兩種基因型菊芋幼苗對鹽脅迫的生長及生理響應
2018-07-04 08:02:10段海霞鄒恭魯楊永鑫呂世奇趙長明
干旱地區(qū)農(nóng)業(yè)研究 2018年3期
關鍵詞:植物
段海霞, 鄒恭魯, 楊永鑫, 李 穎, 呂世奇, 孫 杉, 趙長明
(蘭州大學生命科學學院,草地農(nóng)業(yè)生態(tài)系統(tǒng)國家重點實驗室,甘肅 蘭州 730000)
土壤鹽漬化是影響農(nóng)業(yè)生產(chǎn)以及生態(tài)環(huán)境的一個全球性問題,在全世界干旱和半干旱地區(qū),土壤鹽漬化限制作物生產(chǎn)[1,2]。據(jù)統(tǒng)計,我國有1億多公頃各種鹽漬土地,主要集中于西部六省區(qū),其面積約占全國鹽漬土面積的66.6%,西部鹽漬化耕地約344.7 × 104hm2,占西部地區(qū)耕地面積13.93%,占全國鹽漬化耕地37.4%,其中以典型鹽土面積最大且土壤鹽分以氯化物和硫酸鹽為主要成分[3,4]。鹽脅迫是農(nóng)業(yè)生產(chǎn)中影響植物生產(chǎn)力最主要的非生物脅迫之一,鹽脅迫下植物吸收水分和營養(yǎng)物質(zhì)能力降低,且生產(chǎn)速率迅速降低[5],也能引起一些和水分脅迫十分相似的新陳代謝變化[6]。尤其農(nóng)作物受到鹽脅迫時,在鹽離子豐富的環(huán)境中由于農(nóng)作物各組織中多種營養(yǎng)物質(zhì)可以抵抗該脅迫,將會導致植物營養(yǎng)不足,并且產(chǎn)生特殊毒害[7]。
菊芋(HelianthustuberosusL.)為菊科向日葵屬草本植物,又名洋姜,起源于北美。菊芋生物量大、管理成本低、易轉(zhuǎn)化,是一種理想的非糧能源植物[8-11]。此外,菊芋具有極強的無性繁殖能力和抗逆性,特別是抗旱、抗鹽堿、耐貧瘠等[12],在鹽堿地種植菊芋可產(chǎn)生明顯的經(jīng)濟效益和生態(tài)效應[13]。關于菊芋對單鹽、單堿或是鹽堿混合的響應機理研究報道較多[6, 13-17],但是使用氯化鈉和硫酸鈉中性復合鹽模擬西部鹽漬土對具有不同遺傳背景菊芋種質(zhì)影響的研究較為鮮見。人們總是有意或無意地改變作物的株型以獲得較高的產(chǎn)量,植物為了適應新環(huán)境會在產(chǎn)量和抗逆性之間權(quán)衡[18]。為了探究不同遺傳背景菊芋種質(zhì)耐鹽性的差異及其耐鹽機理,本試驗選擇具有不同遺傳背景的兩種基因型菊芋LZJ028和LZJ017為研究對象,其中LZJ028有性繁殖能力強且營養(yǎng)品質(zhì)高,LZJ017塊莖產(chǎn)量高[19],測定這兩種基因型菊芋幼苗形態(tài)指標、氣體交換參數(shù)及葉綠素熒光參數(shù)等生理指標,探究在鹽脅迫下菊芋生理生態(tài)響應機理,為進一步篩選耐鹽菊芋和改良西部鹽漬土提供理論指導。
1 材料與方法
1.1 試驗材料
選擇具有不同遺傳背景的兩種基因型菊芋為試驗材料,LZJ028有性繁殖能力強且營養(yǎng)品質(zhì)高,推測更傾向于野生化,LZJ017具有較高的塊莖產(chǎn)量,推測傾向于經(jīng)歷了較長時間的人工馴化[19],菊芋來源及編號見表1。

表1 試驗材料編號和來源
1.2 試驗設計
2015年4月1日~11月20日在蘭州大學榆中校區(qū)試驗站遮雨棚內(nèi)進行盆栽試驗。用規(guī)格相同的塑料盆(內(nèi)徑30cm,深45cm),分別裝風干混勻的沙土17 kg(土與沙比例為3∶1)。選取20 g左右、大小一致的完整新鮮菊芋塊莖,用自來水及蒸餾水沖洗干凈,經(jīng)體積分數(shù)為0.05%的多菌靈進行塊莖表面滅菌,然后栽種于塑料盆中。試驗處理使用NaCl和Na2SO4(摩爾比為2∶1)中性復合鹽按照土壤含鹽量的質(zhì)量分數(shù)設置為:低度鹽化即土壤含鹽量2 g·kg-1(2‰)、中度鹽化即土壤含鹽量4 g·kg-1(4‰)、重度鹽化即土壤含鹽量6 g·kg-1(6‰)三個鹽分梯度[20],無鹽處理為對照(CK),每個處理種植8盆,共計64盆,每盆定株一顆。每隔兩天于18∶00時,采用稱重法補充水分,土壤含水量保持在試驗用土最大持水量的70%。待菊芋出苗25 d后,每個處理選取3株長勢均一的植株取樣并測定苗期各指標。
