異源染色體附加對普通小麥氮磷吸收能力和利用效率的影響
2018-06-13 02:13:52劉彩云李慧娟杜瑩瑩張書良胡銀崗
麥類作物學報 2018年5期
劉彩云,李慧娟,杜瑩瑩,張書良,劉 鵬,胡銀崗,3
(1.德州市農業科學研究院,山東德州 253011; 2.西北農林科技大學農學院/旱區作物逆境生物學國家重點實驗室,陜西楊凌 712100; 3.中國旱區節水農業研究院,陜西楊凌 712100)
化肥的使用極大地提高了小麥產量,但是片面追求高產而過量施用肥料,不但造成了資源的巨大浪費,還引起了一系列環境問題。在農業生態系統中,施用的氮肥只有小部分被作物吸收,超過50%的氮被淋洗到土壤中[1]。磷肥的當季利用率只有15%~30%,大部分的磷被堿性土壤中的鈣鹽及酸性土壤中的鐵、鋁等的氫氧化物固定或被土壤膠體吸附,難以被作物再次吸收利用[2]。相對于單純地依賴于肥料的施用,培育氮、磷高效的作物品種是提高養分吸收和利用效率的根本途徑,而氮、磷高效小麥種質資源的發掘是小麥養分高效育種的基礎。
盡管普通小麥中存在著豐富的氮、磷效率的基因型變異,但是長期在人工環境下的選擇育種,使得小麥的基因庫日趨狹窄,一些養分高效利用的相關基因可能已經丟失[3]。小麥的野生近緣種屬和原始祖先種有著豐富的遺傳多樣性,常比小麥更能適應貧瘠惡劣的環境[4]。Gorny等[5]研究了小麥的野生近緣種和原始祖先種苗期的氮、磷效率,發現與普通小麥相比,T.monococcum、Ae.tauschii、Ae.speltoides和Ae.sharonensis等種質具有更發達的根系,耐養分脅迫的能力更強。柳 鵬等[6]根據磷效率和地上部干物質積累量篩選出3份人工合成小麥種質,表明山羊草屬含有與磷高效有關的基因。
通過遠緣雜交和染色體工程創造的攜帶有外源染色體或者遺傳物質的種質資源,如附加系等,也繼承了小麥近緣種屬耐瘠薄的特點。通過對成套的小麥異附加系的研究發現,黑麥的1R、2R和7R染色體,Ag.elongatum的4E和6E染色體攜帶有耐低磷脅迫的基因[7-8]。Wang等[9]通過對部分小麥異附加系的磷效率進行鑒定,篩選出9個高磷效率的種質。趙化田等[10]通過進行耐低氮基因型的苗期水培篩選試驗,發現攜帶冰草、黑麥、偃麥草等外源血統的材料氮利用效率較高。由此可見,小麥近緣種屬異源染色體種質含有豐富的氮、磷高效基因及優良性狀,可應用于小麥氮、磷高效的遺傳改良,因此對氮、磷高效異源染色體種質的篩選尤為重要。
本研究通過苗期水培試驗,以中國春為對照,對43份小麥異源染色體附加系的氮、磷吸收能力和利用效率進行了評價,以期篩選出氮、磷高效種質,為小麥養分高效的遺傳研究和育種應用奠定基礎。
1 材料與方法
1.1 供試材料
供試材料為43個中國春背景的普通小麥異附加系(經鑒定,其染色體組成均為2n=44)及其共同受體親本中國春,附加的染色體來自賴草屬(Leymus)、冰草屬(Agropyron)、山羊草屬(Aegilops)、大麥屬(Hordeum)、黑麥屬(Secale)共5個小麥近緣屬。
