功能群的選擇按照經典的林奈生物學分類法進行。不同組織層次(物種、生活型和功能群)的多樣性指數均選取豐富度、均勻度和優勢度作為測度指標[12,25]。物種、生活型和功能群豐富度均用0.25 m2樣方,分別以其中出現的物種數、生活型種類和功能群種類表示。再用重要值計算樣方群落的物種、生活型和功能群均勻度(Evenness指數)[15]和優勢度(Berger-Parker指數)[14]。計算公式為:
Evenness指數:
Berger-Parker指數:
B=Nmax/N
式中,S為每個樣方總的物種數、生活型種類和功能群種類;N為S個全部物種、生活型和功能群的相對重要值之和;Ni為第i個物種、生活型和功能群的相對重要值,Nmax為相對重要值最大值的物種、生活型和功能群。
1.4 數據分析
對放牧強度作用下物種、生活型和功能群多樣性(豐富度、均勻度和優勢度)做隨放牧年份累積作用下的重復測量方差分析(LSD法進行顯著性檢驗),并對變量進行球形檢驗,用Greenhouse-Geisser法對自由度進行校正。用Pearson相關性分析法對各多樣性指數的相互關系進行分析。采用線性回歸法分析物種、生活型和功能群多樣性(豐富度、均勻度、優勢度)與生物量之間的關系,設定P值為0.05,用決定系數R2,解釋生物量對各多樣性的貢獻大小。所有統計分析均使用SPSS 19.0軟件進行。
2 結果與分析
2.1 地上生物量在放牧干擾下的變化
生物量隨放牧強度和放牧年份均發生顯著變化。2012年的生物量在不同放牧強度區無顯著變化。而2013—2015年,生物量隨放牧強度的增加均表現出顯著降低的變化趨勢。而不同放牧強度區,生物量隨放牧年份的整體變化規律不一致,但2013—2015年不同放牧強度區生物量均低于圍欄放牧前2012年(表2)。

表2 地上生物量在不同放牧強度樣地隨放牧時間的變化
括號內為橫向比較,括號外為縱向比較。顯著水平:P<0.05
2.2 放牧強度作用下多樣性隨年份的變化
對于豐富度,放牧年份對物種、生活型和功能群豐富度的影響均達到顯著水平。而不同放牧強度之間豐富度無顯著差異(P>0.05)。放牧年份和放牧強度對物種豐富度的變化具有交互作用,放牧年份對物種豐富度的作用隨放牧強度的不同而異,表現為低強度放牧樣地(Plot1、Plot2、Plot3)的物種豐富度高于高強度放牧樣地(Plot4、Plot5、Plot6)(圖1)。對于均勻度,放牧年份對物種、生活型和功能群均勻度的影響均達到顯著水平。放牧年份和放牧強度對物種、生活型和功能群均勻度均無交互作用,放牧年份對均勻度的作用隨放牧強度無顯著變化(圖1)。對于優勢度,放牧年份對物種和功能群優勢度的影響達到顯著水平,而對生活型優勢度無影響。放牧年份和放牧強度對物種、生活型和功能群優勢度均無交互作用,放牧年份對優勢度的作用隨放牧強度無顯著變化(圖1)。

圖1 不同放牧強度下豐富度、均勻度和優勢度隨放牧年份的變化Fig.1 Variation of richness, evenness, Berger-parker under the grazing intensity with different grazing year
2.3 放牧干擾下多樣性指數之間的相關關系
在連續放牧干擾下,所有多樣性指數中,僅物種豐富度與其余多樣性指數均表現顯著的相關關系,其與物種均勻度、物種優勢度、生活型優勢度和功能群均勻度顯著負相關,與生活型豐富度、生活型均勻度、功能群豐富度和功能群優勢度顯著正相關。生活型均勻度與生活型優勢度的相關性最高,其相關系數為-0.818。其次為功能群均勻度和功能群優勢度,相關系數為-0.786(表3)。
2.4 不同放牧強度區生物量與豐富度、均勻度和優勢度的關系
2.4.1 豐富度與生物量的關系
低強度放牧樣地的Plot2中物種豐富度隨生物量增加而顯著降低(R2=0.140)。其余放牧強度樣地的物種、生活型和功能群豐富度均與生物量無關(圖2)。
2.4.2 均勻度與生物量的關系
低強度放牧樣地的Plot1中,生活型均勻度隨生物量的增加而顯著降低。而低強度放牧樣地的Plot2中,物種均勻度隨生物量的增加而顯著升高。高強度放牧樣地的Plot5中,功能群均勻度隨生物量的增加而顯著降低。在高強度放牧樣地(Plot4、Plot5、Plot6)中,生活型均勻度均隨生物量的增加而顯著降低。其余多樣性指數與生物量無關(圖3)。