1.3 指標測定
1.3.1 形態(tài)指標和生物量的測定 利用卷尺測定菊芋幼苗株高和冠幅、游標卡尺測定其基徑,用葉面積儀(Li-3000a, Li-COR USA)測定其整株葉面積。幼苗地上部分以單株為單位分別稱葉和莖鮮重,并于115℃殺青30 min,85℃烘干至恒重,稱其生物量干重。
1.3.2 生理生化指標的測定 每個處理采集菊芋幼苗植株上部相同位置(自植株頂部第3~5片)完全展開葉進行液氮保存,然后帶回實驗室采用茚三酮法測定葉片游離脯氨酸含量、NBT光還原法測定SOD活性、蒽酮法測定可溶性糖和丙酮比色法測定葉綠素含量[21]。
1.3.3 氣體交換和葉綠素熒光參數(shù)的測定 在形態(tài)指標測定前,利用便攜式光合作用測定系統(tǒng)(Li-6400, Li-COR USA)于上午10∶00選取相同位置(自植株頂部第3片)完全展開葉測定各處理菊芋幼苗的凈光合速率(Pn)、蒸騰速率(Tr)和氣孔導度(Gs),水分利用效率(WUE)由Pn/Tr計算所得[22]。利用植物效率分析儀(Handy PEA, Hansatech, UK)測定葉綠素熒光參數(shù)指標,包括初始熒光(Fo)、最大熒光(Fm)、可變熒光(Fv)、最大光化學效率(Fv/Fm)、潛在光化學效率(Fv/Fo)和電子傳遞光量子產(chǎn)量(φEo)。
1.4 數(shù)據(jù)處理
采用Excel 2013和SPSS17.0進行數(shù)據(jù)處理和分析,并用單因素方差分析(One-way ANOVA)和最小顯著性差異法(LSD法)分析差異顯著性,用Origin 8.0作圖。數(shù)據(jù)用平均值±標準誤表示。
2 結(jié)果與分析
2.1 鹽脅迫對兩種基因型菊芋幼苗形態(tài)特征和地上生物量的影響
隨著鹽濃度升高,兩種基因型菊芋幼苗株高、葉面積、冠幅和基徑均呈下降趨勢,且存在基因型間差異(圖1)。與對照相比,LZJ028株高、葉面積和冠幅在低鹽(2‰)脅迫時無顯著差異,中、高鹽(4‰和6‰)脅迫時顯著降低,高鹽(6‰)脅迫下,分別下降24.8%, 29.67%和27.98%;而LZJ017株高、葉面積和冠幅在各鹽度下均顯著降低,如:高鹽脅迫下,其株高、葉面積和冠幅分別下降27.56%, 63.18%和45.20%。高鹽脅迫下,LZJ017基徑與對照相比顯著下降,而LZJ028無顯著差異。在對照和低鹽處理組,LZJ017的株高、葉面積和基徑顯著高于LZJ028,但是在中、高鹽脅迫時,LZJ028葉面積顯著高于LZJ017,而高鹽脅迫時兩基因型菊芋株高、冠幅和基徑無顯著差異。
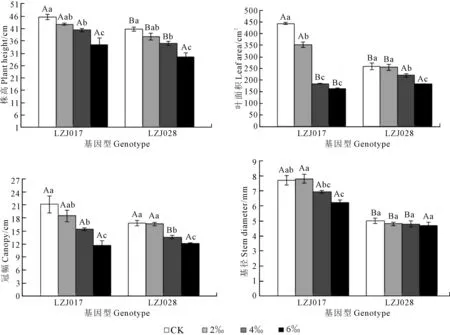
注:同指標數(shù)據(jù)柱上小寫字母不同表示處理間差異顯著(P<0.05);同指標數(shù)據(jù)柱上大寫字母不同表示基因型間差異顯著(P<0.05)。下同。Note: Different small letters show the significant difference between treatments (P<0.05); Different capitals show the significant difference between genotypes (P<0.05). The same as below.圖1 鹽脅迫對菊芋幼苗株高、葉面積、冠幅和基徑的影響Fig. 1 Effects of salt stress on plant height, leaf area, canopy and stem diameter of jerusalem artichoke seedlings