1.2 試驗方法
1.2.1 溫室水培試驗
水培試驗在溫室中進行,培養所用溶液為改良的Hoagland營養液。氮素設置高氮(HN)、中氮(MN)和低氮(LN)3個水平;磷素設置中磷(MP)和低磷(LP)2個水平[11]。試驗共6個處理,即HNMP,MNMP,LNMP,HNLP,MNLP及LNLP。氮以NH4+和NO3-的形式提供,磷以H2PO4-的形式提供,不同處理NH4++NO3-及H2PO4-的供給濃度見表1。試驗采用隨機區組設計,每個處理設置3次重復。
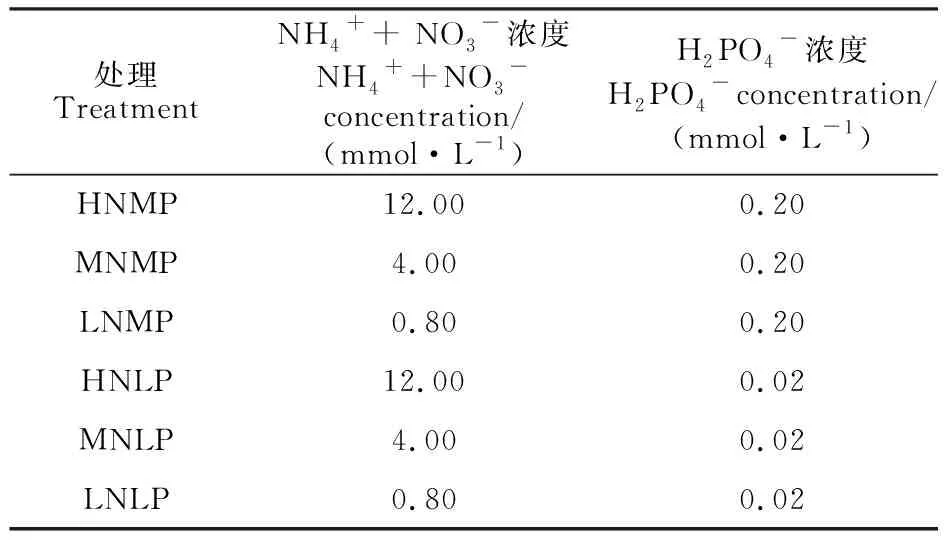
表1 不同處理的氮、磷濃度設置Table 1 The concentration of N and Pused in different treatments
選取大小一致的種子,75%酒精消毒1 min,放入鋪有濕潤濾紙的培養皿中催芽。待萌發幼苗的胚芽鞘長到3~4 cm時,每份種質選取長勢一致的3株幼苗,將其固定到泡沫塑料板(50×40 cm)上的播種孔中。然后將栽滿幼苗的泡沫塑料板放入盛有60 L營養液的黑色塑料根箱中(50×40×30 cm)。用空氣泵向營養液中通氣,以形成健康的根系。培養期間,溫度設置為白天25 ℃,夜晚15 ℃,培養時間為4周。
1.2.2 性狀測量
收獲前,用葉綠素儀SPAD-502活體測定幼苗主莖最頂端已完全展開葉片的相對葉綠素含量SPAD(Soil and Plant Analyzer Development)值。培養4周后,分別收獲莖葉和根系部分。測量苗高、最大根長,統計葉片數。收獲的根系用無菌ddH2O沖洗3~4遍,徹底沖掉附著在根系上的離子。然后將樣品放入烘箱,105 ℃殺青15 min,80 ℃烘干至恒重。稱量獲得莖葉干重和根干重,計算總干重和根冠比。
將供試材料干樣磨成粉,過0.25 cm篩子,測定氮、磷含量。測定時稱取一定量的樣品,經H2SO4與H2O2消解,用凱氏定氮法測定全氮含量[12],用釩鉬黃比色法測定全磷含量[13]。
1.2.3 數據分析
氮(磷)吸收能力用植株吸氮(磷)總量(uptake amount,UpA)來表示,其計算公式如下[14]:
UpA=SDW×CS+RDW×CR
式中,SDW為莖葉干重;RDW為根干重;CS為莖葉中的氮(磷)濃度;CR為根系中的氮(磷)濃度。
氮/磷利用效率(utilization efficiency,UtE)的計算公式如下[9,14]:
UtE=SDW/UpA
式中,SDW為莖干重;UpA為植株氮(磷)吸收總量。
各測量性狀不同處理間的比較和分析采用Microsoft Excel 2010以及SAS 8.1軟件進行。
2 結果與分析
2.1 不同氮、磷處理下的植株表型變異
