6個新疆地方梨品種S基因型鑒定
2018-08-01 07:53:20張校立艾沙江買買提徐葉挺王繼勛
江蘇農業科學 2018年13期
關鍵詞:新疆
張校立, 艾沙江·買買提, 徐葉挺, 許 娟, 鄧 莉, 王繼勛
(新疆農業科學院園藝作物研究所/農業部新疆地區果樹科學觀測試驗站,新疆烏魯木齊 830091)
新疆地方梨品種屬于新疆梨系統。新疆梨系統是梨五大栽培系統之一,代表品種是著名的庫爾勒香梨(PyrusbretschneideriRehd)。庫爾勒香梨維吾爾語名乃西米提或乃西普提,蒙古語名為開登木,原產新疆庫爾勒地區,在新疆已有1 400多年的栽培歷史,其果實香味濃郁、皮薄肉脆、清甜爽口、細嫩多汁、耐貯藏性強,是新疆名特優果品之一。截至2016年,庫爾勒香梨種植面積達7.39萬hm2,產量達128.04萬t[1],香梨產業已成為庫爾勒地區的主導產業。庫爾勒香梨自交不親和,S基因型為S22S28[2],自交結實率低,需要配置授粉品種,但不同的授粉品種會對香梨果實產生顯著的花粉直感效應,突出表現為不同花粉授粉后果實萼片脫落或宿存,萼片脫落后的幼果發育成為“母梨”,萼片宿存的幼果發育成為“公梨”,二者不但外在品質果形上差異很大,而且內在品質和口感上也差異明顯,市場價格差異非常顯著。徐勝利等研究表明,選擇新疆梨作為授粉品種能夠明顯改善香梨果形和果實品質[3]。鑒定新疆地方梨品種S-等位基因對庫爾勒香梨合理授粉品種的選配具有重要的理論意義。梨自交不親和性基因的研究始于20世紀50年代,國外最早研究的是Kikuchi、Machida等日本學者,采用廣泛的田間授粉雜交試驗,從日本梨中鑒定了7個S等位基因,命名為S1、S2、S3、S4、S5、S6、S7[4-6]。國內最早研究的是烏云塔娜,2003年,從中國白梨中分離鑒定了7個新的S基因,分別記為S17、S19、S20、S21、S22、S26、S27等位基因[7]。譚曉風等分別從白梨、沙梨及秋子梨等亞洲梨種群分離鑒定了34個S等位基因[8]。從2003年開始至今采用PCR-RFLP、DNA測序、雜交檢測等方法開展了中國梨自交不親和性的研究,發現了包括S1~S10基因在內的42個S基因,從中國梨中克隆了S12、S13、S15、S17~S22、S25~S30、S32、S34、S35、S38~S40、S42、S46等S基因全長cDNA序列,確定了近500個梨品種的S基因型。近500個品種中大部分都是白梨系統、秋子梨系統、沙梨系統的品種,新建梨系統的品種非常少,目前,只有陳慧等鑒定了黃句句(S22S34)、伊犁紅句句(S22S28)、早熟句句(S17S19)、乃希木特阿木提(S19S28)、沙01號(S22S28)、斯爾克甫梨(S22S28)、墨梨(S26Sb)、貴德長把(S19Sb)等8個新疆梨系統品種S-基因型[9]。