低鹽脅迫下,兩種基因型菊芋葉片和莖的干鮮物質(zhì)積累與其對照相比差異不明顯,但中、高鹽脅迫下各指標含量呈顯著下降趨勢(表2)。與對照相比,中、高鹽脅迫下LZJ028葉鮮物質(zhì)積累分別減少10.19%和24.75%,而LZJ017分別減少28.08%和59.32%,表明LZJ028葉鮮物質(zhì)積累減少幅度明顯小于LZJ017。葉干物質(zhì)和莖干、鮮物質(zhì)積累均呈現(xiàn)類似規(guī)律,即LZJ028減少幅度同樣小于LZJ017。鹽脅迫LZJ028形態(tài)指標和地上生物量積累在低鹽脅迫時與對照無顯著差異,而在高鹽脅迫時所測各指標下降幅度均低于LZJ017。
2.2 鹽脅迫對兩種基因型菊芋幼苗生理特性的影響
隨鹽濃度升高,兩種基因型菊芋SOD活性、游離脯氨酸和可溶性糖含量呈上升趨勢(表3)。高鹽脅迫下,LZJ028 SOD活性為對照的2.87倍,而LZJ017升高幅度較小,為對照的1.41倍;LZJ017脯氨酸含量升高幅度明顯低于LZJ028,兩基因型在高鹽度下分別為對照的9.26和43.99倍。對照和低鹽處理時,LZJ028與LZJ017可溶性糖含量無顯著差異,但在中、高鹽脅迫時LZJ028可溶性糖含量顯著高于LZJ017的含量。
表2鹽脅迫對菊芋幼苗地上部分鮮物質(zhì)和干物質(zhì)含量的影響
Table 2 Effects of salt stress on fresh and dry biomass of aerial part of jerusalem artichoke seedlings
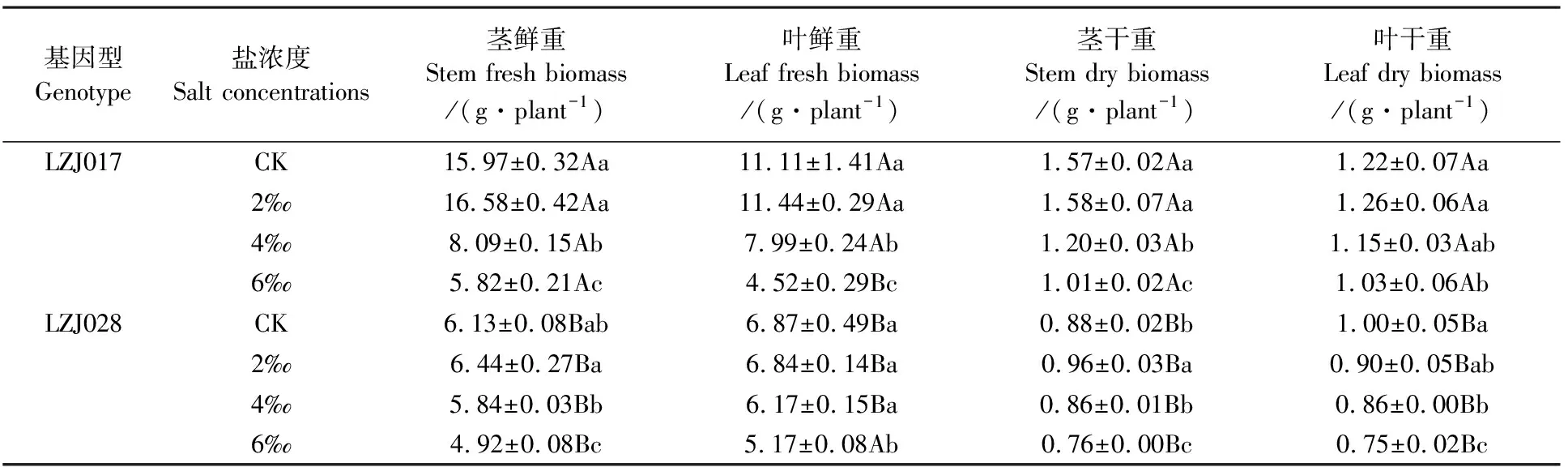
基因型Genotype鹽濃度Salt concentrations莖鮮重Stem fresh biomass/(g·plant-1)葉鮮重Leaf fresh biomass/(g·plant-1)莖干重Stem dry biomass/(g·plant-1)葉干重Leaf dry biomass/(g·plant-1)LZJ017CK15.97±0.32Aa11.11±1.41Aa1.57±0.02Aa1.22±0.07Aa2‰16.58±0.42Aa11.44±0.29Aa1.58±0.07Aa1.26±0.06Aa4‰8.09±0.15Ab7.99±0.24Ab1.20±0.03Ab 1.15±0.03Aab6‰5.82±0.21Ac4.52±0.29Bc1.01±0.02Ac1.03±0.06AbLZJ028CK6.13±0.08Bab6.87±0.49Ba0.88±0.02Bb1.00±0.05Ba2‰6.44±0.27Ba6.84±0.14Ba0.96±0.03Ba 0.90±0.05Bab4‰5.84±0.03Bb6.17±0.15Ba0.86±0.01Bb0.86±0.00Bb6‰4.92±0.08Bc5.17±0.08Ab0.76±0.00Bc0.75±0.02Bc
注:同列小寫字母不同表示處理間差異顯著(P<0.05);同列大寫字母不同表示基因型間差異顯著(P<0.05)。下同。
Note:In same column different small letters show the significant difference between treatments (P<0.05); In same column different capitals show the significant difference between treatments (P<0.05). The same as below.
