基于Ecopath模型的長江口生態系統營養結構和能量流動研究
2018-06-29 02:54:36王思凱
海洋漁業 2018年3期
徐 超, 王思凱, 趙 峰, 宋 超, 莊 平
(1. 南京農業大學無錫漁業學院, 江蘇無錫 214081;2. 中國水產科學研究院東海水產研究所,農業部東海與長江口漁業資源環境科學觀測實驗站, 上海 200090)
河口是海洋生態系統的重要組成部分,以其獨特的地理位置、水動力條件和物質基礎,為眾多海洋生物的生存和繁衍提供了有利的生存條件[1]。長江口作為我國最大的河口,位于暖溫性的黃海生態系統與暖水性的東海生態系統的交界處,受長江徑流及黃海冷水團相互消長的影響,環境因素變化劇烈,生態環境錯綜復雜。源源不斷的長江徑流帶來了大量營養物質,維持了長江口生態系統的巨大生產力,孕育了豐富的餌料資源,使長江口成為許多重要經濟魚類的產卵場、索餌場、育幼場和洄游通道[2]。近年來對長江口的研究主要集中在對水生生物群落結構[3-4]、水生動物食性[5-6]及水環境質量評價[7-8]等方面。然而,隨著捕撈強度的增大、環境污染和漁場海洋環境條件的變化,長江口生物多樣性和生態系統環境質量受到前所未有的脅迫和影響,因而對長江口生態系統結構和功能的研究就顯得十分必要。
Ecopath with Ecosim (EwE)是一組物質平衡模型軟件[9]。它以食物網為主線,根據營養動力學原理,用一組聯立方程平衡系統中各功能組生物生產和消費的能量流動,直接構造以物質平衡為基礎的生態系統模型。這種方法最早由POLOVINA提出,用來評估穩定狀態水域生態系統組成的生物量和食物消耗,經過與ULANOWICZ的能量分析生態學理論結合,逐步發展成為一種生態系統營養成分流動分析方法[10]。該方法可用于研究:生態系統的規模、穩定性和成熟度;物流、能流的分布和循環;系統內部的捕食和層級關系;各層級間能量流動的效率;中間生態位的競爭及彼此互利或危害的程度等,被譽為新一代水域生態系統研究的核心工具[11]。目前,Ecopath模型在水生生態系統中得到了廣泛應用,國內外學者利用Ecopath模型描述了渤海[12]、南黃海[13]、東海[14]和南海[15]等不同中國近海海域生態系統的能流過程,而對河口水域的研究相對較少。因此,本文通過利用Ecopath with Ecosim (版本6.4.3)軟件構建2016-2017年長江口及鄰近水域生態系統的能流網絡模式,并對此歷史時期生態系統結構與功能進行綜合評估,旨在通過對該河口生態系統的結構、能量流動特征的定量分析,為我國長江口生態學研究積累新的資料,并為長江口漁業資源的利用和環境管理提供科學的理論依據。
1 材料與方法
1.1 研究區域
分別于2016年8月(夏季)、11月(秋季)、2017年2月(冬季)、5月(春季)在長江口水域(31°00′00″~31°48′00″N、121°00′00″~122°48′00″E)設置的22個站點進行4次拖網調查(圖1)。調查內容包括浮游植物、浮游動物、漁業生物、底棲動物等。漁業生物(主要包括魚類、蝦類、蟹類和頭足類)使用的調查船功率為45 kW,單船桁桿拖網為桁桿長6 m的雙囊網,網具總長15 m,高2 m,網囊孔徑2 cm,每個位點拖網時間0.5 h,平均航速3 kn;分別在漲潮和落潮時使用Ⅰ型浮游動物網和Ⅱ型浮游植物網采集浮游生物;底棲生物樣品則是通過各站位用采泥器取兩次做一個樣品,經0.5 mm篩網沖洗獲取。
圖1 長江口采樣地點示意圖Fig.1 Location of sampling sites in the Yangtze Estuary
1.2 數據來源
