生育酚及其衍生物保護自由基誘導的生物大分子損傷和抑制HepG2細胞增殖的作用
2018-07-23 08:22:16韓靜靜田丹丹高玉星肖春霞
食品科學 2018年13期
關鍵詞:生物
韓靜靜,田丹丹,高玉星,肖春霞
(西北農林科技大學食品科學與工程學院,陜西 楊凌 712100)
VE又名生育酚,是生物體內的天然抗氧化劑,廣泛分布于谷物、種子、果蔬和動物產品中,具有清除自由基[1]、維持哺乳動物及禽類生育[2-3]、調節免疫[4]、抑制血小板凝集和黏附[5]等功能。但VE不溶于水,遇堿不穩定,對氧氣、光照敏感,易被氧化,并且某些金屬離子如Fe2+可加速其氧化[6]。此外,Reboul等[7]發現VE的生物利用率不穩定,容易受到食物基質、食品加工過程等的影響,因此限制了其在食品、藥品和化妝品等領域中的應用。隨著研究的深入,一系列生育酚衍生物如生育酚醋酸酯(tocopheryl acetate,TA)、生育酚煙酸酯(tocopheryl nicotinate,TN)、生育酚琥珀酸酯(α-tocopheryl succinate,α-TOS)等被陸續報道。這些衍生物均是VE的游離羥基與醋酸、煙酸、琥珀酸的羧基酯化形成的酯類衍生物,由于VE的活潑羥基被保護,這些衍生物具有更好的穩定性,而親水性羧基基團則增加了其水溶性。此外,這些衍生物還表現出良好的生物活性,這可能是由于新引入的酸酐結構導致了VE功能的變化[8]。Badraoui等[9]通過體內、體外實驗證明,TA能抑制氧化應激損傷,并能誘導乳腺癌細胞凋亡。Schlieper等[10]發現TN具有改善心律不齊的功效。Prasad等[11]研究發現,與TA、TN相比,α-TOS對黑色素瘤細胞生長分化的抑制作用更明顯。此外,α-TOS對結腸癌[12]、乳腺癌[13]、胃癌[14]等多種惡性腫瘤細胞的增殖有良好的抑制作用。Rego等[15]比較了TN和α-TOS對視網膜細胞氧化的抑制作用,結果表明后者作用更為明顯。大量文獻表明,在眾多生育酚衍生物中α-TOS的生物活性較為突出[11-15]。近年來,一種新的生育酚衍生物——生育酚聚乙二醇1000琥珀酸酯(tocopheryl polyethylene glycol 1000 succinate,TPGS)的綠色合成研究備受關注[16],TPGS是α-TOS的游離羧基與大量聚乙二醇酯化而成,親水性聚乙二醇極大地改善了TPGS的水溶性,由于其兩親性質,可作為增溶劑、乳化劑、穩定劑等使用[17-18]。Thi等[19]以TPGS為載體制備了不溶性藥物的傳遞體系,可延長藥物在血液及組織中的循環時間。Nguyen等[20]報道TPGS包裹紫杉醇可增加其體外攝取率,使其更容易進入固態瘤中心。Zhu Xianbing等[21]發現槲皮素經TPGS包裹,增加了其水溶性及抗紫外線損傷能力,并提高了其在皮膚表皮層和真皮層的存留量。TPGS已被美國食品藥品監督管理局批準為安全的藥用輔料[22],但關于TPGS的生物活性鮮有報道。