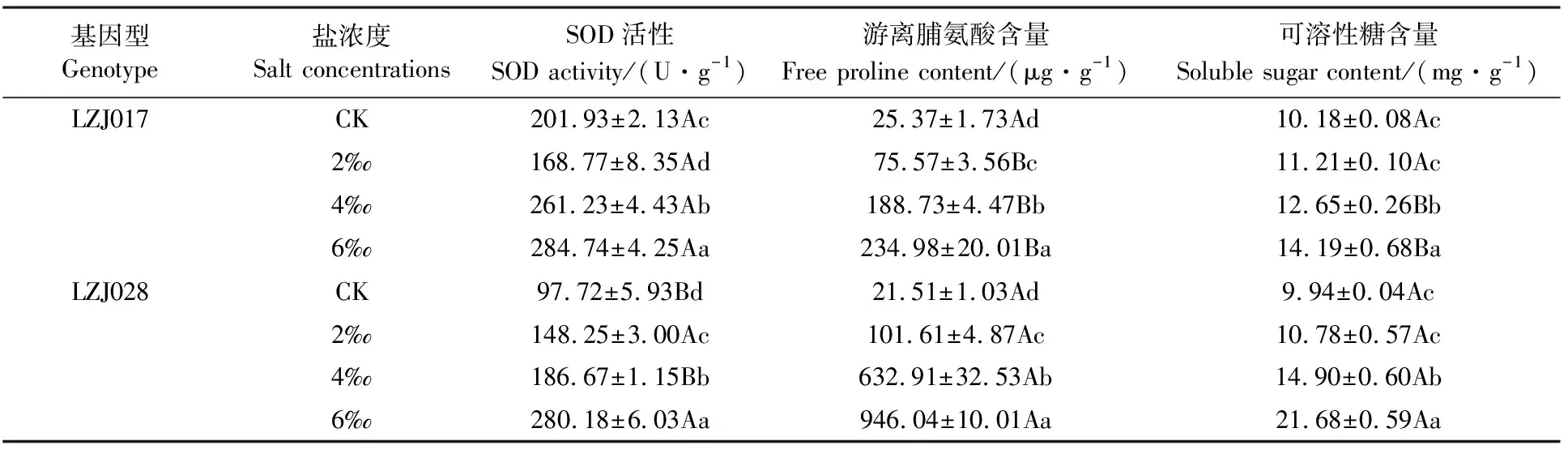
表3 鹽脅迫對菊芋幼苗SOD活性、游離脯氨酸和可溶性糖含量的影響
2.3 鹽脅迫對兩種基因型菊芋幼苗氣體交換參數(shù)的影響
兩種基因型菊芋葉片凈光合速率(Pn)和氣孔導度(Gs)隨著鹽濃度升高均呈下降趨勢(圖2),LZJ017的蒸騰速率(Tr)隨著鹽濃度升高而下降,LZJ028的蒸騰速率大體呈下降趨勢,在鹽濃度為6‰時其Tr比4‰有所升高。高鹽脅迫下,LZJ028Pn下降幅度較小,與對照比下降18.27%;而LZJ017下降26.29%,降幅較大。高鹽脅迫下LZJ028Gs和Tr分別下降29.00%和13.71%,而LZJ017分別下降47.00%和23.16%,降幅顯著大于LZJ028。兩種基因型菊芋葉片瞬時水分利用效率(WUE)隨著鹽濃度升高呈先上升后下降的趨勢(圖2)。LZJ028 WUE最大值出現(xiàn)在鹽濃度為4‰,而LZJ017 WUE最大值出現(xiàn)在2‰;且在各處理下LZJ028 WUE明顯高于LZJ017。相關性分析表明Pn與Gs、葉綠素含量變化均呈顯著正相關關系(圖3)。
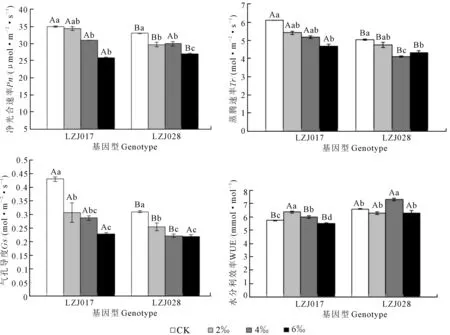
圖2 鹽脅迫對菊芋幼苗氣體交換參數(shù)的影響Fig. 2 Effects of salt stress on gas exchange parameters of jerusalem artichoke seedlings
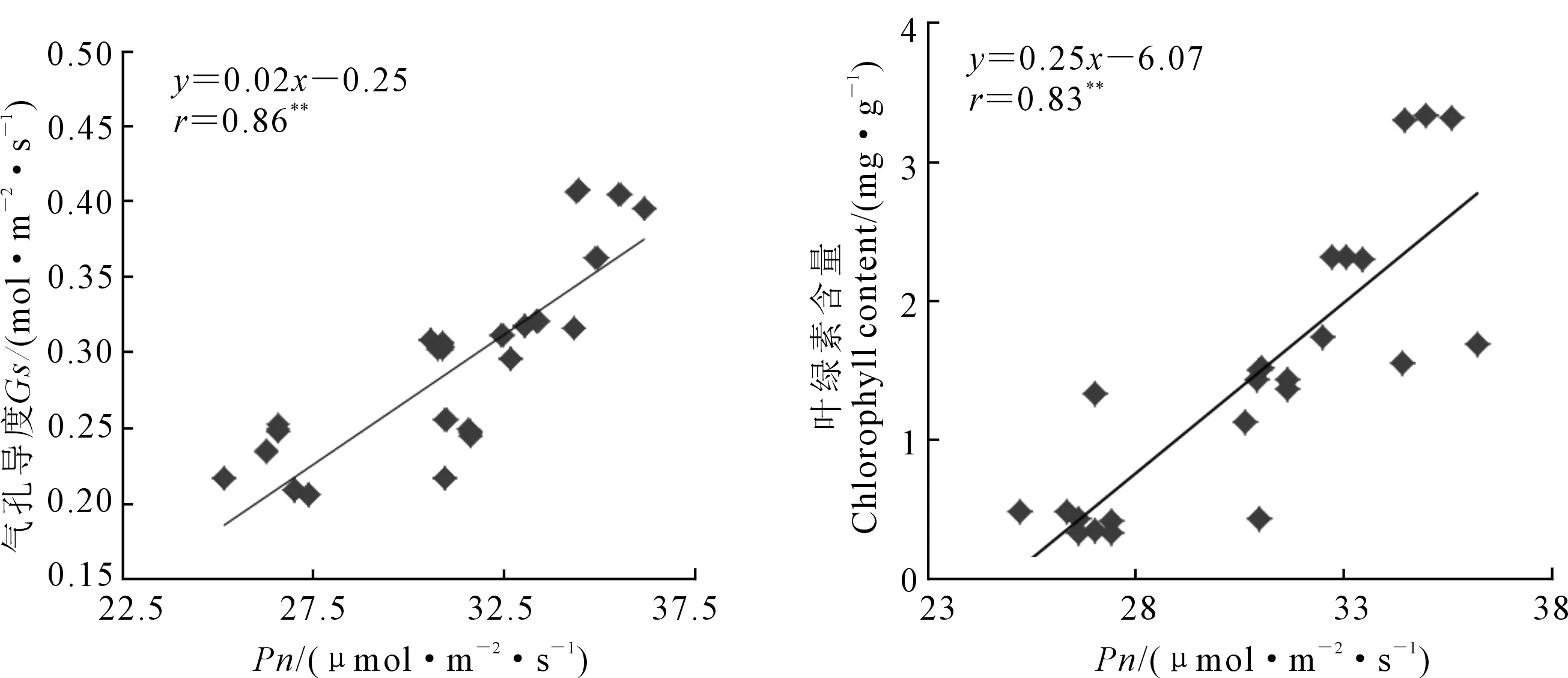
注: **表示相關性顯著(P<0.01)。Note:** means significant correlation (P<0.01).圖3 Pn與Gs和Chl的相關性分析Fig. 3 Relationships between Pn and Gs, Pn and Chl
2.4 鹽脅迫對兩種基因型菊芋幼苗葉綠素熒光參數(shù)的影響
兩種基因型菊芋葉片葉綠素含量隨鹽濃度升高而顯著下降(圖4)。在對照和低鹽處理時,LZJ017的葉綠素含量顯著高于 LZJ028,但當鹽濃度達到4‰和6‰時二者葉綠素含量無顯著差異,可知LZJ017的下降幅度明顯大于LZJ028。

圖4 鹽脅迫對菊芋幼苗葉綠素含量的影響Fig. 4 Effects of salt stress on chlorophyll content of jerusalem artichoke seedlings
隨著鹽濃度升高,兩種基因型菊芋Fv/Fo、Fv/Fm和Fm均呈下降趨勢,尤其在高鹽脅迫時顯著下降(表4)。高鹽脅迫下,LZJ028的Fv/Fo和Fv/Fm與對照相比分別下降47.88%和9.81%,而LZJ017下降78.04%和42.59%。在對照和低鹽脅迫時,LZJ017與LZJ028的Fv/Fo和Fv/Fm無顯著差異,當中、高鹽脅迫時, LZJ017該指標顯著低于LZJ028,可見LZJ017的降幅較大。LZJ028下降幅度小于LZJ017,說明高鹽脅迫下LZJ028葉片PSⅡ的光化學效率穩(wěn)定性較好。兩種基因型菊芋Fo隨鹽脅迫的加劇均呈上升趨勢(表4),LZJ028上升幅度顯著小于LZJ017,如:高鹽脅迫下其只比對照升高0.05倍,說明其PSⅡ反應中心受損傷程度較小。LZJ028φEo變化不明顯,而LZJ017在中、高鹽脅迫時顯著下降,也說明LZJ028 PSⅡ電子傳遞受到影響較小。