新疆梨地方品種是我國特有的梨樹資源,新疆地方梨品種S基因型的鑒定研究比較少,大部分新疆地方梨品種的S基因型還未鑒定,我們研究的6個新疆地方梨品種的基因型目前尚未有人報道。我們利用DNA測序技術鑒定了6個新疆地方梨品種的S基因型。6個品種S基因型的鑒定,以期豐富我國梨品種S基因型信息;為庫爾勒香梨篩選最佳的授粉梨品種提供理論基礎,并可為庫爾勒香梨的豐產栽培技術提供科學依據,也為香梨品種的遺傳改良奠定基礎。
1 材料與方法
1.1 材料
供試品種為褐色句句梨、霍城冬黃梨、八月梨、庫爾勒黃酸梨、奎克阿木特2號、也歷克阿木特。品種均來自新疆農業科學院輪臺國家果樹資源圃。于2017年4月上旬采集各品種幼嫩葉片置于液氮中,帶回實驗室后于-80 ℃冷凍保存。
1.2 試劑
ExTaq、pMD18-T Vector、DNA回收試劑盒,購自北京全式金生物技術(TransGen Biotech)有限公司;DNA提取試劑盒,購于天根科技生化有限公司;其他藥品均購自上海生工生物工程技術服務有限公司。
1.3 基因組DNA提取
參照植物基因組DNA提取試劑盒說明書(柱式植物DNAout2.0,天根科技生化有限公司)提取梨基因組DNA,在1%瓊脂糖凝膠、1×TAE緩沖液、3~5 V/cm下電泳,電泳結果用上海復日FR-980凝膠圖成像系統照相,檢測DNA樣品。
1.4 PCR擴增及檢測
根據已報道的梨的S基因相關信息,獲知梨S基因的一級結構由5個保守區、1個高頻可變區及1個內含子組成。高頻可變區是花粉識別的部位。S基因多態性主要源于高頻可變區的差異,通常根據高頻可變區的差別判斷不同的S基因。因此,根據S基因一級結構特點,利用S基因高變區(hypervariable region,HV)的保守序列設計合成引物。
正向引物PF:5′-TTTACGCAGCAATATCAG-3′;
反向引物PR:5′-AC(A/G)TTCGGCCAAATAATT-3′。
在江南等擴增反應體系及反應過程[10]的基礎上略作調整,使用ExTaq酶保證擴增結果的可靠性,擴增產物在1.4%瓊脂糖凝膠上于100 V電泳30~45 min,上海復日FR-980凝膠圖像系統分析拍照并記錄分析。
1.5 PCR產物回收、克隆、測序
利用凝膠回收試劑盒回收目的DNA片段,與PEASY-T5載體16 ℃條件下過夜連接,連接產物轉化至感受態大腸桿菌Trans1-T1,涂于SOC/Amp50固體培養基上37 ℃過夜。挑選陽性克隆,菌液PCR篩選帶有目的片段長度的克隆。每個S基因片段挑選3個陽性克隆,送上海生工生物工程技術服務有限公司測序。測序結果利用Blast與GenBank中已知的S-RNase基因序列比較,確定其S基因型。