本研究對比了VE、α-TOS、TPGS對自由基誘導的生物大分子氧化損傷的保護作用,以及對人肝癌細胞(HepG2細胞)增殖的抑制作用,旨在比較研究VE不同衍生物之間生物活性的差異,探討其內在機理,為VE及其衍生物在食品中的廣泛應用提供一定的理論依據。
1 材料與方法
1.1 材料與試劑
VE、α-TOS、TPGS(結構式見圖1) 成都艾科化學技術有限公司;牛血清白蛋白(bovine serum albumin,BSA) 上海澤龍生物工程有限公司;2,2’-偶氮二異丁基脒二鹽酸鹽(2,2’-azobis-2-methylpropanimidamide,dihydrochloride,AAPH)、鯡魚精DNA(herring sperm DNA,hsDNA)、亞油酸(linoleic acid,LA)、偶氮二異庚腈(2,2’-azobis(2,4-dimethyl)valeronitrile,AMVN)、3-(4,5-二甲基噻唑-2)-2,5-二苯基四氮唑溴鹽(3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide,MTT) 美國Sigma公司;2,4-二硝基苯肼(2,4-dinitrophenyl hydrazine,DNPH)、二甲基亞砜(dimethyl sulfoxide,DMSO) 美國Amresco公司;三氯乙酸 天津市博迪化工有限公司;硫代巴比妥酸(thiobarbituric acid,TBA) 國藥集團藥業股份有限公司;考馬斯亮藍 北京鼎國昌盛生物技術有限責任公司;五水硫酸銅、30%過氧化氫、七水合硫酸亞鐵、磷酸二氫鈉、磷酸氫二鈉、無水乙醇、甲醇、冰乙酸均為分析純。
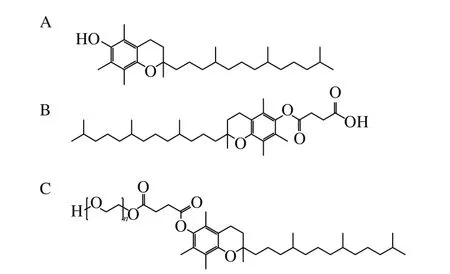
圖1 VE(A)、α-TOS(B)和TPGS(C)的結構式Fig. 1 Structures of VE (A), α-TOS (B) and TPGS (C)
HepG2細胞購自上海生命科學研究院,用含有10%胎牛血清、100 U/mL青霉素和100 μg/mL鏈霉素的RPMI 1640培養基在37 ℃、5% CO2條件下培養。
1.2 儀器與設備