建立Ecopath模型需要真實可靠的數據,收集的數據應盡可能的完整,覆蓋生態系統中的所有生物。模型需要輸入的參數包括生物量(B)、生產量與生物量比值(P/B)、消費量與生物量比值(Q/B)和生態營養效率(EE)這4個參數,需要輸入其中的3個,另外一個參數(通常為EE)由模型估計得出;同時食物組成矩陣(DC)和捕撈量也必須輸入。
本研究的數據來源于2016年8月-2017年5月在長江口及鄰近水域調查所得,對水生生物作了詳細的生物學測定和胃含物分析。其中,魚類、蝦蟹類和頭足類的生物量數據通過采樣調查獲得,轉化公式如下:
M=G/[S(1-E)]
(1)
式(1)中,M為生物量,G為調查生物量,S為網口的掃海面積,E為逃逸率。
隱埋性底棲動物與浮游生物的生物量分別靠底泥采樣和浮游生物垂直拖網樣品估計,有機碎屑生物量用有機碎屑與初級生產力的經驗公式估算[16],食物組成矩陣通過詳細的胃含物食性分析法所得,捕撈量通過《中國漁業統計年鑒》中的資料估算,對于難以估計的參數(P/B與Q/B),則主要通過國際水生資源管理中心網站(www.fishbase.org)和歷史資料數據所得[16-18],生態營養效率(EE)作為未知參數,通過模型估計得出。
1.3 研究方法
1.3.1 Ecopath模型基本原理
Ecopath模型定義生態系統是由一系列生態關聯的功能組組成,并遵循熱力學定律研究生態系統能量流動與網絡分析。功能群應涵蓋生態系統能流的全過程[11]。該模型假定在給定一段時間內(通常為1a)系統能量守恒。它包含兩個核心方程,一個方程描述各功能群生產量的流向[下式(2)],一個方程用來描述各功能群的能量守恒[下式(3)]。
Pi=Yi+M2iBi+Ei+BAi+M0iBi
(2)
Qi=Pi+Ri+Ui
(3)
式(2)、式(3)中,Pi表示生產量,Yi為漁獲量,M2i為捕食死亡率,Bi為生物量,Ei為凈遷出量,BAi為生物量累積,M0i為其它死亡率, 即功能群未被生態系統所利用的那部分能量所占的比重;Qi表示第i功能群對其它功能群的消費量,Pi表示生產量,Ri表示呼吸量,Ui表示未同化量。
1.3.2 功能組的劃分
按照研究需要,將生態學或分類地位上相似的物種加以整合,也可以把單個物種或者單個物種的某個年齡階段(成體或幼體)作為獨立的功能組[19],其中功能組中必須包含1個或數個碎屑功能組[20]。本研究主要根據不同物種生態學與生物學特性(食性、棲息地、死亡率等)的相似性以及在漁業方面的重要程度來劃分功能組,共14個功能組(表1)。
1.3.3 Ecopath模型的調試
對于模型的建立,首先要考慮的是生態營養效率EE值應介于0~1之間,其次要考慮總效率GE值,它被定義為生產量與消耗量(P/Q)之比。在許多情況下GE值應介于0.1~0.3之間[21]。在食性數據、捕撈數據、基礎數據3大類數據輸入模型后,模型并未成為平衡狀態,仍有生態轉移效率大于1的功能組,長江口生態通道模型調試主要從食物組成著手[22],對各功能組的食物構成做出調整,以達到模型平衡狀態。
2 結果與分析
2.1 長江口生態系統的營養級
由表2可見,2016-2017年長江口及鄰近海域生態系統各功能群的營養級集中在1~3.93。其中,浮游植物與碎屑營養級為1;浮游動物營養級為2.18;隱埋性底棲生物、頭足類及蝦、蟹類的營養級集中在2.47~3.11;魚類營養級分布較廣,除中上、下層浮游生物碎屑食性魚類的營養級較低位于2.41~2.47,其它魚類功能群營養級為3.49~3.93。