同中氮中磷處理相比,各處理下大多數小麥異附加系出現苗高降低、根長縮短以及次生根數目減少等癥狀。低磷脅迫下(高氮低磷、中氮低磷、低氮低磷),附加系老葉開始變黃,并且從葉尖處逐漸萎蔫;根系呈現深褐色,形成大量根毛。
不同供氮處理下,各測量性狀之間差異顯著(表2)。除根冠比外,所有測量性狀的最低值均出現在高氮處理,表明過量氮會抑制小麥整個植株的生長。低氮處理的苗高、莖干重和總干重顯著低于中氮處理;根長、葉數、SPAD值及根冠比顯著高于中氮處理。不同磷水平下,各測量性狀之間的差異也達到顯著水平(表2)。莖葉干重、根冠比在低磷水平下較中磷水平顯著增加,其余所測性狀在低磷水平下均顯著降低。氮、磷互作下,除根冠比外,各性狀的最低值均出現在高氮低磷處理。小麥異附加系對低磷和低氮脅迫的表現不同:除根冠比外,其余性狀值在中氮低磷處理下均顯著低于低氮中磷處理,根冠比的增加也主要源于莖葉干重的顯著降低;同中氮低磷相比,低氮低磷只顯著增加了根長,表明缺磷比缺氮對小麥異附加系的生長影響更大。
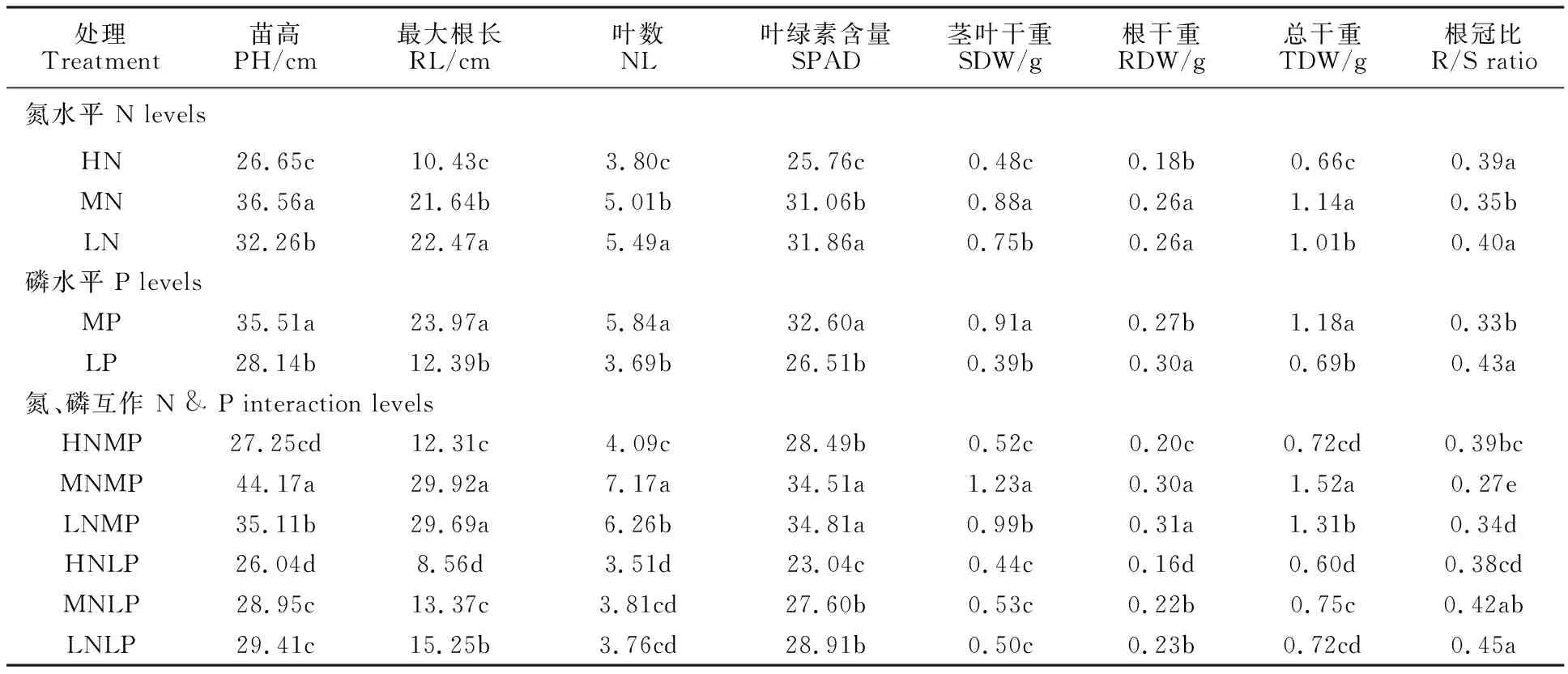
表2 不同氮、磷處理對小麥異附加系表型性狀的影響Table 2 Effect of different N and P levels on the phenotypic traits of wheat addition lines
表中數據為每一性狀的平均值;同列數據后的不同字母表示在0.05水平上差異顯著。
Each value represents mean value.Different letters in same column indicated significant difference at 0.05 level by Duncan test.
PH:Plant hight;RL:Max of root lenghth;NL:Leaf number;SDW:Shoot dry weight;RDW:Root dry weight;TDW:Total dry weight;R/S:Root/Shoot.
2.2 中氮及低氮處理下的植株總干重
植株干重是植物營養吸收能力和利用效率的綜合反映。中氮中磷處理下,小麥異附加系植株總干重在0.34~2.71 g之間;低氮中磷處理下,植株總干重變異范圍為0.32~2.54 g(表3)。低氮水平下,能夠生產較多干物質的種質為氮高效種質;中氮水平下,能夠生產較多干物質的種質為高生產潛力種質[5]。以中國春在中氮中磷和低氮中磷處理下的總干重為參照,可以將供試的小麥異附加系分為4類(圖1)。其中,第Ⅰ類包含13個小麥異附加系,其特點是氮高效且具有高產潛力,能夠穩定高效利用氮肥。第Ⅱ類包括10個小麥異附加系,其特點是氮高效但生產潛力低,更適應低氮環境。第Ⅲ類包含3個小麥異附加系,其特點是對氮缺乏十分敏感,更適應氮充足環境。歸為第Ⅳ類的15個小麥異附加系,對氮素極不敏感,為典型的“氮低效、低生產潛力”種質。
2.3 小麥異附加系的氮吸收能力和利用效率