電泳儀(電泳供電裝置)、凝膠成像系統 美國伯樂公司;UV-2450型紫外-可見分光光度計 日本Shimadzu公司;BP211D型精密電子天平 德國Startorius公司;HH-2型恒溫水浴鍋 江蘇省金壇市環宇科學儀器廠;Max M2型多功能微孔板檢測儀 美國Molecular公司。
1.3 方法
1.3.1 Cu2+/H2O2誘導BSA氧化、羰基化的測定
在BSA終質量濃度為0.8 mg/mL的反應體系中,先加入不同濃度的VE、α-TOS和TPGS使其終濃度分別為10、50、100、500、1 000 μmol/L(均用甲醇溶解),空白組以等體積的磷酸鹽緩沖液代替,模型組以等體積的甲醇代替,漩渦振蕩混勻,置于37 ℃水浴中預處理10 min。然后加入Cu2+/H2O2反應體系(空白組不加),并將所有反應組用磷酸鹽緩沖液定容至500 μL,使Cu2+和H2O2的終濃度分別為100 μmol/L和2.5 mmol/L,37 ℃水浴反應60 min。反應結束后,采用考馬斯亮藍R-250染色法檢測BSA氧化損傷程度,采用DNPH比色法檢測BSA羰基化程度[23]。
1.3.2 AAPH誘導BSA氧化、羰基化的測定
蛋白模型和VE、α-TOS、TPGS處理方法同1.3.1節,預處理結束后加入AAPH(空白組不加),使其終濃度為50 mmol/L,置于37 ℃水浴反應5 h。反應結束后,采用考馬斯亮藍R-250染色法檢測BSA氧化損傷程度,采用DNPH比色法檢測BSA羰基化程度[23]。
1.3.3 自由基誘導LA過氧化的測定
采用Fe2+/VC反應體系產生的羥自由基(·OH)[24]、AMVN熱分解產生的烷氧自由基(ROO·)[25]誘導LA發生脂質過氧化反應。
在LA(甲醇助溶)終濃度為1 mmol/L的反應體系中,先分別加入不同濃度的VE、α-TOS和TPGS使其終濃度分別為10、50、100、500、1 000 μmol/L(均用甲醇溶解),空白組以等體積的磷酸鹽緩沖液代替,模型組以等體積的甲醇代替,混勻后加入Fe2+/VC或AMVN(空白組不加),將所有反應組用磷酸鹽緩沖液定容至1 mL,使反應體系中Fe2+、VC、AMVN的終濃度分別為50 μmol/L和1、10 mmol/L。Fe2+/VC體系37 ℃水浴避光反應24 h,AMVN體系反應12 h。反應結束后,以硫代巴比妥酸反應產物(thiobarbituric acid reactive substances,TBARS)濃度表示LA脂質過氧化水平[26]。
1.3.4 AAPH誘導DNA氧化損傷的測定