2 結果與分析
2.1 S基因特異性擴增
采用S等位基因特異性引物PF和PR對6個梨品種基因組DNA進行PCR擴增,瓊脂糖電泳檢測結果見圖1。
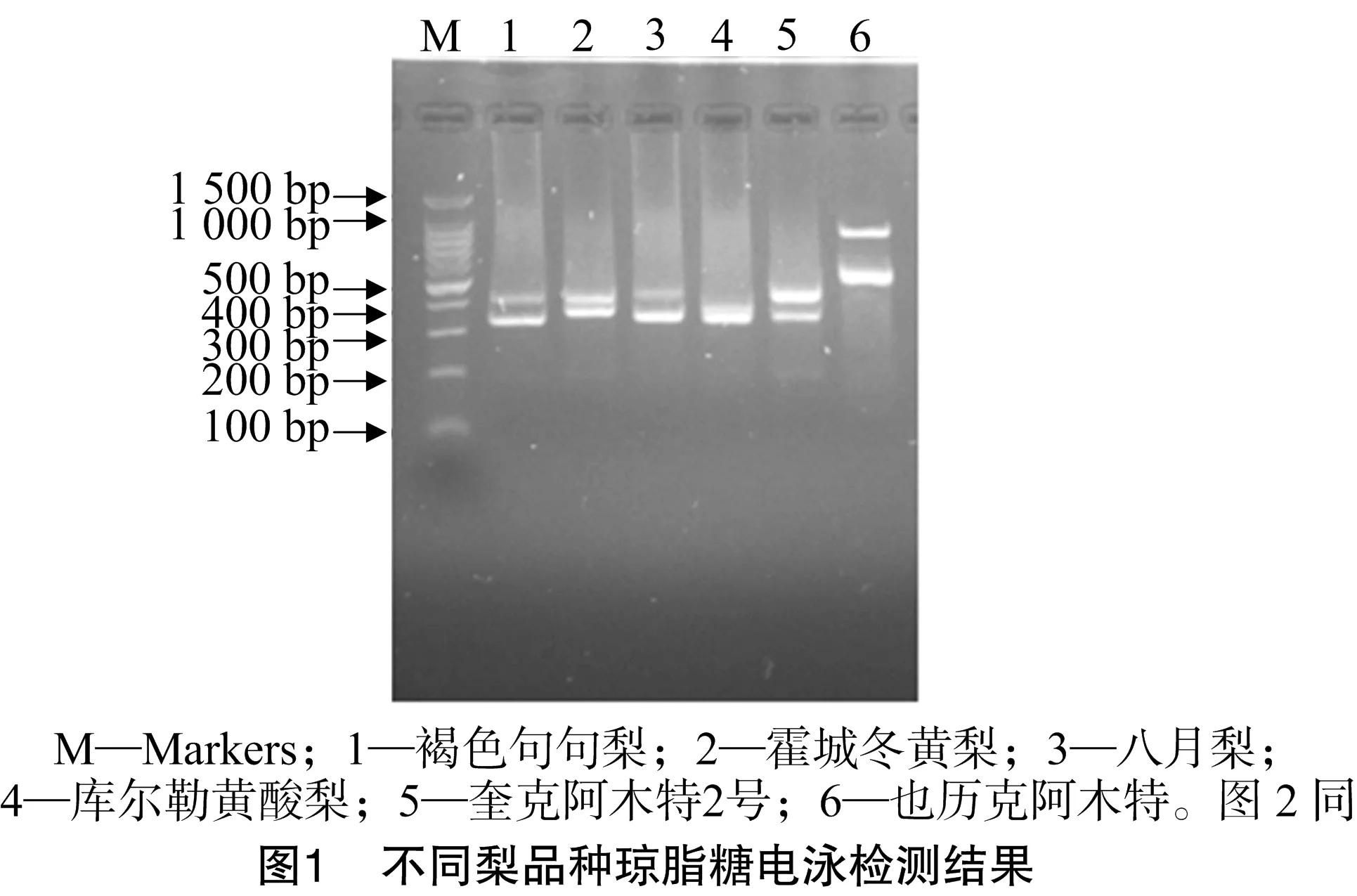
從圖1可以看出,每個梨品種擴增都得到2條S基因特異片段,大約在300~1 500 bp。褐色句句梨、霍城冬黃梨、八月梨、奎克阿木特2號、也歷克阿木特等5個品種都擴增出明顯的2條帶;庫爾勒黃酸梨擴增出來的2條帶不是太明顯,說明這個品種的S等位基因片段大小相差很小。
2.2 梨品種PCR擴增產物的聚丙烯酰胺凝膠電泳分離
將PCR擴增產物進一步進行聚丙烯酰胺凝膠電泳分離,PCR產物在聚丙烯酰胺凝膠上電泳隨遷移率的不同,表現為不同位置的條帶。從圖2可以看出,6個品種在聚丙烯酰胺凝膠上均呈現2條明顯的條帶,褐色句句梨、霍城冬黃梨、八月梨、奎克阿木特2號4個品種的2條帶都在300~500 bp之間;褐色句句梨、霍城冬黃梨的1條約450~500 bp 之間的擴增條帶在聚丙烯酰胺凝膠上的位置相同,它們的S基因的堿基序列是否相同有待進一步分析;也歷克阿木特的1條帶在1000~1 500 bp內,另1條帶在600~1 000 bp 內。
2.3 梨品種S基因型的鑒定
聚丙烯酰胺凝膠電泳后回收目的條帶,將克隆后的陽性克隆送上海生工生物工程技術服務有限公司測序,將12條測定的結果序列使用生物信息學軟件進行Blast搜索、同源性比較,根據分析GenBank內S基因的堿基序列及同源性,確定6個新疆地方梨品種的S基因型。從表1可以看出,6個品種中S基因的核苷酸序列與GenBank中已登錄的梨S基因有極高的同源相似性,褐色句句梨的S基因與S28、S4基因的相似性達98%、99%;霍城冬黃梨的S基因與S28、S4基因的相似性達100%、99%;八月梨S基因與S28、S4基因的相似性達99%、100%;庫爾勒黃酸梨的S基因與S22、S35基因的相似性達100%、100%;奎克阿木特2號的S基因與S4、S16基因的相似性達100%、100%;也歷克阿木特S基因與S18、S12基因的相似性達94%、100%。
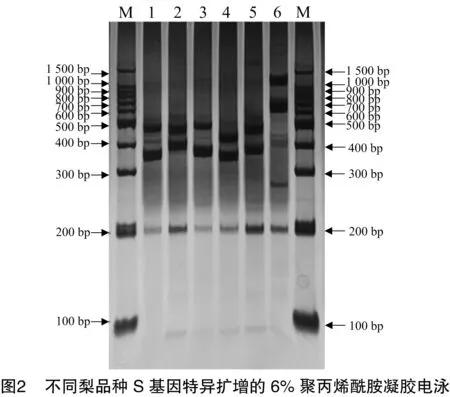
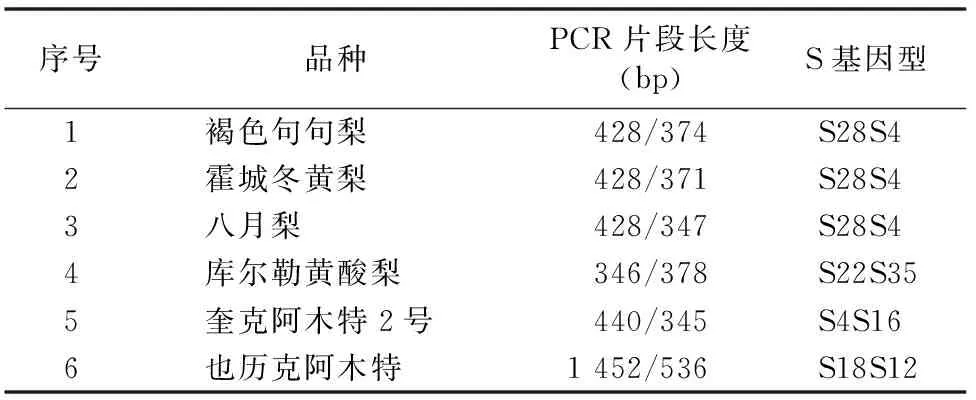
表1 6個新疆地方梨品種的S基因型
注:系統為新疆梨;引物為PF/FR。
3 討論與結論