不同供氮條件下,各小麥異附加系的氮吸收能力各不相同。中氮中磷處理下,小麥異附加系的植株吸氮總量在32.36~164.26 mg之間(表4);與對照中國春(105.76 mg)相比,氮吸收能力顯著增加的附加系有10個,占供試材料的23.26%;其中攜帶Ps.huashanicaE染色體的附加系的吸氮總量最高,較中國春增幅達55.31%;其次為攜帶Ag.intermediumB染色體的附加系,增幅為54.14%。低氮中磷處理下,同一附加系的氮吸收能力較中氮中磷處理明顯降低,范圍在20.24~86.29 mg之間;與中國春相比,9個小麥異附加系(20.93%)的植株吸氮總量顯著提高。攜帶H.chilense7Hch染色體的種質吸氮總量最高,其次為攜帶Ae.searsii1SS和Ae.longissima2S染色體的附加系。綜合來看,Ps.HuashanicaE、Ag.intermediumB、Ae.longissima2S、Ae.searsii1SS及Ae.searsii4SS異附加系在氮充足及脅迫條件下,顯著提高了氮吸收總量,表明這些異源染色體上可能攜帶與氮吸收能力相關的基因。
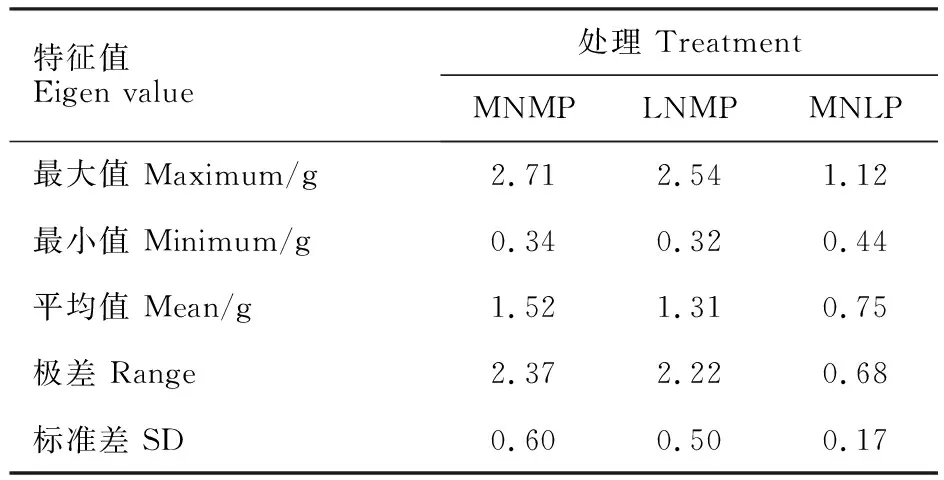
表3 中氮中磷、低氮中磷及中氮低磷處理下小麥異附加系總干重的變異Table 3 Variation of total dry weight in MNMP,LNMP and MNLP treatments

圖1 小麥異附加系在低氮中磷及中氮中磷處理下總干重的分布Fig.1 Biplots and classified groups of wheat additionlines based on total dry weight(TDW) underLNMP and MNMP treatments
中氮中磷處理下,小麥異附加系的氮利用效率范圍在10.50~17.93 g TDW·g-1N之間(表4);與中國春(16.04 g TDW·g-1N)相比,只有2個小麥異附加系的氮利用效率顯著降低,其余附加系無明顯差異。低氮中磷處理下,Ps.huashanicaC、Ps.huashanicaE、Ae.searsii1SS及Ag.elongatum6E等4個小麥異附加系的氮利用效率顯著高于中國春;攜帶Ps.huashanicaC異源染色體的附加系氮利用能力最強。
2.4 中磷及低磷處理下的植株總干重
中氮低磷處理下,小麥異附加系的總干重較中氮中磷處理顯著降低,其變異范圍為0.44~1.12 g(表3)。以中國春在中氮中磷和中氮低磷處理下的總干重為參照,可以將供試材料分成4類(圖2)。中氮中磷處理下,16個小麥異附加系的植株總干重高于中國春,其中5個小麥異附加系在中氮低磷下也能維持高于中國春的總干重(第Ⅰ類)。10個小麥異附加系在中氮中磷處理下的植株總干重低于中國春,但在中氮低磷處理下高于中國春,劃入第Ⅱ類。劃為第Ⅰ、Ⅱ類的小麥異附加系均具有“磷高效”的特點,其中攜帶有異源染色體Ag.intermediumB和Ae.searsii4SS的小麥異附加系表現最突出。對低磷脅迫最敏感的10個小麥異附加系劃入第Ⅲ類,該類種質適合磷充足環境;在中氮中磷和中氮低磷處理下表現均低于中國春的15個小麥異附加系歸于第Ⅳ類,為磷低效和低產潛力種質。