在hsDNA終質量濃度為2 mg/mL的反應體系中,先加入不同濃度的VE、α-TOS和TPGS使其終濃度分別為10、50、100、500、1 000 μmol/L(均用甲醇溶解),空白組以等體積的磷酸鹽緩沖液代替,模型組以等體積的甲醇代替,再加入AAPH溶液(空白組不加),并用磷酸鹽緩沖液將所有反應組定容至1 mL,使AAPH的終濃度為40 mmol/L,混勻后37 ℃水浴反應12 h。反應結束后采用TBA法檢測DNA氧化損傷程度[27]。
1.3.5 MTT法測定細胞活力
將HepG2細胞接種于96 孔板中,置于37 ℃、5% CO2培養箱培養過夜,使細胞貼壁。分別加入不同濃度(10、20、30、40、50 μmol/L)的VE、α-TOS和TPGS,空白組不加。每組設置8 個平行,置于37 ℃、5% CO2培養箱中繼續培養24 h。棄上清液,每孔加入100 μL終質量濃度為0.5 mg/mL的MTT溶液,置于培養箱中繼續培養,4 h后棄去培養液,每孔加入100 μL DMSO,置于搖床上低速振蕩10 min,使結晶物充分溶解。使用多功能微孔板檢測儀測定各孔在570 nm波長處的OD值,并根據下式計算細胞存活率。

1.4 數據分析
2 結果與分析
2.1 VE、α-TOS和TPGS對自由基誘導蛋白質損傷的影響
2.1.1 VE、α-TOS和TPGS對Cu2+/H2O2誘導BSA氧化降解的影響
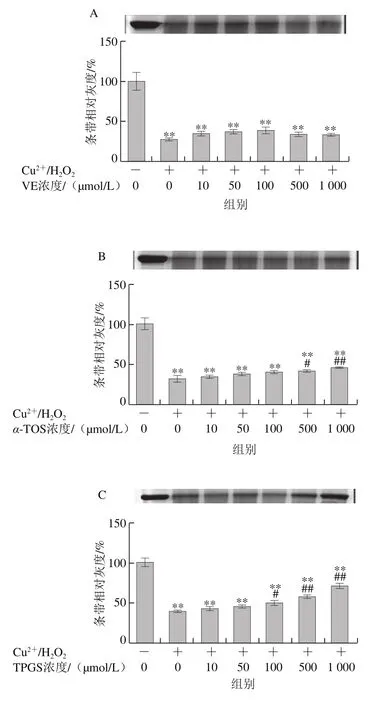
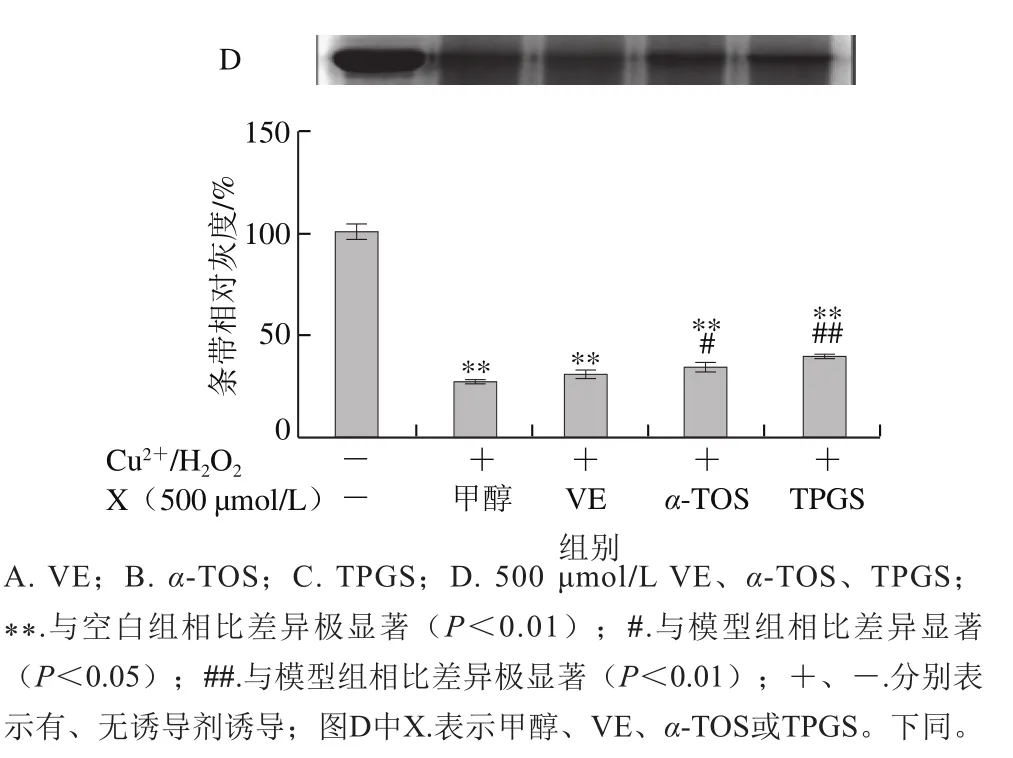
圖2 VE、α-TOS和TPGS對Cu2+/H2O2誘導BSA氧化降解的影響Fig. 2 Effects of VE, α-TOS and TPGS on Cu2+/H2O2-induced BSA oxidative degradation
Fe2+、Fe3+、Cu2+、Mn2+、Ni2+等金屬離子可以催化H2O2分解,產生氧化活性更高的·OH,·OH能與所有氨基酸反應[28],因此本實驗采用Cu2+/H2O2反應體系誘導BSA氧化損傷,探討VE、α-TOS和TPGS對自由基誘導BSA氧化降解的抑制作用。從圖2A可以看出,VE對Cu2+/H2O2誘導的BSA氧化降解沒有明顯規律性。而α-TOS、TPGS均濃度依賴性地抑制了BSA的氧化降解,在500、1 000 μmol/L濃度下,α-TOS和TPGS的抑制作用顯著。由圖2D可知,相同濃度下(500 μmol/L),與VE、α-TOS相比,TPGS對Cu2+/H2O2誘導的BSA氧化降解的抑制作用最明顯,與模型組相比差異極顯著(P<0.01)。
2.1.2 VE、α-TOS和TPGS對AAPH誘導BSA氧化降解的影響
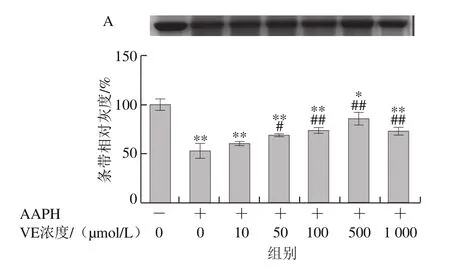