表1 長江口2016-2017年生態系統模型的功能組Tab.1 Functional groups of ecosystem model in 2016-2017 in the Yangtze Estuary

表2 長江口生態系統Ecopath模型功能組估算參數Tab.2 In and out parameters of the Ecopath of the Yangtze Estuary ecosystem
注:加粗字體為模型計算的參數
Note: The bold font is the parameter of the model calculation
2.2 長江口生態系統營養級間的物質流動
Lindeman模式可以將復雜的食物網簡化成線性的食物鏈。通過營養級分解[23],將各功能組的能流重組,形成生態系統各營養級的能流分布。2016-2017年長江口生態系統能流分布主要集中在營養級Ⅰ~Ⅲ中(圖2),當營養級為Ⅳ甚至更高時能流可以忽略不計。其中來自營養級Ⅰ級的能流在系統總能流中所占比例最高,為53.67%;其次為第Ⅱ營養級所占比例為45.19%。整個營養級Ⅰ流入到營養級Ⅱ的物流通量為1 637 t·km-2·a-1;從營養級Ⅱ到營養級Ⅴ的總流量隨其營養級的增加而依次降低,分別為36.52 t·km-2·a-1、6.463 t·km-2·a-1、0.967 t·km-2·a-1。此外,營養級Ⅱ、Ⅲ、Ⅳ的能流轉換效率依次分別為2.31%、19.30%和17.80%,平均傳輸效率為13.14%。
2.3 長江口生態系統營養級間的能量轉化效率
營養級間的能流轉化效率是指該營養級輸出和被攝食的流量與總流量之間的比值,體現了該營養級在生態系統中的被利用的效率[24]。通過比較不同營養級間的能量轉換效率(表3)可以看出,長江口初級生產者到第Ⅱ營養級的轉換效率為2.2%,而碎屑到第Ⅱ營養級的轉換效率為4.3%,生產者和碎屑總能量轉換效率為2.3%;第Ⅱ營養級到第Ⅲ營養級轉換效率很高,綜合轉換效率為19.3%;第Ⅲ營養級到第Ⅳ營養級綜合轉換效率為17.8%;第Ⅳ營養級到第Ⅴ營養級綜合轉化效率為9.1%。可見,低營養級間的能流轉化效率相對較低,而高營養級間的能流轉化效率相對較高。來自初級生產者和碎屑的能量轉換效率分別為9.1%和10.6%,且長江口生態系統的總體能量轉換效率為9.3%。根據生態金字塔能量轉換的“十分之一定律”[25],該河口能量的利用效率未達到最適程度。
2.4 長江口生態系統食物網結構
從圖3可以看出,長江口生態系統食物網結構較為復雜,營養流動主要有兩條途徑:一是從浮游植物開始的牧食食物鏈,如:浮游植物—浮游動物—中上層浮游生物碎屑食性魚類—中上層混合食性魚類;二是從碎屑開始的碎屑食物鏈,如碎屑—隱埋性底棲生物—蝦類—底層游泳生物食性魚類。表4中聯結指數(CI)和系統雜食指數(SOI)分別為0.345、0.321,說明長江口食物鏈普遍較短。因此,一個物種數量的變動或者消失都可能影響到整個長江口生態系統的食物網結構。

圖2 長江口生態系統各營養級間的物質流動Fig.2 Trophic flows transmitted through aggregated trophic levels in the Yangtze Estuary注:P表示初級生產者浮游植物;D表示有機碎屑Note: P denotes primary producer phytoplankton; D denotes organic detritus