表3 多樣性指數的相關關系
*P<0.05; **P<0.01

圖2 不同放牧強度區豐富度與生物量的關系Fig.2 Relationship between richness and biomass in different plots
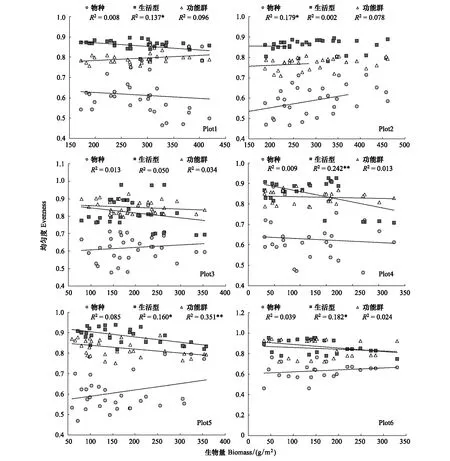
圖3 不同放牧強度區均勻度與生物量的關系Fig.3 Relationship between evenness and biomass in different plots
2.4.3 優勢度與生物量的關系
在低強度放牧樣地的Plot1中,功能群優勢度隨生物量的增加而顯著降低。在高強度放牧樣地(Plot4、Plot5、Plot6)中,生活型優勢度均隨生物量的增加而顯著升高。其余多樣性指數與生物量無關(圖4)。