AAPH是一種水溶性的偶氮類化合物,通過均裂產生ROO·,在37 ℃和中性pH值條件下能穩定地產生自由基[29]。如圖3所示,VE、α-TOS和TPGS均能顯著抑制AAPH誘導的BSA氧化降解,在10~500 μmol/L濃度范圍內呈濃度依賴性,并且在500 μmol/L時最強,在1 000 μmol/L時有所減弱,但與模型組相比仍有極顯著差異(P<0.01)。圖3D表明,相同濃度(500 μmol/L)處理時,TPGS的抑制作用最明顯,且與空白組相比沒有顯著差異,即500 μmol/L的TPGS很好地抑制了AAPH誘導的BSA氧化降解。
2.1.3 VE、α-TOS和TPGS對Cu2+/H2O2誘導BSA羰基化的影響
如圖4所示,BSA本身羰基化程度很低,在Cu2+/H2O2作用下發生明顯的羰基化修飾。加入VE、α-TOS和TPGS后,BSA羰基化程度明顯受到抑制,且抑制作用隨濃度的升高而增強。由圖4D可知,相同濃度(500 μmol/L)的VE、α-TOS和TPGS均能顯著抑制BSA羰基的產生,但TPGS抑制作用最明顯。
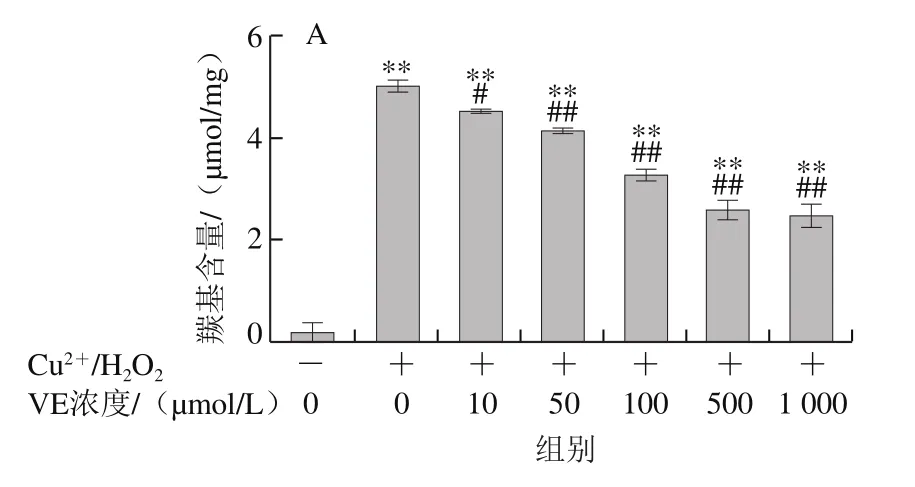
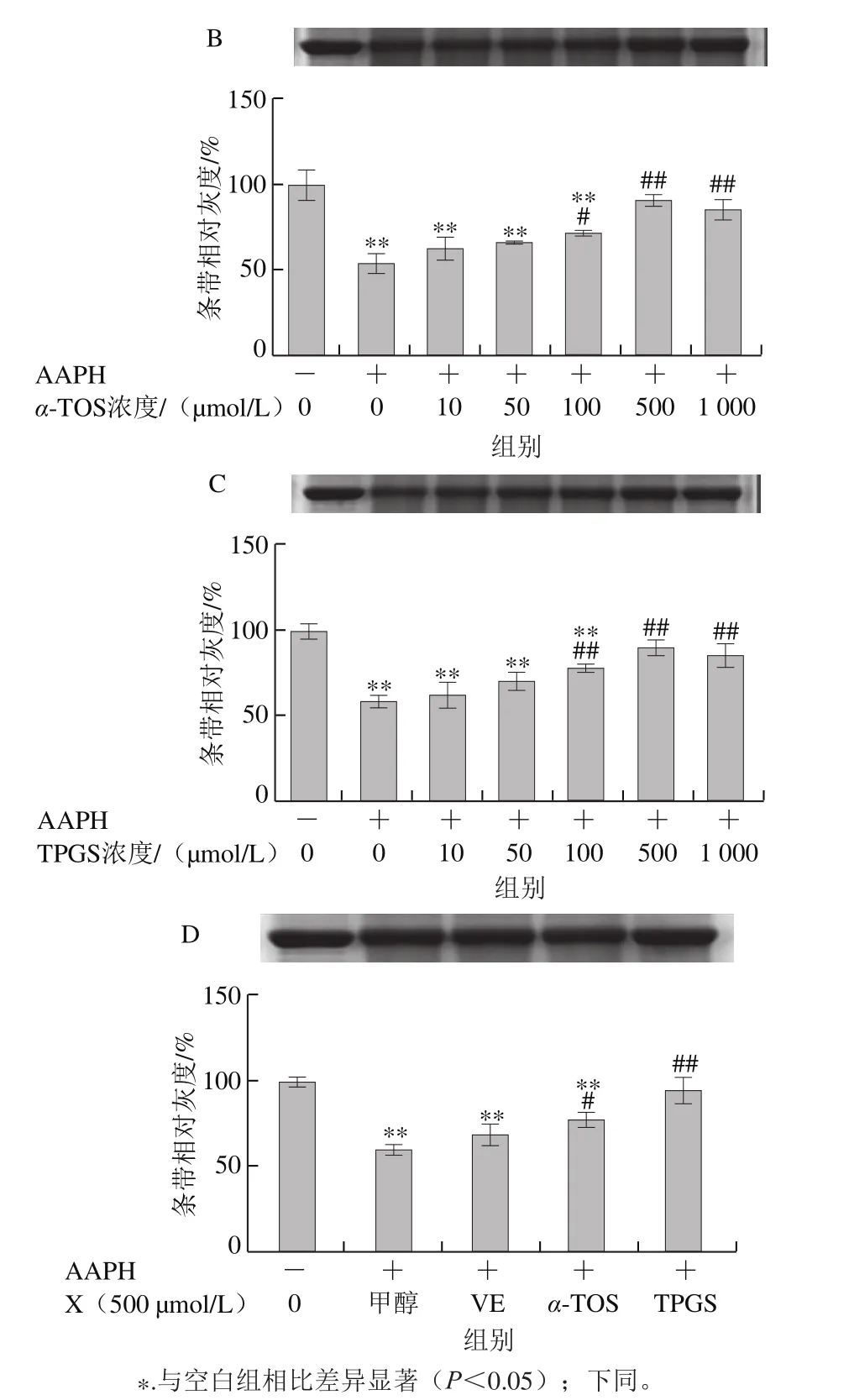
圖3 VE、α-TOS和TPGS對AAPH誘導BSA氧化降解的影響Fig. 3 Effects of VE, α-TOS and TPGS on AAPH -induced BSA oxidative degradation
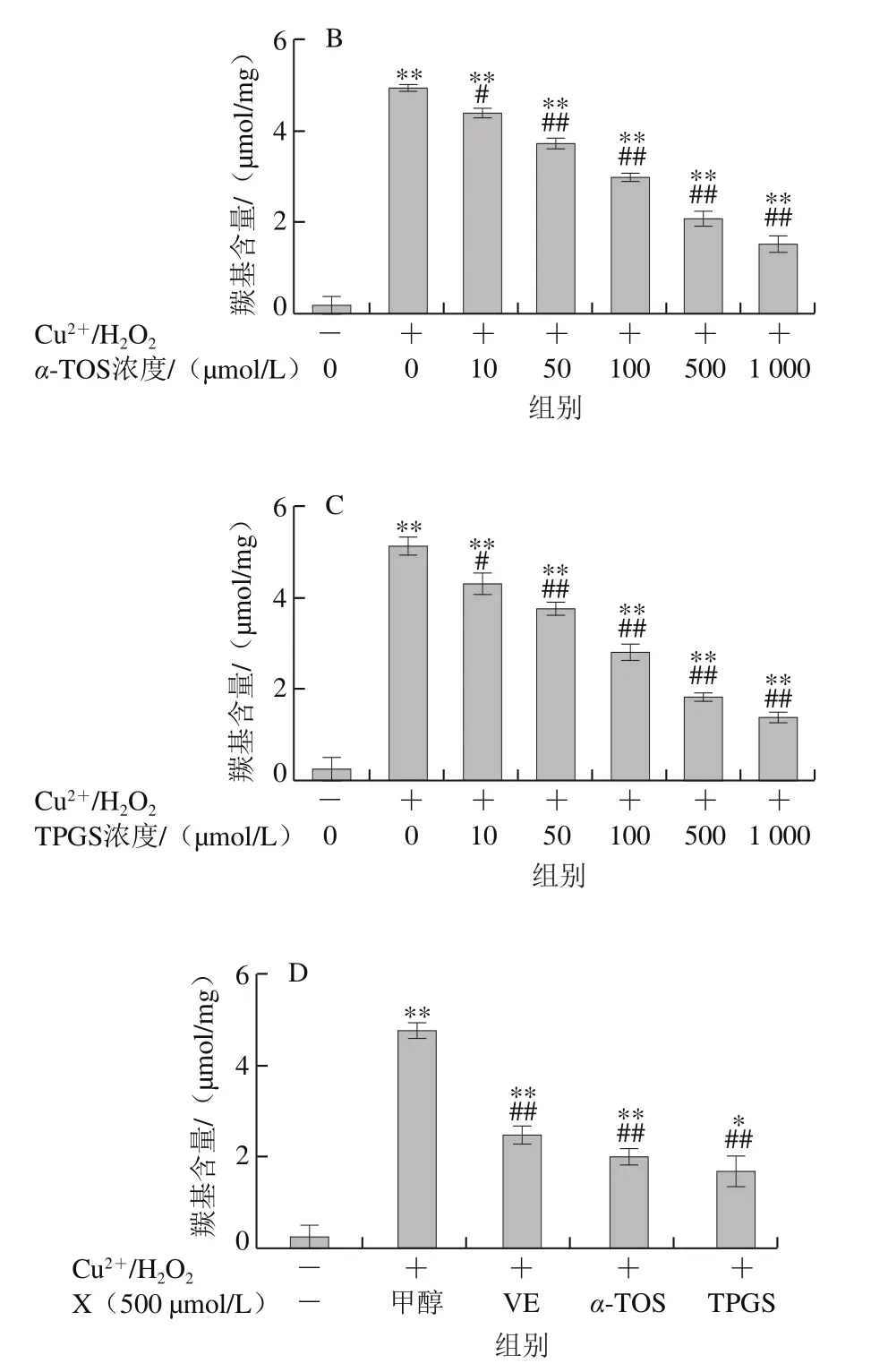
圖4 VE、α-TOS和TPGS對Cu2+/H2O2誘導BSA羰基化的影響Fig. 4 Effect of VE, α-TOS and TPGS on Cu2+/H2O2-induced carbonylation of BSA
2.1.4 VE、α-TOS和TPGS對AAPH誘導BSA羰基化的影響

圖5 VE、α-TOS和TPGS對AAPH誘導BSA羰基化的影響Fig. 5 Effects of VE, α-TOS and TPGS on AAPH-induced carbonylation of BSA
由圖5可知,AAPH誘導BSA發生明顯的羰基化修飾。VE、α-TOS和TPGS均濃度依賴性地抑制BSA羰基化修飾。如圖5D所示,500 μmol/L的VE、α-TOS和TPGS均顯著抑制了BSA羰基的產生,其中TPGS對AAPH誘導的BSA羰基化損傷的抑制作用最明顯。
2.2 VE、α-TOS和TPGS對自由基誘導LA過氧化的影響