來源Source營養級Trophic levelⅡⅢⅣⅤⅥⅦⅧ生產者 Producer2.219.018.59.25.4-碎屑 Detritus4.321.413.27.75.6-總流動 Total flow2.319.317.89.15.48.6-碎屑所占能流比 Proportion of detritus to total flow0.14總轉換效率 Transfer efficiencies (Calculated as genometric mean for TL Ⅱ-Ⅳ)來自初級生產者 From primary producers9.1來自碎屑 From detritus10.6系統 Ecosystem9.3
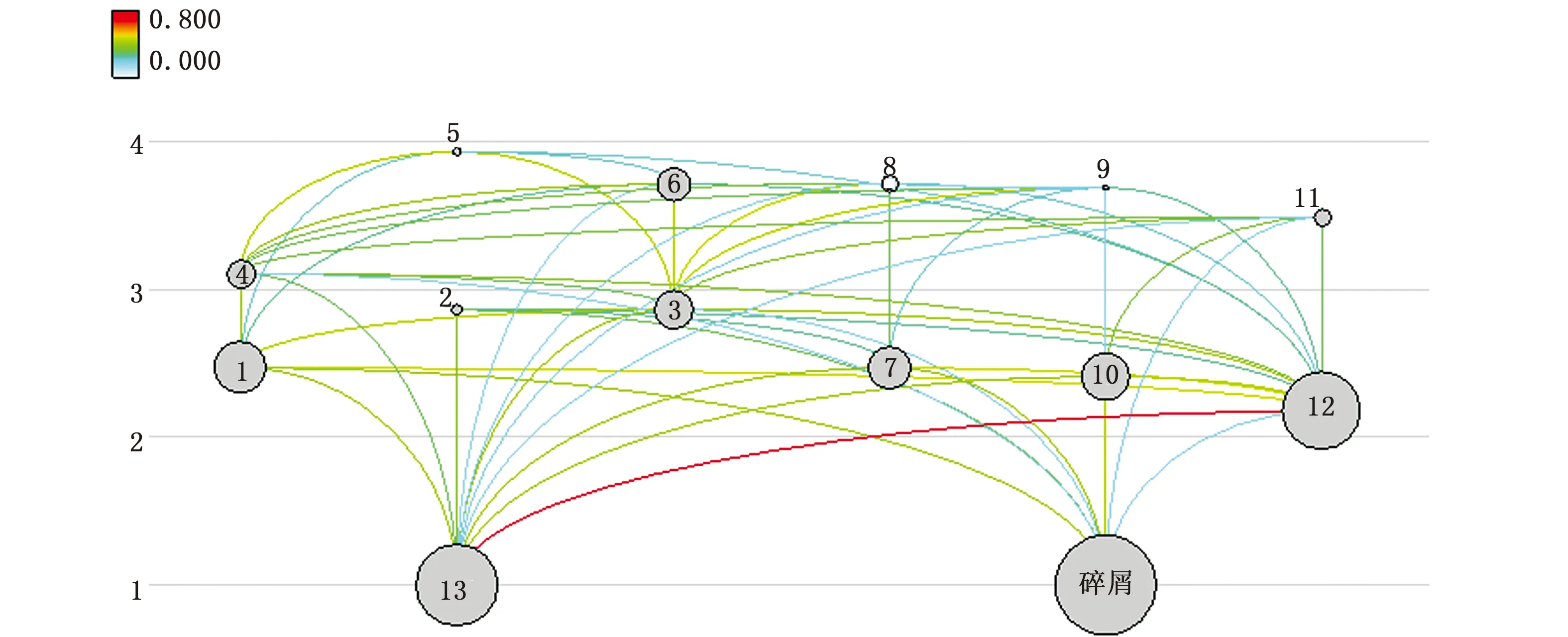
圖3 長江口生態系統的食物網Fig.3 Food web of the Yangtze Estuary ecosystem注:1.隱埋性底棲生物; 2.頭足類; 3.蝦類; 4.蟹類; 5.底層游泳生物食性魚類; 6.底層混合食性魚類; 7.中下層浮游生物碎屑食性魚類;8.中下層游泳生物食性魚類; 9.中下層混合食性魚類; 10.中上層浮游生物碎屑食性魚類; 11.中上層混合食性魚類; 12.浮游動物; 13.浮游植物Note: 1.Benthic producers; 2.Cephalopods; 3.Shrimps; 4.Crabs; 5.Benthic predators; 6.Benthic mixed predators; 7.Middle-lower plankton detritus predators; 8.Middle-lower predators; 9.Middle-lower mixed predators; 10.Middle-upper plankton detritus predators; 11.Middle-upper mixed predators; 12. Zooplankton; 13.Phytoplankton
2.5 長江口生態系統發育成熟度
生態系統的很多特征均可用于表征生態系統的成熟度[26]。生態系統發育早期,通常系統的凈初級生產量Pp較高,而系統的呼吸R消耗較小,因此Pp/R>1;隨著生態系統的發育不斷趨于成熟,其生物量不斷累積,呼吸消耗逐步增大,則Pp/R接近1[27]。通過軟件自動計算(表4)可見,2016-2017年長江口的Pp/R值為1.245大于1,表明該河口生態系統趨于不成熟。此外,從生態系統發育的信息學特征來研究生態系統的發育成熟度,聯結指數(CI)和系統雜食指數(SOI)是表征系統內部聯系復雜程度的指標,越是成熟的生態系統,CI和SOI的數值越接近于1,其各功能組間的聯系越強,系統越趨于穩定[28]。長江口生態系統的CI和SOI分別為0.345和0.321,表明該生態系統的初級生產力較高,但各功能組的聚合度較低,聯結程度較為松散。

表4 2016-2017長江口生態系統的總體特征Tab.4 General properties of the Yangtze Estuary ecosystem in 2016-2017
3 討論
本文通過Ecopath模型分析了長江口2016-2017年生態系統營養結構和能量流動,結果表明:長江口尚處于不成熟的發展階段,其生態系統穩定性較低,生物資源種類組成較為簡單,系統的初級生產力/呼吸量比值大于1,說明系統中仍有較多的剩余能量未消耗。