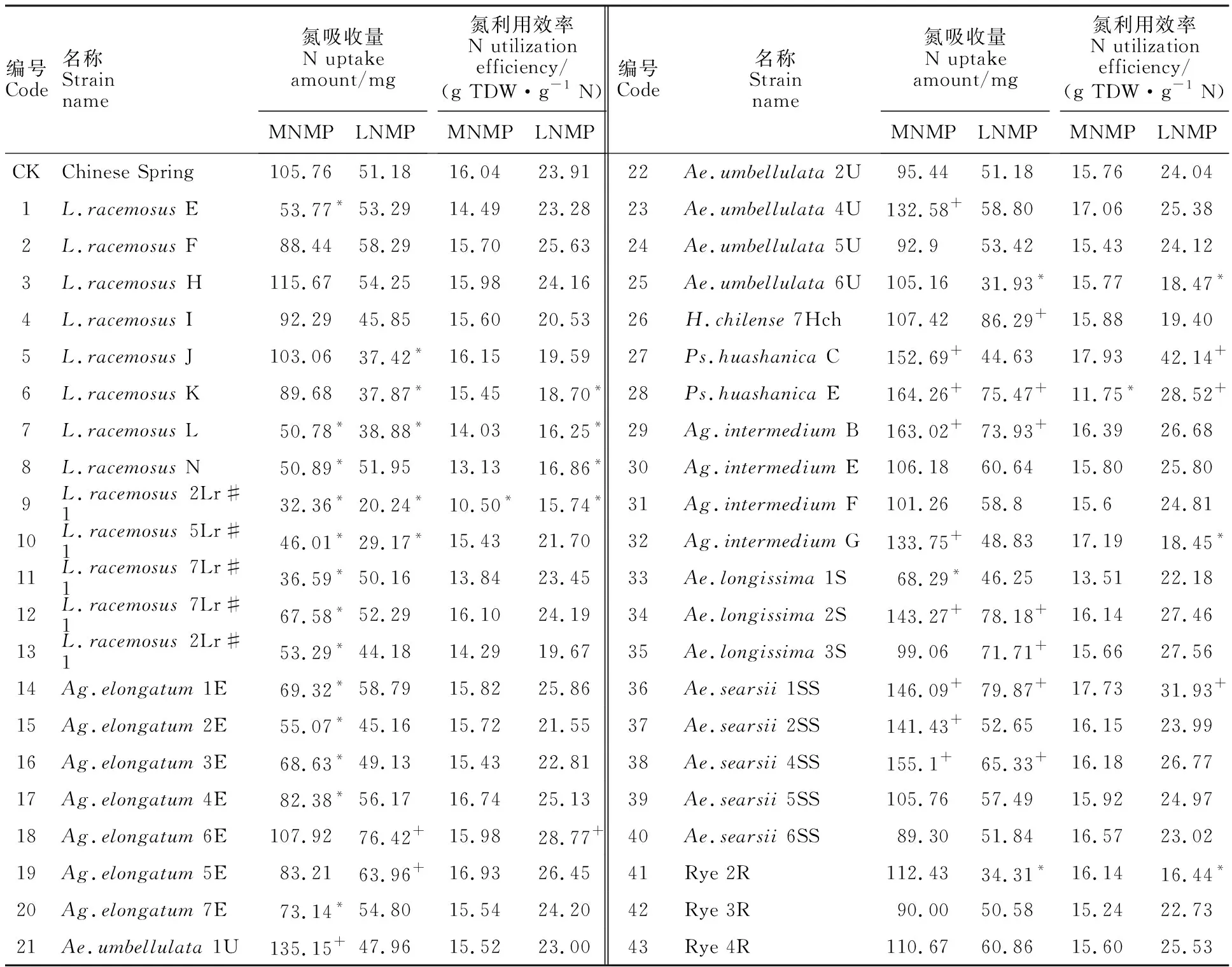
表4 不同處理下小麥異附加系的氮吸收和利用效率Table 4 Nitrogen uptake amount and utilization efficiency of wheat addition lines under different treatments
+表示顯著高于中國春,*表示顯著低于中國春,P<0.05。表5同。
Values with * and + in the same column indicate significantly lower and higher than CS at 0.05 level,respectively.The same in table 5.
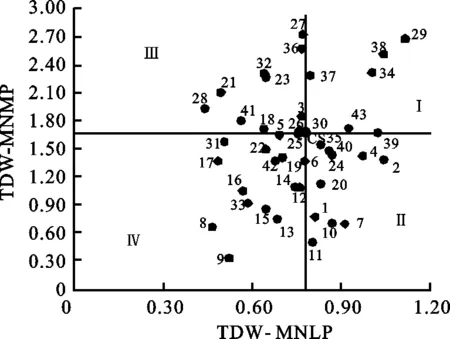
圖2 小麥異附加系在中氮低磷及中氮中磷處理下總干重分布圖Fig.2 Biplots and classified groups of wheat additionlines based on total dry weight(TDW) underMNLP and MNMP treatments
2.5 小麥異附加系的磷吸收能力和利用效率
中氮中磷處理下,小麥異附加系的植株吸磷總量在5.24~30.31 mg之間(表5),磷吸收能力顯著高于中國春的小麥異附加系有9個。中氮低磷脅迫下,小麥異附加系的吸磷總量較正常供磷處理明顯降低,其范圍在0.94~3.06 mg之間;Ag.intermediumB、Ae.longissima2S、Ae.searsii5SS及Ae.umbellulata5U等4個小麥異附加系的吸磷總量顯著高于中國春。Ag.intermediumB和Ae.longissima2S附加系在中氮中磷處理下吸磷能力強,在磷脅迫時(中氮低磷處理)也能維持較高的吸磷量,表明該染色體上可能攜帶有調控磷吸收能力的相關基因。
中氮中磷處理下,小麥異附加系的磷利用效率為64.85~104.20 g TDW·g-1P(表5);與中國春(91.71 g TDW·g-1P)相比,只有L.racemosusH異附加系磷利用效率顯著提高。中氮低磷處理下,同一小麥異附加系的磷利用效率為其在中氮中磷處理下的4倍左右;與中國春(568.38 g TDW·g-1P)相比,L.racemosusl附加系、Ag.elongatum6E附加系的磷利用效率顯著提高;Ag.intermediumF等11個小麥異附加系的磷利用效率顯著降低。