2.2.1 VE、α-TOS和TPGS對Fe2+/VC誘導LA過氧化的影響
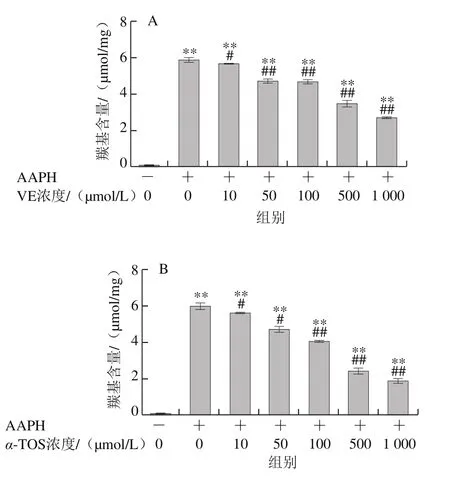
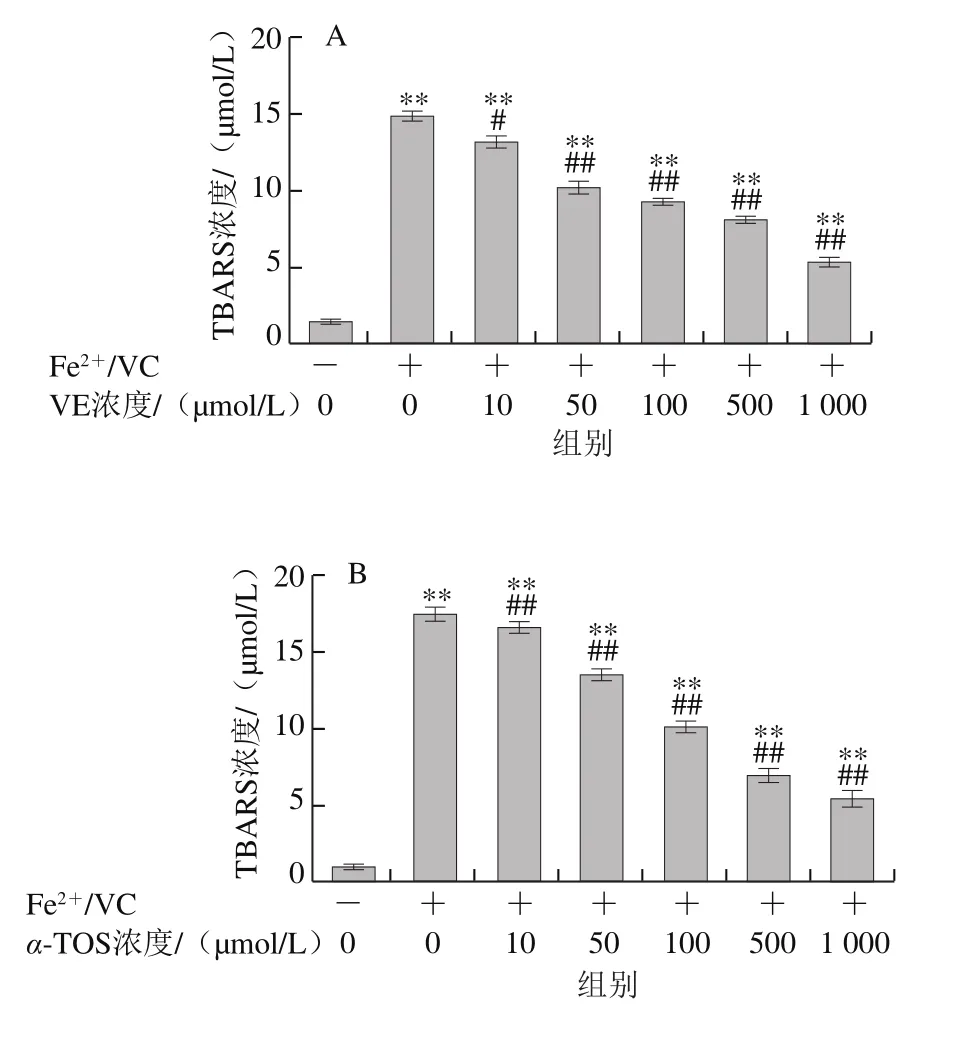
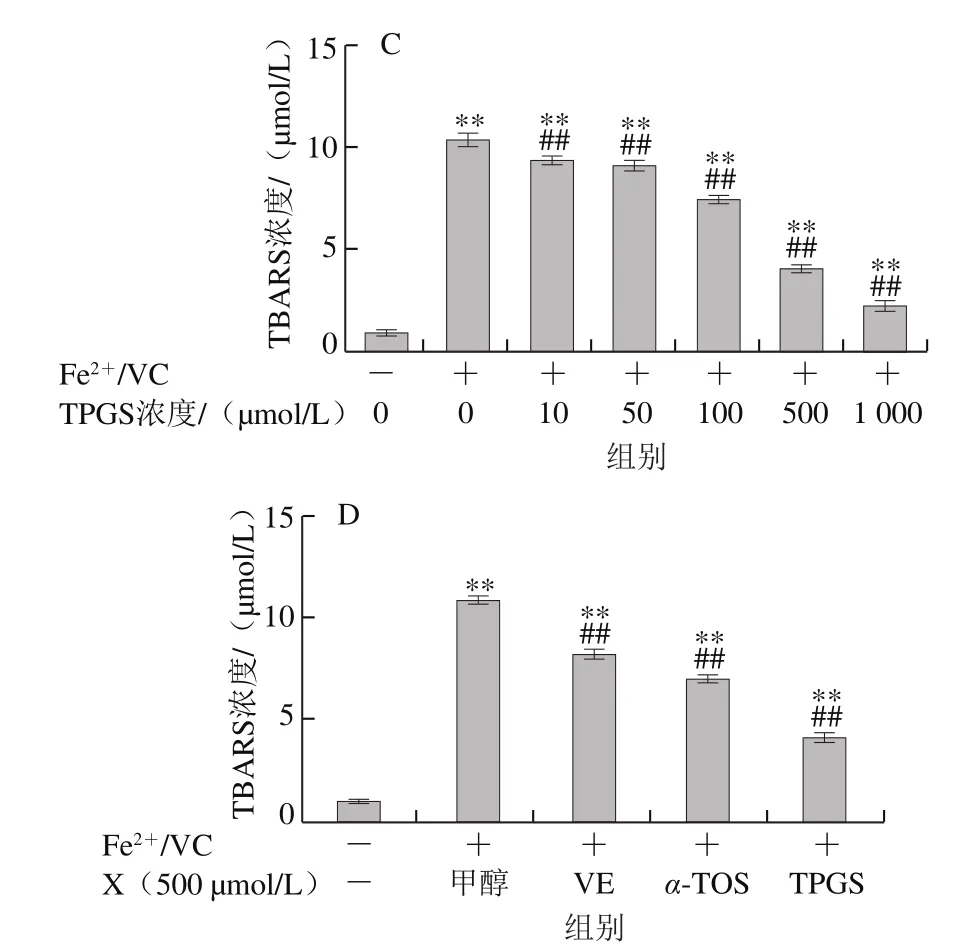
圖6 VE、α-TOS和TPGS對Fe2+/VC誘導LA過氧化的影響Fig. 6 Effects of VE, α-TOS and TPGS on LA peroxidation induced by Fe2+/VC
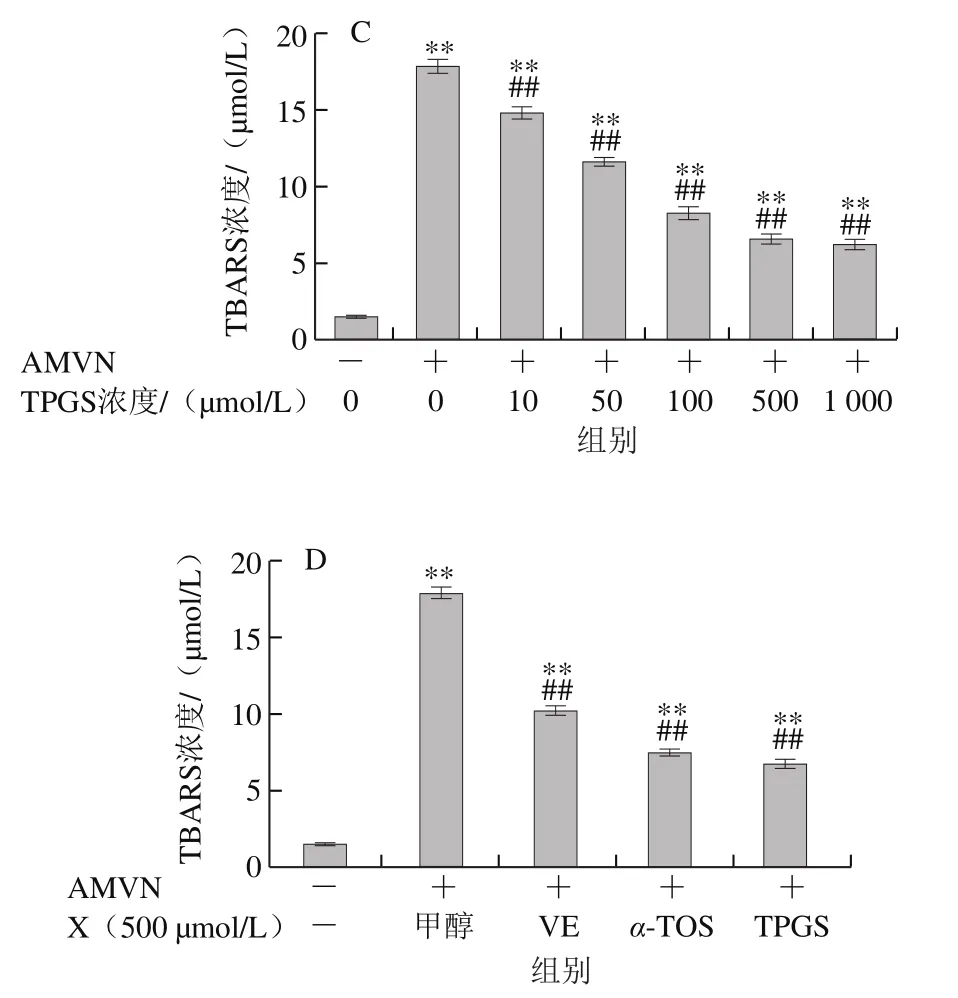
圖7 VE、α-TOS和TPGS對AMVN誘導LA過氧化的影響Fig. 7 Effects of VE, α-TOS and TPGS on LA peroxidation induced by AMVN
LA在脂質過氧化過程中會形成亞油酸氫過氧化物、4-羥基壬烯醛、丙二醛等氧化產物,這些產物能與TBA發生反應生成TBARS,其含量是衡量脂質過氧化程度的重要指標之一[30]。由圖6可知,LA在自然條件下自氧化反應很慢,生成TBARS濃度較低,Fe2+/VC反應體系誘導LA發生過氧化反應,VE、α-TOS和TPGS均顯著抑制了LA過氧化,且其抑制作用隨濃度的升高而增強。圖6D表明,相同濃度(500 μmol/L)下,TPGS對LA過氧化的抑制作用最強。
2.2.2 VE、α-TOS和TPGS對AMVN誘導LA過氧化的影響

AMVN是一種脂溶性偶氮類化合物,通過熱分解產生ROO·,常作為自由基引發劑用于脂質過氧化反應模型的建立[31]。如圖7所示,LA自然條件下生成的TBARS濃度很低,AMVN誘導LA發生過氧化反應,TBARS濃度極顯著增加(P<0.01)。圖7A~C表明,VE、α-TOS和TPGS均濃度依賴性地抑制了LA過氧化。圖7D表明,相同濃度(500 μmol/L)的VE、α-TOS和TPGS中,TPGS對AMVN誘導的LA過氧化的抑制作用最強。
2.3 VE、α-TOS和TPGS對AAPH誘導DNA氧化損傷的影響

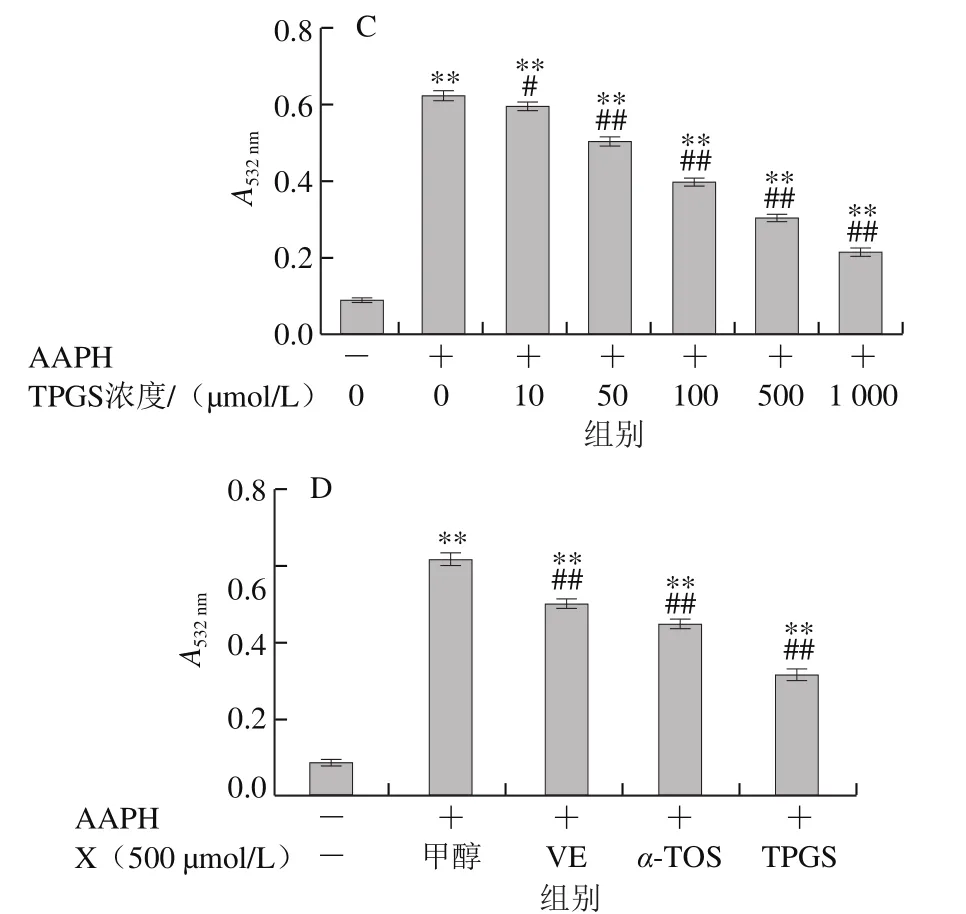
圖8 VE、α-TOS和TPGS對AAPH誘導DNA氧化損傷的影響Fig. 8 Effects of VE, α-TOS and TPGS on AAPH-initiated DNA oxidation