本文計算生態系統功能組的營養級集中在1~3.93,這與張效嘉等[16]構建的長江口1985-1986年Ecopath模型所計算的營養級集中在1~4.52相差較大;此外,羅秉征等[29]運用胃含物分析法研究了長江口20世紀90年代魚類的營養級大小,集中在3.1~4.5。通過對比發現,長江口水生生物的營養級逐漸降低,而水生生物平均營養級的數值大小反映了漁業資源開發利用程度。隨著捕撈強度的增大,長江口水生生物資源量處于退化狀態。李建生等[30]通過對長江口2003年漁場漁業資源的調查,并與2000-2002年同期比較,發現漁獲物資源量以及種類數都有不同程度的降低。目前以底拖網為主的捕撈漁業使底層的大型經濟漁業資源迅速減少,漁獲物間的捕食關系發生改變,食物網組成結構也由以往肉食性、長壽的底層魚類向浮游生物食性、短壽的上層魚類轉變[3];此外,長江口及其鄰近海域由于長期受到環境污染和漁場海洋環境條件的變化,如三峽大壩的成功截流、洋山港深水碼頭以及長江口深水航道治理工程的建設等因素,水生生物群落結構發生顯著變化[31]。RYDER等[32]指出,過度捕撈和環境退化使生物群落的生態系統失去恢復力和完整性,生態系統的穩定性變差,使依賴生態系統產出的漁業產量在質和量兩個方面具有不可預見的變化。因而長江口水生生物的營養級降低很大程度上應該是環境因素和捕撈因素的共同作用結果[33];此外,本研究選取的參數如P/B、Q/B,以及研究水域與張效嘉等[16]略有差異,因而計算出的營養級不同。
長江口生態系統能量流動和轉換通過牧食食物鏈和碎屑食物鏈共同實現,且主要發生在5個營養級間。其中,直接來自碎屑僅占總能流的14%,來自初級生產者的占總能流的86%。這與張效嘉等[16]研究的1985-1986年長江口生態系統能量流動存在差異,其研究結果表明,來自碎屑的占總能流的43%,來自初級生產者的占總能流的57%。此外,韓瑞等[22]研究發現,三峽工程蓄水前后碎屑食物鏈在系統能量流動占比由44%下降到41%。三峽工程蓄水后,長江入海徑流量減少,長江沖淡水面積減少,陸源營養物質和懸浮顆粒的輸入減少,使得生態系統能量傳遞更依賴牧食食物鏈[22]。此外,長江口2016-2017年生態系統中生物量、生產量和流量基本上均呈金字塔型分布,即底層營養級的生物量和流量較大,越到頂級越小,基本上符合能量和生物量金字塔規律。
在生態系統演替過程中,群落結構和功能的同時變化是有規律的,在“幼態”時期,生物多表現為r-對策。隨著發育的進程,k-對策生物的比例開始增加[27]。在功能方面,隨著演替的不斷進行,初級生產量與呼吸量的比值從大于1逐漸發展到接近于1。而群落凈生產量從高到低,能流渠道從線性到網狀,碎屑食物鏈的重要性逐漸增加[34]。目前,長江口生態系統在人類活動和自然環境因素的影響下發生了逆行演替的現象,系統特征從較為“成熟”的狀態向“幼態”轉變。2016-2017年長江口生態系統的CI、SOI值分別為0.345、0.321;表明長江口生態系統的成熟度和穩定性以及系統抵抗外力干擾的能力較低。在自然進化過程中,生態系統的演替是一個較為緩慢的過程,但是自然環境變動和人類活動可以加速演替的進程[27]。因此,如何可持續開發與利用長江口漁業資源,維護其生態系統的健康和穩定,合理控制其生態演替的速度,是目前急需解決的問題。從長江口生態系統的結構、物流和能流的特征來看,應依據生態學原理,包括生態演替、食物鏈網結構、物質循環和轉化、生物間的交互營養關系等原則,從不斷完善生態系統的結構組成、促進功能的發展成熟角度出發,建立可持續發展的生態系統,從而保護長江口生態環境、實現人與自然的和諧發展。
參考文獻:
[1] KIMMERER W J. Physical, biological, and management responses to variable Freshwater flow into the San Francisco estuary [J]. Estuary, 2002, 25(6B): 1275-1290.