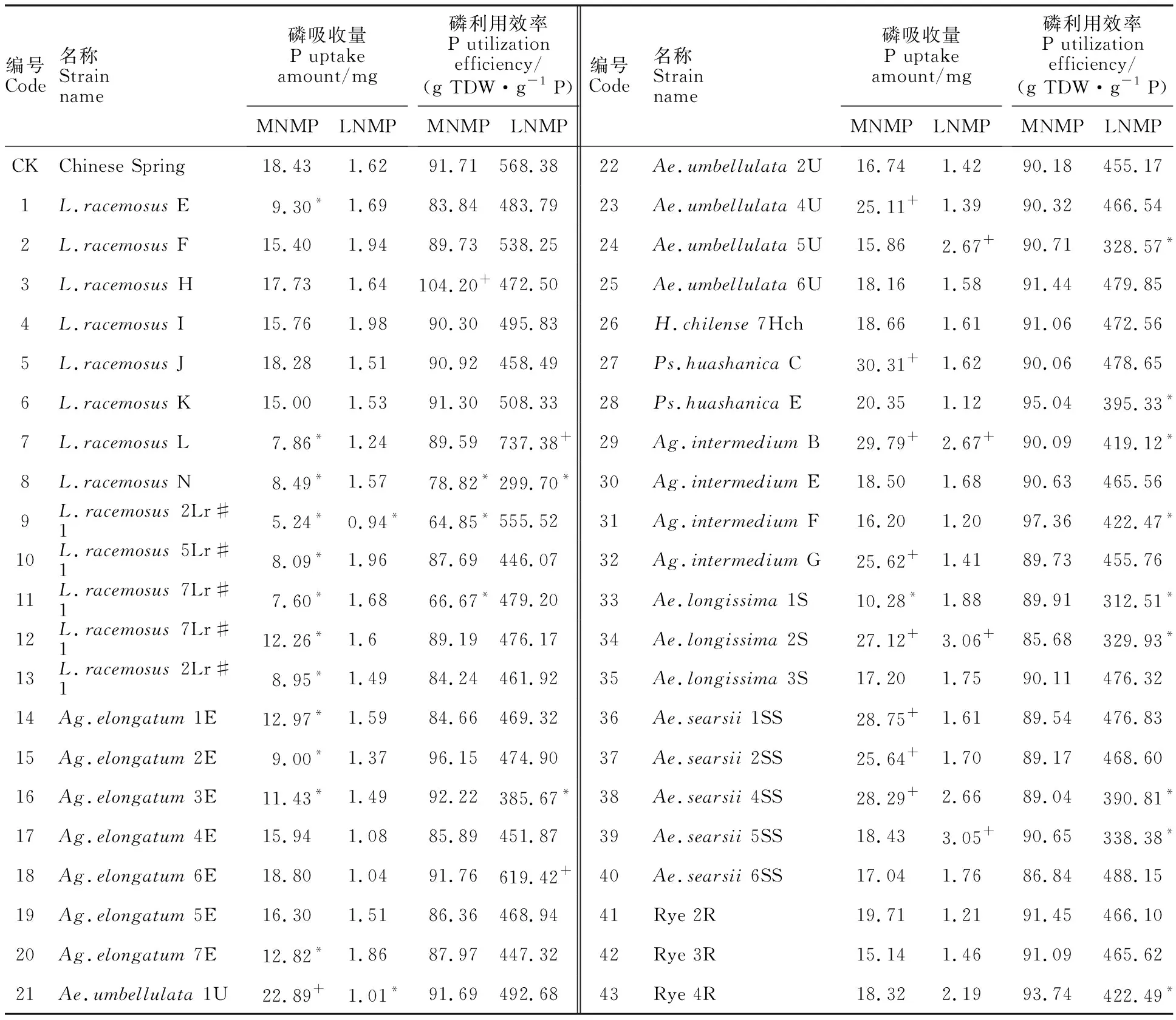
表5 不同處理下小麥異附加系的磷吸收和利用效率Table 5 Prosperous uptake amount and utilization efficiency of wheat addition lines under different treatments
3 討 論
為明確不同氮磷濃度水平對小麥異附加系生長的影響,本研究先對培養4周的小麥植株的苗高、根長、葉數、SPAD值、莖干重、根干重、根莖比等基本形態指標進行了調查,發現低氮脅迫降低了苗期植株的株高、莖干重和總干重,提高了根長、葉數、SPAD值以及根莖比;而磷缺乏提高了根干重和根莖比,抑制了其余性狀的生長;表明氮、磷脅迫均會抑制小麥地上部分的生長,促進地下部分的生長。大量研究表明,SPAD值可以估計葉片單位面積含氮量,且SPAD值大小與氮素水平呈顯著正相關[15-17]。但在本研究中,低氮處理下SPAD值高于正常供氮處理,究其原因,可能是測定時選擇的葉片為幼苗主莖最頂端完全展開葉片,而氮屬于可移動元素,植株缺氮時會從老葉轉移到新葉,優先滿足新葉的生長需求[18];同時,葉色是許多因素綜合影響的結果,當植株缺磷、缺鉀或缺乏微量元素時,葉色會發生變化,此時葉片光譜也會發生改變,影響測定結果;此外,本研究中低氮水平包含了低氮中磷和低氮低磷兩個處理,正常供氮水平包含了中氮中磷和中氮低磷,氮磷互作效應可能也對結果產生了影響。由此可見,在采用SPAD值作為測定氮含量的次級指標時,已完全展開且進入功能盛期的下部葉片可能較上部葉片更適合作為氮素營養的指示葉,且多葉片測定要比單張葉片更可靠[19]。
小麥的近緣種屬作為重要的遺傳資源庫,常被用來向小麥導入新的性狀或者基因,以提高品質,抵御各種生物及非生物脅迫等[4]。由于小麥近緣種屬長期生長在貧瘠的自然環境中,其對肥料的吸收利用能力可能較栽培品種強[5]。要使來源于近緣種屬的有益性狀或者基因能夠在育種中得以應用,首先要保證的是其在普通小麥背景中能夠正常表達和發揮作用。而小麥異附加系,作為攜帶異源染色體的材料,可以用來研究獨立的異源染色體在普通小麥背景中的效應。本研究采用生物量總干重、氮吸收總量、氮利用效率、磷吸收總量、磷利用效率等指標對小麥異附加系的氮、磷效率進行評價,發現供試的小麥異附加系存在豐富的遺傳變異:附加到普通小麥背景中的外源染色體對各測定指標都有一定影響,部分外源染色體的附加可以顯著提高小麥的氮、磷效率,篩選出的部分氮/磷效率較高的種質,可以利用常規雜交或者染色體工程技術,將其氮、磷高效基因導入小麥,從而培育出磷高效且優質高產的品種。