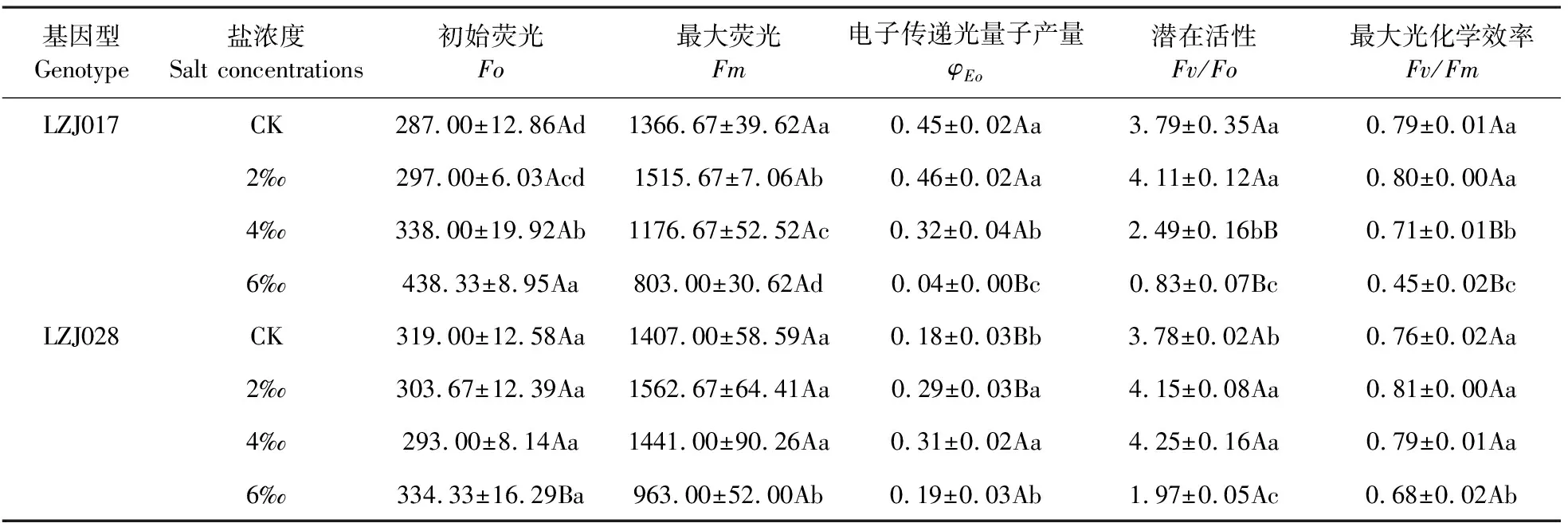
表4 鹽脅迫對菊芋幼苗熒光參數(shù)的影響
3 討論與結(jié)論
植物通過光合作用獲得的能量大部分用于維持生命活動,只有一小部分用于生長發(fā)育及生物量積累等,當植物受到鹽脅迫時,用于生長發(fā)育的那部分能量會重新分配用于抵御逆境[23,24]。本研究結(jié)果顯示兩種基因型菊芋株高、葉面積和冠幅隨鹽濃度升高呈下降趨勢,與對照相比,LZJ028的株高、葉面積和冠幅在低鹽脅迫時無顯著差異,在高鹽脅迫時顯著降低,而LZJ017該指標在低鹽和高鹽脅迫下均顯著低于對照,且下降幅度高于LZJ028,說明LZJ028較耐鹽。生物量積累是維持植物生物活性和評價各種脅迫對植物影響的最重要指標[25]。菊芋地上生物量隨鹽脅迫的加劇而逐漸減少,且LZJ017降幅高于LZJ028,說明其受鹽脅迫影響較大。LZJ028的株高、冠幅、葉面積以及生物量積累受到的影響較小可能由于其自身較耐鹽,用于抵御鹽脅迫的能量比例低于LZJ017,使其可以有更多能量用于自身的生長發(fā)育。
鹽脅迫下,植物光合作用受到不同程度的抑制[26],氣孔導度是CO2進入植物體內(nèi)進行碳同化的關鍵限制因子[27]。兩種基因型菊芋隨著鹽濃度升高其凈光合速率下降。LZJ017和LZJ028在高鹽脅迫時,與對照相比Gs降低,與Win等[28]對于黑綠豆的研究結(jié)果一致,且兩種基因型菊芋Pn與Gs變化趨勢一致,表明其光合速率的下降受氣孔導度影響。葉綠素是反映光合強度的重要指標[29],本試驗顯示鹽濃度升高,菊芋葉片葉綠素含量下降,且兩種基因型菊芋的Pn與Chl具有顯著的正相關性,表明菊芋光合速率的下降也與葉綠體的活性下降有關。蒸騰作用是植物對水分吸收和運輸?shù)闹饕獎恿Γ梢蚤g接地為光合作用提供所需水分和礦質(zhì)元素[30]。本研究中隨著鹽濃度升高,兩種基因型菊芋Tr逐漸下降,與Win等[28]研究結(jié)果一致,表明高鹽脅迫時,菊芋降低蒸騰速率,從而維持各組織水分,在一定程度抑制了光合作用,但對其在逆境條件下生存有重要意義。在中、高鹽脅迫時,LZJ028的水分利用效率較高,而LZJ017較低,表明鹽逆境時LZJ028對于水分的利用效率較高,使其保持一定的含水量。
與“表觀性”的氣體交換參數(shù)相比,葉綠素熒光參數(shù)更具有反映“內(nèi)在性”的特點[31]。光系統(tǒng)Ⅱ(PSⅡ)是植物葉片進行有機物合成的光合器官重要組分之一,其功能穩(wěn)定性決定著植物光能的吸收和電子傳遞水平,從而限制植物有機物累積能力[32],且葉綠素熒光與光合作用中各種反應過程密切相關,能夠用來驗證植物受到脅迫時被傷害的程度[33, 34]。PSⅡ反應中心的破壞或可逆失活則能引起Fo的增加[35],F(xiàn)o的變化程度可用來鑒別植物的不同抵抗或耐鹽能力[14],φEo指吸收光能用于QA-以后的電子傳遞的能量比例,F(xiàn)v/Fm是反映PSⅡ光化學效率的穩(wěn)定指標。本研究結(jié)果顯示在高鹽脅迫下,與對照相比LZJ028φEo和Fv/Fm下降幅度較小且Fo變化不顯著,而LZJ017在中、高鹽脅迫時其變化幅度較大,說明LZJ028 PSⅡ電子傳遞受到的影響較小,PSⅡ的損傷不明顯,具有較高的耐鹽性,而LZJ017 PSⅡ受到較大損害。