[2] 羅秉征, 沈煥庭. 三峽工程與河口生態環境[M]. 北京: 科學出版社, 1994.
LUO B Z, SHEN H T. Impacts of the Three Gorges Project on estuarine ecology and environment [M]. Beijing: Science Press, 1994.
[3] 金顯仕, 單秀娟, 郭學武, 等. 長江口及其鄰近海域漁業生物的群落結構特征[J]. 生態學報, 2009, 29(9): 4761-4772.
JIN X S, SHAN X J, GUO X W,etal. Community structure of fishery biology in the Yangtze River estuary and its adjacent waters [J]. Acta Ecological Sinica, 2009, 29(9): 4761-4772.
[4] 吳 強, 李顯森, 王 俊, 等. 長江口及鄰近海域無脊椎動物群落及其生物多樣性研究[J]. 水生態學雜志, 2009, 2(2): 73-79.
WU Q, LI X S, WANG J,etal. Community structure and biodiversity of invertebrate resources in the Yangtze River Estuary and its adjacent waters [J]. Journal of Hydroecology, 2009, 2(2): 73-79.
[5] 羅 剛, 莊 平, 章龍珍, 等. 長江口中華鱘幼魚的食性組成及攝食習性[J]. 應用生態學報, 2008, 19(1): 144-150.
LUO G, ZHUANG P, ZHANG L Z,etal. Diet composition and feeding habits of juvenile Acipenser sinensis in Yangtze River estuary [J]. Chinese Journal of Applied Ecology, 2008, 19(1): 144-150.
[6] 洪巧巧, 莊 平, 楊 剛, 等. 長江口中國花鱸食性分析[J]. 生態學報, 2012, 32(13): 4181-4190.
HONG Q Q, ZHUANG P, YANG G,etal. Feeding habits of Lateolabrax maculates in Yangtze River estuary [J]. Acta Ecological Sinica, 2012, 32(13): 4181-4190.
[7] 劉啟貞, 李九發, 戴志軍, 等. 長江口顆粒態金屬污染五時空分布規律分析[J]. 海洋環境科學, 2009, 28(3): 251-255.
LIU Q Z, LI J F, DAI Z J,etal. Spatial and temporal distribution of particulate metal pollutants in Changjiang Estuary [J]. Marine Environmental Scinece, 2009, 28(3): 251-255.
[8] 畢春娟, 陳振樓, 許世遠, 等. 長江口潮灘大型底棲動物對重金屬的累積特征[J]. 應用生態學報, 2006, 17(2): 309-314.