AAPH熱分解生成的ROO·使DNA分子的雙螺旋結構解旋,并最終生成含有羰基的20余種小分子化合物,DNA形成的裂解產物在酸性條件下與TBA反應生成TBARS[27],本實驗通過檢測TBARS含量來研究不同濃度的VE、α-TOS和TPGS對ROO·誘導DNA氧化損傷的抑制作用。由圖8可知,hsDNA在AAPH誘導下發生氧化損傷。隨著加入VE、α-TOS和TPGS濃度的增加,其對DNA氧化損傷的抑制作用增強。從圖8D可以看出,相同濃度(500 μmol/L)下,TPGS對AAPH誘導的DNA氧化損傷的抑制作用最強。
2.4 VE、α-TOS和TPGS對HepG2細胞活力的影響
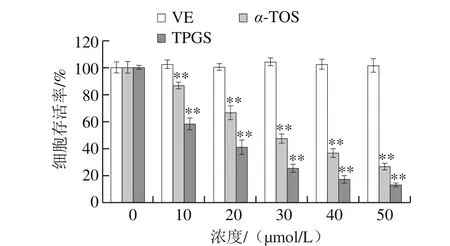
圖9 VE、α-TOS和TPGS對HepG2細胞活力的影響Fig. 9 Effects of VE, α-TOS and TPGS on HepG2 cell viability
活細胞能將外源性的MTT還原為水不溶性的藍紫色結晶甲瓚,并沉積在細胞中,經DMSO溶解并測定其OD值可間接反映活細胞數量。MTT法是測定細胞活力最常見、直接的方法[32]。如圖9所示,以空白組細胞存活率為100%,VE對HepG2細胞存活率沒有明顯影響;而隨著α-TOS、TPGS濃度的增加,HepG2細胞存活率逐漸下降,與空白組相比差異極顯著(P<0.01),其中TPGS對HepG2細胞增殖抑制作用最強。
3 討 論
VE常作為油脂加工中的抗氧化劑應用在肉類腌制中防止亞硝胺的生成,此外還具有多種生理功能。Azzi等[33]報道VE能改善與氧化應激相關的心血管疾病、癌癥、神經退行性疾病、白內障等疾病。Bo?kovi?等[34]發現補充一定量的VE可改善精神分裂癥患者的運動阻滯狀況。但VE在氧氣存在條件下不穩定,α-TOS和TPGS通過酯化修飾提高了化學穩定性,其中α-TOS能顯著抑制巨噬細胞NO的生成[35],并降低環孢菌素A對大鼠肝細胞的毒性[36]。TPGS由于具有良好的水溶性,被廣泛應用于難溶藥物的傳遞體系中,以提高藥物的滲透性和吸收效率[18,37-38],但尚鮮有針對VE和其水溶性衍生物α-TOS、TPGS生物活性差異的報道。
自由基具有未配對電子,化學性質非常活潑。在正常條件下,自由基參與細胞內的信號轉導、免疫防御、增殖、凋亡等多種生理活動[39]。當體內氧化和抗氧化體系平衡被打破時,過量的自由基就會攻擊體內的蛋白質、脂質、DNA等生物大分子,最終造成機體損傷。活性氧簇(reactive oxygen species,ROS)是自由基的一種,在應激狀態下可攻擊蛋白質、脂質、DNA等,進而誘發多種疾病,例如腫瘤[40]、神經退行性疾病[41]等。研究表明,ROS不僅參與腫瘤的產生,還與腫瘤的轉移密切相關[42]。