光合系統(tǒng)受到破壞抑制了光合作用的原初反應[36],在光抑制過程中電子傳遞受阻導致能量滯留,產(chǎn)生大量活性氧[37]。SOD是植物體內(nèi)防御系統(tǒng)的第一道防御,是清除植物體內(nèi)活性氧的主要抗氧化酶[38]。本研究表明隨著鹽濃度升高,兩種基因型菊芋SOD活性均增強,與王恩軍等人[39]的研究結(jié)果一致。植物利用各種策略對抗土壤中的鹽,最重要的一個策略就是滲透調(diào)節(jié),所有細胞調(diào)控積累足夠的可溶性滲透物質(zhì)如脯氨酸和可溶性糖用于降低細胞滲透勢,保持細胞膨壓[23]。本試驗中兩種基因型菊芋葉片脯氨酸和可溶性糖含量隨鹽濃度升高均逐漸增加,與Muchate等人[40]的研究結(jié)果一致,且LZJ028的增幅大于LZJ017,表明LZJ028具有較高的耐鹽性。有研究表明隨著鹽脅迫程度加深,植物可溶性糖含量增加可能是淀粉含量和總糖含量積累引起的[38]。
綜上,鹽脅迫使菊芋葉片葉綠素含量和氣孔導度下降,破壞葉綠素合成系統(tǒng),且通過破壞PSⅡ的反應中心以及阻礙電子傳遞,使能量滯留產(chǎn)生大量活性氧,進而影響最大光化學效率和抑制光合作用,最終抑制菊芋生長。本研究通過對兩種基因型菊芋在不同濃度鹽脅迫下的形態(tài)特征、生理特性和光合特性進行研究和對比,結(jié)果表LZJ028較耐鹽,LZJ017對鹽較敏感,兩基因型菊芋在鹽脅迫下主要通過提高SOD抗氧化酶活性清除體內(nèi)的活性氧,且積累大量的可溶性滲透物質(zhì)脯氨酸和可溶性糖降低細胞滲透勢維持細胞膨壓,進而抵御逆境。
參考文獻:
[1] Athar H U R, Zafar Z U, Ashraf M. Glycinebetaine improved photosynthesis in canola under salt stress: Evaluation of chlorophyll fluorescence parameters as potential indicators [J]. Journal of Agronomy and Crop Science, 2015, 201(6):428-442.
[2] Munns R. Comparative physiology of salt and water stress[J]. Plant Cell and Environment, 2002. 25(2):239-250.
[3] 顧國安. 新疆鹽漬化土壤的形成及其防治[J]. 干旱區(qū)地理, 1984, 7(4):1-16.
[4] 樊自立,馬英杰,馬映軍. 中國西部地區(qū)的鹽漬土及其改良利用[J]. 干旱區(qū)研究, 2001, 18(3):1-6.
[5] Li Q, Tang Z, Hu Y, et al. Functional analyses of a putative plasma membrane Na+/H+antiporter gene isolated from salt tolerant Helianthus tuberosus[J]. Molecular Biology Reports, 2014,41(8):5097-5108.