就氮效率而言,趙化田等[10]通過進行耐低氮基因型的苗期水培篩選試驗,發現攜帶冰草、黑麥、偃麥草等外源血統的材料氮利用效率較高;本研究也發現來源于冰草屬的Ag.elongatum6E附加系在低氮環境下表現出較高的氮利用效率,同時還發現來源于華山新麥草屬Ps.huashanica(兩對)和山羊草屬Ae.searsii(1對)的異源染色體也能提高普通小麥在氮脅迫下的氮利用效率,而來源于賴草屬L.racemosus的異源染色體種質相對于其他種質對氮效率有更多不利影響。就磷效率而言,研究發現,黑麥的1R、2R和7R染色體,Ag.elongatum的4E和6E染色體攜帶有耐低磷脅迫的基因[7-8];Ae.peregrina2U、Ae.peregrina3U、Ae.peregrina6U、Ae.searsii4S、Ae.geniculata4M、Ps.huashanicaC、El.trachycaulus5S、Ha.villosa1V和El.trachycaulus7HtS等異源染色體能顯著提高普通小麥的磷效率[9]。本研究同樣發現來自山羊草屬的Ae.umbellulata5U、Ae.longissima2S、Ae.searsii5SS染色體在磷脅迫處理下能顯著提高中國春的磷吸收能力,同時還發現分別來源于賴草屬和冰草屬的L.racemosusL和Ag.elongatum6E染色體能顯著提高中國春的磷利用效率;而不論在磷充足還是磷脅迫處理下,攜帶Ag.intermediumB(No.29)、Ae.longissima2S(No.34)、Ae.searsii2SS(No.37)、Ae.searsii4SS(No.38)和Rye 4R(No.43)染色體的小麥異附加系的總干重總是高于中國春,表現出穩定的磷高效。由于研究材料有限,進一步分析氮、磷高效的小麥異附加系攜帶的異源染色體,沒有觀察到染色體部分同源群聚集的現象。
參考文獻:
[1] KAN T S,BI Y M,ROTHSTEIN S J.Understanding plant response to nitrogen limitation for the improvement of crop nitrogen use efficiency[J].JournalofExperimentalBotany,2010,62(4):1499.
[2] HOLFORD I C R.Soil phosphorus:Its measurement,and its uptake by plants [J].SoilResearch,1997,35(2):228.
[3] 樊龍江,王衛娣,王 斌,等.作物育種相關數據及大數據技術育種利用[J].浙江大學學報(農業與生命科學版),2016,42(1):31.
FAN L J,WANG W D,WANG B,etal.Crop breeding-related data and application of big data technologies in crop breeding[J].JournalofZhejiangUniversity(Agric&LifeSci.),2016,42(1):31.