本研究應用PCR技術和對S基因特異性擴增條帶的測序,通過DNA序列分析確定了6個新疆地方梨品種的S基因型。鑒定結果為:褐色句句梨S28S4、霍城冬黃梨S28S4、八月梨S28S4、庫爾勒黃酸梨S22S35、奎克阿木特2號S4S16、也歷克阿木特S18S12。同為新疆地方梨品種,它們之間既有相同之處,也有不同之處,如褐色句句梨、霍城冬黃梨、八月梨等3個品種具有相同的基因型S28S4,這3個品種間不能相互作授粉品種,因為它們具有相同的基因型,如果相互間作授粉品種,就會出現不結果或結果率極低;庫爾勒黃酸梨、奎克阿木特2號、也歷克阿木特3個品種的基因型不相同,這3個品種間可以相互作授粉品種。褐色句句梨、霍城冬黃梨、八月梨3個品種具有相同的基因型,并且都具有庫爾勒香梨的S28,庫爾勒黃酸梨具有庫爾勒香梨的S22基因,表明褐色句句梨、霍城冬黃梨、八月梨、庫爾勒黃酸梨與庫爾勒香梨在起源上相關性比較大,奎克阿木特2號、也歷克阿木特與庫爾勒香梨在起源上相關性比較小。鑒定出的6個新疆地方梨品種分別含有S4、S12、S16、S18、S22、S28、S35等S基因,這些基因分別屬于白梨系統、沙梨系統和西洋梨系統的S基因,初步推論新疆地方梨品種可能是白梨系統、沙梨系統和西洋梨系統的雜種,雜種后代又經過長期的自然雜交,互相演化而來成現在的品種。在果樹育種學上,沈德緒等認為,新疆梨可能是西洋梨與白梨的雜交種[11]。新疆地方梨品種具體的起源與分類還需對其更多品種的S-基因進行鑒定,并需要進一步的分析與研究。
6個新疆地方梨品種基因型的鑒定,增加了我國6個新疆梨品種的S基因型信息,可為庫爾勒香梨授粉品種的篩選新疆地方梨品種提供理論依據。褐色句句梨、霍城冬黃梨、八月梨3個品種具有庫爾勒香梨的S28基因,庫爾勒黃酸梨具有庫爾勒香梨的S22基因,表明褐色句句梨、霍城冬黃梨、八月梨和庫爾勒黃酸梨不太適宜作庫爾勒香梨的授粉品種;奎克阿木特2號與也歷克阿木特可以作為庫爾勒香梨的授粉品種,但其授粉后對庫爾勒香梨品質的影響,還需開展進一步研究。
猜你喜歡
國畫家(2022年2期)2022-04-13 09:07:46
四川文學(2021年4期)2021-07-22 07:11:54
新作文·高中版(2021年4期)2021-07-14 11:55:06
環球人物(2021年8期)2021-05-06 08:00:43
音樂天地(音樂創作版)(2020年12期)2021-01-11 01:13:40
音樂天地(音樂創作版)(2020年7期)2020-09-11 06:17:18
華人時刊(2018年15期)2018-11-10 03:25:32
人民中國(日文版)(2015年10期)2015-04-16 03:53:48
絲綢之路(2014年9期)2015-01-22 04:24:46
新疆人文地理(2009年7期)2009-09-29 09:56:14