[6] Xue Y F, Liu Z P. Antioxidant enzymes and physiological characteristics in two Jerusalem artichoke cultivars under salt stress [J]. Russian Journal of Plant Physiology, 2008,55(6):776-781.
[7] Tuteja N, Gill S S. Na+tolerance and Na+transport in higher plants [J]. Annals of Botany, 2003,91(5):503-527.
[8] Li X F, Hou S L, Su M, et al. Major energy plants and their potential for bioenergy development in China [J]. Environmental Management, 2010,46(4):579-589.
[9] 寇一翾, 呂世奇, 劉建全, 等. 寡糖類能源植物菊芋及其綜合利用研究進展[J]. 生命科學, 2014, 26(5):451-457.
[10] 呂世奇, 寇一翾, 楊彬, 等. 半干旱地區(qū)菊芋品系植株表型與光合特性分析[J]. 作物學報,2014, 40(10):1857-1864.
[11] 劉祖昕, 謝光輝. 菊芋作為能源植物的研究進展[J]. 中國農(nóng)業(yè)大學學報, 2012, 17(6):122-132.
[12] Zhuang D F, Jiang D, Liu L, et al. Assessment of bioenergy potential on marginal land in China [J]. Renewable & Sustainable Energy Reviews, 2011,15(2):1050-1056.
[13] Lu Y, Ye H J, Geng S B, et al. Effects of NaCl stress on growth, leaf photosynthetic parameters and ion distribution of Helianthus tuberosus seedling [J]. Journal of Plant Resources and Environmen, 2010,19(2):86-91.
[14] Long X H, Liu Z P, Zheng Q S, et al. Effects of seawater with different concentrations on growth and physiological and biochemical characteristics ofHelianthustuberosusseedlings [J]. Acta Ecologica Sinica, 2005, 25(8):1881-1889.
[15] 薛延豐, 劉兆普. 不同濃度NaCl和Na2CO3處理對菊芋幼苗光合及葉綠素熒光的影響[J]. 植物生態(tài)學報, 2008, 32(1):161-167.
[16] 吳成龍, 周春霖, 尹金來, 等. 堿脅迫對不同品種菊芋幼苗生物量分配和可溶性滲透物質(zhì)含量的影響[J]. 中國農(nóng)業(yè)科學, 2008, 41(3): 901-909.