本實驗以Cu2+/H2O2、AAPH、AMVN等體系誘導產生自由基,對比研究VE、α-TOS、TPGS對自由基引起的生物大分子體外損傷的保護作用,結果表明,與VE、α-TOS相比,TPGS對生物大分子(蛋白質、脂質、DNA)損傷的保護作用最強。許多學者曾認為α-TOS發揮抗腫瘤功效是由于α-TOS水解釋放的VE發揮的作用,但多項研究表明,α-TOS是以整體的分子形式發揮作用,并不依賴于VE[43-44]。本研究結果顯示,TPGS對自由基誘導的生物大分子損傷的保護作用優于VE,推測TPGS可能也是以整體分子形式發揮抗氧化作用的,而不依賴于其水解產生的VE。TPGS結構中的琥珀酸酯保護了化學性質活潑的酚羥基,提高其穩定性,而高度聚合的聚乙二醇結構保護琥珀酸的另一個游離羧基,隨著聚乙二醇鏈段長度的增加,親水性也隨之增加。TPGS由于具有兩親性,在磷酸鹽緩沖液中,其疏水端與生物大分子(蛋白質、脂質、DNA)相互作用,吸附在生物大分子表面,而聚乙二醇與水具有較強的親和力,在磷酸鹽緩沖液中充分伸展,為自由基攻擊生物大分子提供了較大的空間位阻,從而達到對生物大分子的最佳保護效果。在HepG2細胞模型中,VE對HepG2細胞的存活率沒有明顯影響,而α-TOS和TPGS均能夠顯著降低HepG2細胞存活率,其中TPGS的抑制作用更為明顯。這可能是由于TPGS親水性強,與生物膜磷脂成分的相互作用弱,透過生物膜時的阻力減小、效率增加,在HepG2細胞內有效濃度大大增加。P-糖蛋白是一種由基因編碼的跨膜蛋白,在小腸、肝、腎等組織中分布廣泛,能夠外排大量結構和功能各異的外源性分子,Dintaman等[45]的研究表明TPGS是一種P-糖蛋白的抑制劑,可以抑制P-糖蛋白的外排作用,由此推測TPGS可能通過抑制HepG2細胞膜P-糖蛋白的外排作用,增加其在細胞內的濃度,從而更有效地抑制HepG2細胞增殖。
4 結 論
本實驗采用多種自由基體系對比研究VE及其衍生物α-TOS、TPGS對自由基引起的生物大分子體外損傷的保護作用,并從抑制HepG2細胞增殖方面比較其生物活性的差異,發現TPGS的抑制作用最顯著。這可能是由于TPGS引入了親水性強的聚乙二醇結構,在磷酸鹽緩沖液中充分伸展,為自由基攻擊生物大分子提供了較大的空間位阻;并且TPGS親水性強,更容易透過生物膜,減少生物膜的外排作用,提高在細胞中的有效濃度。作為一種安全的食品藥品傳遞載體,相較于VE和傳統的VE衍生物,TPGS具有更好的水溶性和穩定性。本研究結果將對VE及其衍生物的廣泛應用提供一定的理論依據。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