[4] 張志明,湯才國,楊三維,等.小麥(TriticumaestivumL.) 野生資源的發掘,利用研究進展[J].生物技術進展,2016,6(5):306.
ZHANG Z M,TANG C G,YANG S W,etal.Advance on gene discovery and utilization of wild relatives ofTriticumaestivumL.[J].CurrentBiotechnology,2016,6(5):306.
[5] GORNYA G,GARCZYNSKI S.Nitrogen and phosphorus efficiency in wild and cultivated species of wheat [J].JournalofPlantNutrition,2008,31(2):273.
[6] 柳 鵬,王仕穩,殷俐娜,等.硬粒小麥-粗山羊草人工合成小麥的磷效率研究[J].麥類作物學報,2014,34(3):332.
LIU P,WANG S W,YIN L N,etal.Study on phosphorus efficiency and relative phosiological mechanism of wheat alien chromosome addition lines[J].JournalofTriticeaeCrops,2014,34(3):332.
[7] 李玉京,劉建中,李 濱,等.普通小麥基因組中耐低磷脅迫特性的染色體控制[J].遺傳學報,1999,26(5):535.
LI Y J,LIU J Z,LI B,etal.Chromosomal control of the tolerance to soil phosphorus deficiency in genome of common wheat [J].JournalofGeneticsandGenomics,1999,26:535.
[8] LIU J Z,LI Y J,TONG Y P,etal.Chromosomal location of genes conferring the tolerance to Pi starvation stress and acid phosphatase(APase) secretion in the genome of rye(SecaleL.)[J].PlantandSoil,2001,237:267.
[9] WANG S W,YIN L N,TANAKA H,etal.Identification of wheat alien chromosome addition lines for breeding wheat with high phosphorus efficiency [J].BreedingScience,2010,60(4):371.
[10] 趙化田,王瑞芳,許云峰,等.小麥苗期耐低氮基因型的篩選與評價[J].中國生態農業學報,2011,19(5):1199.
ZHAO H T,WANG R F,XU Y F,etal.Screening and evaluating low nitrogen tolerant wheat genotypeat seedling stage[J].ChineseJournalofEco-Agriculture,2011,19(5):1199.
[11] ZHANG H C,CAO Z H,SHEN Q R,etal.Effect of phosphate fertilizer application on phosphorus(P) losses from paddy soils in Taihu Lake Region:I.Effect of phosphate fertilizer rate on P losses from paddy soil[J].Chemosphere,2003,50(6):697.
[12] KJELDAHL J.A new method for the determination of nitrogen in organic matter[J].AnalyticalandBioanalyticalChemistry,1883.22:12.
[13] TSANG S,PHU F,BAUMM M,etal.Determination of phosphate/arsenate by amodified molybdenum blue method and reduction ofarsenate by S2O42-[J].Talanta,2007,71:1562.
[14] 米國華,劉建安,張福鎖.玉米雜交種的氮農學效率及其構成因素剖析[J].中國農業大學學報,1998,3:99.
MI G H,LIU J A,ZHANG F S.Analysis on agronomic nitrogen efficiency and its components of maize hybrids[J].JournalofChinaAgriculturalUniversity,1998,3:99.
[15] 徐福榮,湯翠鳳,余藤瓊,等.利用葉綠素儀SPAD 值篩選耐低氮水稻種質[J].分子植物育種,2005,3(5):696.
XU F R,TANG C F,YU T Q,etal.Screening of rice germplasm for tolerance to low-nitrogen using SPAD-value by chlorophyll meter[J].MolecularPlantBreeding,2005,3(5):696.
[16] 史力超,翟 勇,王雪艷,等.基于葉片 SPAD 值的滴灌春小麥氮肥分期施用推薦模型[J].干旱地區農業研究,2017,35(1):103.
SHI L C,ZHAI Y,WANG X Y,etal.Recommendation model of nitrogen fertilizer based on SPAD ofleaves for drip-irrigated spring wheat[J].AgriculturalResearchintheAridAreas,2017,35(1):103.
[17] 李 杰,馮躍華,王 旭,等.水稻葉片 SPAD 值分布特征及其與施氮量的關系[J].南方農業學報,2017,48(1):44.
LI J,FENG Y H,WANG X,etal.Distribution characteristics of SPAD value in rice leaf and itsrelationship with nitrogen application rate[J].JournalofSouthernAgriculture,2017,48(1):44.
[18] 朱新開,盛海君,顧 晶,等.應用 SPAD 值預測小麥葉片葉綠素和氮含量的初步研究[J].麥類作物學報,2005,25(2):47.
ZHU X K,SHENG H J,GU J,etal.Primary study on application of SPAD value to estimate chlorophylland nitrogen content in wheat leaves[J].JournalofTriticeaeCrops,2005,25(2):47.
[19] 王紹華,曹衛星,王強盛,等.水稻葉色分布特點與氮素營養診斷[J].中國農業科學,2002,35(12):1462.
WANG S H,CAO W X,WANG Q S,etal.Positional distribution of leaf color and diagnosisof nitrogen nutrition in rice plant [J].ScientiaAgriculturaSinica,2002,35(12):1462.