[17] 黃增榮, 隆小華, 劉兆普, 等. KNO3對NaCl脅迫下兩菊芋品種幼苗生長及光合能力的影響[J]. 草業(yè)學報,2011,20(1):82-88.
[18] 王建永, 李樸芳, 程正國, 等. 旱地小麥理想株型與生長冗余[J]. 生態(tài)學報, 2015, 35 (8):2428-2437.
[19] 寇一翾. 菊芋種質(zhì)資源多樣性及高產(chǎn)量形成機理研究[D]. 蘭州:蘭州大學, 2013.
[20] 陳麗娟,馮起,成愛芳. 民勤綠洲土壤水鹽空間分布特征及鹽漬化成因分析[J]. 干旱區(qū)資源與環(huán)境,2013,27(11):99-105.
[21] 高俊鳳. 植物生理學實驗指導[M]. 北京:高等教育出版社, 2006.
[22] 曲桂敏, 束懷瑞, 王鴻霞. 鉀對蘋果樹水分利用效率及有關參數(shù)的影響[J]. 土壤學報, 2000, 37(2):257-262.
[23] Rana M, Matthew G. Salinity tolerance of crops - what is the cost? [J]. New Phytologist, 2015, 208(3):668-73.
[24] Jacoby R P, Taylor N L, Millar A H. The role of mitochondrial respiration in salinity tolerance [J]. Trends in Plant Science, 2011, 16(11):614-23.
[25] Gong B, Wen D, Vandenlangenberg K, et al. Comparative effects of NaCl and NaHCO3stress on photosynthetic parameters, nutrient metabolism, and the antioxidant system in tomato leaves [J]. Scientia Horticulturae, 2013,157(3):1-12.
[26] Chaves M M, Flexas J, Pinheiro C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell[J]. Annals of Botany, 2009, 103(4):551-60.
[27] 寇偉鋒, 劉兆普, 陳銘達, 等 . 不同濃度海水對油葵幼苗光合作用和葉綠素熒光特性的影響[J]. 西北植物學報, 2006, 26(1):73-77.
[28] Win K T, Oo A Z, Ookawa T, et al. Changes in hydraulic conductance cause the difference in growth response to short-term salt stress between salt-tolerant and -sensitive black gram (Vignamungo) varieties [J]. Journal of Plant Physiology, 2016, 193:71-78.
[29] Lin Z H, Chen L S, Chen R B, et al. CO2assimilation, ribulose-1,5-bisphosphate carboxylase/oxygenase, carbohydrates and photosynthetic electron transport probed by the JIP-test, of tea leaves in response to phosphorus supply [J]. Bmc Plant Biology, 2009, 9(4):43.
[30] 劉金祥, 陳偉云, 肖生鴻. 黑籽雀稗的光合生理特性研究[J]. 草業(yè)學報, 2009, 18(6):254-258.
[31] 沈允鋼, 施教耐, 許大全. 動態(tài)光合作用[M]. 北京:科學出版社,1998.
[32] 馬飛, 徐婷婷, 陳立同, 等. 低溫脅迫下二倍體雜交種高山松光系統(tǒng)Ⅱ功能穩(wěn)定性研究[J]. 西北植物學報, 2011, 31(6):1174-1179.
[33] 馬清, 樓潔瓊, 王鎖民. Na+對滲透脅迫下霸王幼苗光合特性的影響[J]. 草業(yè)學報, 2010, 19(3):198-203.
[34] 許耀照, 曾秀存, 郁繼華, 等. 水楊酸對高溫脅迫下黃瓜幼苗葉綠素熒光參數(shù)的影響[J]. 西北植物學報,2007,27(2):267-271.
[35] 李磊, 李向義, 林麗莎, 等. 兩種生境條件下6種牧草葉綠素含量及熒光參數(shù)的比較[J]. 植物生態(tài)學報, 2011, 35(6):672-680.
[36] Meloni D A, Oliva M A, Martinez C A, et al. Photosynthesis and activity of superoxide dismutase, peroxidase and glutathione reductase in cotton under salt stress [J]. Environmental & Experimental Botany, 2003,49(49):69-76.
[37] Duarte B, Santos D, Marques J C, et al. Ecophysiological adaptations of two halophytes to salt stress: Photosynthesis, PS II photochemistry and anti-oxidant feedback-Implications for resilience in climate change [J]. Plant Physiology & Biochemistry, 2013,67(3):178-188.
[38] Farhangi-Abriz S, Torabian S. Antioxidant enzyme and osmotic adjustment changes in bean seedlings as affected by biochar under salt stress[J]. Ecotoxicology & Environmental Safety, 2017:64-70.
[39] 王恩軍,李善家,韓多紅,等. 中性鹽和堿性鹽脅迫對黑果枸杞種子萌發(fā)及幼苗生長的影響[J]. 干旱地區(qū)農(nóng)業(yè)研究,2014,32(06):64-69.
[40] Muchate N S, Nikalje G C, Rajurkar N S, et al. Physiological responses of the halophyte Sesuvium portulacastrum, to salt stress and their relevance for saline soil bio-reclamation[J]. Flora - Morphology, Distribution, Functional Ecology of Plants, 2016, 224:96-105.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