圖4 不同放牧強度區優勢度與生物量的關系Fig.4 Relationship between Berger-parker index and biomass in different plots
3 討論與結論
3.1 物種豐富度可作為生物多樣性測度的代表性指標
植物多樣性指數是研究群落物種組成結構和定量比較的重要指標[12]。生態學家為簡化群落物種研究的復雜性[16]和強調具體多樣性測定中的可操作性,提出了功能多樣性的概念[17]。但究竟物種多樣性和功能多樣性哪個多樣性指標可以更好、更快捷地對群落進行定量比較仍然不甚清晰。原因是生態學研究中有很多多樣性指數可供選擇[12]。功能多樣性作為物種組合類群的多少,不同的研究者有不同的劃分方法[6 8]。但以生活型態[18,23]和經典的林奈分類學方法[6,8,24]進行功能分類被大多數研究者所認可。而不論是物種多樣性還是功能多樣性,多樣性統計量均可分為豐富度[13]、優勢度[14]和均勻性[15]測度。因此,面對復雜、種類繁多的多樣性指數[12],明晰放牧干擾下不同層次多樣性之間的相互關系,找出放牧干擾下多樣性的代表性測度指標就有很重要的實踐意義。本研究在青藏高原東緣的高寒草甸連續4 a進行放牧干擾下物種、生活型和功能群多樣性之間相關關系的研究。結果表明,物種豐富度與其余多樣性指數之間均呈顯著相關關系,尤其與功能多樣性表現極強的相關性。物種豐富度作為某一選定集聚群落內已知物種的種數,其簡單有效的定義和具體實踐的可操作性[25],可將其作為生物多樣性比較的代表性測度指標。雖然物種豐富度并不意味著它是生物多樣性的唯一測度,但因為越是物種豐富的群落就存在越多的功能類群,本研究中物種豐富度與功能多樣性也表現出顯著相關關系,其對生態系統功能多樣性強大的預測潛能,也引起研究者的關注[25]。但因為研究者所采用的物種概念不同就會使豐富度的測度受到很大的影響[26]。因而,在物種概念較為一致的情況下,物種豐富度可以作為生物多樣性測度的代表性指標。
3.2 放牧強度對物種豐富度的影響隨放牧年份有累積作用
物種多樣性作為一門比較性科學[12,25]。研究者通常想知道在空間尺度上是否一個地區的多樣性超過另一個地區?或特定地理區域的自然群落在時間演替過程中多樣性如何變化[25]?對于放牧干擾下的草地生態系統,研究者較多關注放牧強度或時間尺度單一因素影響下的多樣性比較[3]。但對于放牧而言,干擾的空間強度特征及其在時間尺度上的累計效應是同時存在的[12]。而植物本身又具有補償性生長和每年重新萌發的特性,因而研究哪些多樣性指數隨放牧強度在時間尺度上具有累積效應,才能了解放牧干擾對多樣性影響的持續性。本研究分析了放牧強度在放牧年份的累積作用下對物種、生活型和功能群多樣性的作用大小。結果表明,放牧年份和放牧強度僅對物種豐富度具有交互作用,也就是說放牧強度對物種豐富度的作用隨放牧年份具有累積作用。而其余層次的多樣性指數雖都隨放牧年份延長發生顯著變化,但沒有產生隨放牧強度變化的時間累積效應。這就能夠解釋以物種豐富度作為群落物種多樣性的代表性測度指標時,放牧強度能夠造成多樣性變化的原因。因為物種豐富度隨放牧時間的累積在不同的放牧強度區發生明顯的分異現象,低放牧強度區的物種豐富度高于高放牧強度區。物種豐富度(即植物種類的多少)作為多樣性測度中最直接的指標,累積效應的產生意味著研究者必須用長遠的眼光來看待某一時刻放牧強度的差異,關注放牧對多樣性產生影響的長時間效應,而不只是短期的結果。
3.3 高強度放牧干擾下生活型多樣性與生物量依然顯著相關
植物功能類型是具有確定功能性狀的一系列植物的組合,是生態學家為研究植被應對環境變化而引入的生態學概念[17]。功能多樣性簡化了群落物種研究的復雜性[17],也因其對群落生產力強大的預測潛能[10],被研究者廣泛用于多樣性與生產力關系的研究[6- 8]。但也有研究者發現功能多樣性與生產力無關[8,11]。對草地生態系統而言,放牧作為首要且不可避免的干擾方式[12],擾亂了多樣性與生產力之間的相互關系,使得物種和功能多樣性哪一個對生產力有較顯著影響更加不確定。最主要的原因是研究者在野外試驗中多采用草地植物的現存量來表示生產力水平[8-10,19],而未考慮草地普遍存在且不可避免的放牧強度造成的生物量差異。本研究連續4年對不同層次多樣性與生物量的關系進行研究表明,在不同的放牧強度區豐富度(物種、生活型和功能群)與生物量之間的關系均不顯著。而對于均勻度和優勢度而言,僅在高強度放牧區(Plot4、Plot5、Plot6)生活型均勻度隨生物量的增加而降低,生活型優勢度隨生物量的增加而增加。生活型作為植物適應生態環境而具有的一定形態外貌、結構和習性[23]。在放牧干擾下,植物群落的生活型調整決定群落能否適應放牧生境而維持生存[23]。本研究中,在高強度放牧區,生活型均勻度和優勢度對地上生物量表現出積極作用,更敏感的反映了生物量的變化。而豐富度對生物量的變異不敏感,原因可能是雖然物種豐富度在放牧強度的時間累積作用下發生分異,但豐富度(物種和物種組合類群的多少)的變化總是緩慢的。而均勻度和優勢度易受放牧干擾的直接影響,使得能夠適應高強度放牧干擾的優勢物種和優勢類群的比例增加。高強度放牧干擾激發了這一部分植物的生存潛力,因而高強度放牧區群落優勢物種和優勢類群的生存優勢就越明顯[12],即優勢度越高、均勻度越低,群落在放牧干擾下的生產力也就越高。高強度放牧區生產力的增加主要通過優勢物種或優勢類群的生存優勢來實現。同樣是物種組合類群,為什么是生活型多樣性與放牧干擾下的生物量有更強烈的密切關系,而不是功能群多樣性。原因可能是同樣作為物種組合類型的指標,功能群多樣性和生活型多樣性均可用于簡化群落物種研究的復雜性[17]。但功能群是基于生理、生活史或其它與某一生態過程相關以及與物種行為相聯系的一些生物學特性來劃分的,且干擾下群落的純生理性功能分化需在較長時期才能實現[27-28]。而群落生活型即植物的外部形態可以在較短時間內對干擾做出響應[23],進而影響群落結構,具有更強的操作性。生活型劃分是以植物趨同適應的形態表型為基礎[23],而功能群劃分以植物的生理生態特性的趨同性為特征[28]。放牧干擾下的生活型態的表型變化往往先于生理生態特性的變化,因而生活型多樣性在高強度放牧干擾下可以表現與生物量的顯著相關關系。生活型多樣性可作為放牧干擾下生物量變化快速預測的有效指標。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
趣味(語文)(2020年3期)2020-07-27 01:42:46
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
成才之路(2017年21期)2017-07-27 20:52:40
安徽農學通報(2017年9期)2017-05-19 23:03:05
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
現代農業科技(2016年22期)2017-03-24 12:21:57
學周刊·下旬刊(2015年10期)2015-07-22 12:36:17
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