BI C J, CHEN Z L, XU S Y,etal. Heavy metals accumulation in macrobenthos in intertidal flat of Yangtze Estuary [J]. Chinese Journal of Applied Ecology, 2006, 17(2): 309-314.
[9] 陳 亮, 劉 一, 禹 娜, 等. 分水江水庫浮游動物群落結構的初步研究及水質評價[J]. 華東師范大學學報(自然科學版), 2010(6): 72-82.
CHEN L, LIU Y, YU N,etal. Preliminary study on the metazoan zooplankton community structure of Fenshuijiang Reservoir and water quality assessment [J]. Journal of East China Normal University (Natural Science), 2010(6): 72-82.
[10] PAYLY D, CHRISTENSEN V, WALTER C J. Ecopath, ecosim and ecospace as tools for evaluating ecosystem impact of fisheries [J]. ICES Journal of Marine Science, 2000(57): 697-706.
[11] CHRISTENSEN V, PAULY D. ECOPATHⅡ-a software for balancing steady-state ecosystem models and calculating network characteristics [J]. Ecological Modelling, 1992, 61(3): 169-185.
[12] 仝 齡, 唐啟升. 渤海生態通道模型初探[J]. 應用生態學報, 2000, 11(3): 435-440.
TONG L, TANG Q S. A Preliminary approach on mass-balance ecopath model of the Bohai Sea [J]. Chinese Journal of Applied Ecology, 2000, 11(3): 435-440.
[13] LIN Q, JIN X, ZHANG B. Trophic interactions, ecosystem structure and function in the southern Yellow Sea [J]. Chinese Journal of Oceanology and Limnology, 2013, 31(1): 46-58.
[14] CHEN J H, CHENG W W, PITCHER T J. Mass-balance ecosystem model of the East China Sea [J]. Progress in Natural Science, 2009(10): 1271-1280.
[15] 陳作志, 邱永松. 南海北部生態系統食物網結構,能量流動及系統特征[J]. 生態學報, 2010, 30(18): 4855-4865.
CHEN Z Z, QIU Y S. Assessment of the food-web structure, energy flows, and system attribute of northern South China Sea ecosystem [J]. Acta Ecologica Sinica, 2010, 30(18): 4855-4865.
[16] 張效嘉, 線薇薇. 1985-1986年長江口生態系統能流網絡分析[J]. 海洋科學, 2016, 40(7): 60-72.
ZHANG X J, XIAN W W. Energy flow and network analysis of the Yangtze Estuary ecosystem during 1985-1986 [J]. Marine Sciences, 2016, 40(7): 60-72.
[17] 林 群, 金顯仕, 郭學武, 等. 基于Ecopath模型的長江口及毗鄰水域生態系統結構和能量流動研究[J]. 水生態學雜志, 2009, 2(2): 28-36.
LIN Q, JIN X S, GUO X W,etal. Study on the structure and energy flow of the Yangtze River Estuary and adjacent waters ecosystem based on ecopath model [J]. Journal of Hydroecology, 2009, 2(2): 28-36.
[18] 李 睿, 韓 震, 程和琴, 等. 基于ECOPATH模型的東海區生物資源能量流動規律的初步研究[J]. 資源科學, 2010, 32(4): 600-605.
LI R, HAN Z, CHENG H Q,etal. A preliminary study on biological resources energy flows based on the ecopath model in the East China Sea [J]. Resources Science, 2010, 32(4): 600-605.
[19] CHRISTENSEN V, WALTERS C, AHRENS R,etal. Database-driven models of the world's large marine ecosystems [J]. Ecological Modelling, 2009, 9(10): 1984-1996.
[20] LI Y K, YONG C, BING S,etal. Ecosystem structure and functioning of Lake Taihu (China) and the impacts of fishing [J]. Fisheries Research, 2009(95): 309-324.
[21] VILLY C, CARL J W, DANIEL P. Ecopath with ecosim: A user’s guide [D]. Vancouer Canada: Fisheries Centre University of British Columbia, 2004: 36-42.
[22] 韓 瑞, 陳求穩, 王 麗, 等. 基于生態通道模型的長江口水域生態系統結構與能量流動分析[J]. 生態學報, 2016, 36(15): 4907-4918.
HAN R, CHEN Q W, WANG L,etal. Analysis of the ecosystem structure and energy flow of the Yangtze River estuary and adjacent seas, based on the Ecopath model [J]. Acta Ecologica Sinica, 2016, 36(15): 4907-4918.
[23] ULANPWICZ R. Ecosystem trophic foundations: Lindeman exonerata. Complex ecology: The part-whole relation in ecosystems prentice-hall [J]. Englewood Cliffs, 1995: 549-560.
[24] JACKSON B C, KIRBY M X, BERGER W H,etal. Historial overfishing and the recent collapse of coastal ecosystems [J]. Science, 2001(293): 629-637.
[25] LINDEMAN R L. The trophic-dynamic aspect of ecology [J]. Ecology, 1942(23): 339-417.
[26] ODUM E P. The strategy of ecosystem development [J]. Science, 1969(164): 262-270.
[27] 武 震, 賈佩嶠, 胡忠軍, 等. 基于Ecopath模型分析分水江水庫生態系統結構和功能[J]. 應用生態學報, 2012, 23(3): 812-818.
WU Z, JIA P Q, HU Z J,etal. Structure and function of Fenshuijiang Reservoir ecosystem based on the analysis with Ecopath model [J]. Chinese Journal of Applied Ecology, 2012, 23(3): 812-818.
[28] CHRISTENSEN V. Ecosystem maturity-towards quantification[J]. Ecological Modelling, 1995, 77(1): 3-32.
[29] 羅秉征, 韋 晟, 竇碩增. 長江口魚類食物網與營養結構的研究[J]. 海洋科學集刊, 1997(3): 143-153.
LUO B Z, WEI S, DOU S Z. Study on food web and trophic structure of fish in the Changjiang river estuary [J]. Studia Marina Sinica, 1997(3): 143-153.
[30] 李建生, 程家驊. 長江口漁場漁業生物資源動態分析[J]. 海洋漁業, 2005, 27(1): 33-37.
LI J S, CHENG J H. Analysis on the dynamic of fishery biological resources on the fishing grounds of the Changjiang estuary [J]. Marine Fisheries, 2005, 27(1): 33-37.
[31] 陳吉余, 陳沈良. 近20年來長江口生態系統變化[J]. 凈水技術, 2002, 1(4): 1-3.
CHEN J Y, CHEN S L. Ecological environment changes in the Changjiang Estuary over recent 20 years [J]. Water Purification Technology, 2002, 1(4): 1-3.
[32] RYDER R A, KERR S R, TAYLOR W W,etal. Community consequences of fish stock diversity [J].Canadian Journal of Fisheries and Aquatic Sciences, 1981(38): 1856-1866.
[33] FINNEY B P, GREGORY-EAVES I, DOUGLAS M S V,etal. Fisheries productivity in the northeastern Pacific Ocean over the past 2 200 years [J]. Nature, 2002(416): 729-733.
[34] 陳作志, 邱永松, 賈曉平, 等. 基于Ecopath模型的北部灣生態系統結構和功能[J]. 中國水產科學, 2008, 15(3): 460-468.
CHEN Z Z, QIU Y S, JIA X P,etal. Structure and function of Beibu Gulf ecosystem based on Ecopath model [J]. Journal of Fishery Sciences of China, 2008, 15(3): 460-468.
猜你喜歡
今日農業(2021年11期)2021-08-13 08:53:34
保健醫苑(2021年7期)2021-08-13 08:48:02
甘肅教育(2020年14期)2020-09-11 07:57:42
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
中國生殖健康(2019年8期)2019-01-07 01:18:24
幸福(2018年33期)2018-12-05 05:22:46
海峽姐妹(2016年5期)2016-02-27 15:20:20
時代英語·高二(2015年1期)2015-03-16 00:08:11
領導文萃(2015年4期)2015-02-28 09:19:05
