氨基酸對小麥成熟胚愈傷組織誘導率的影響
2018-05-11 09:34:08吳一磊孫清栩曲繼鵬彭正松楊在君
麥類作物學報 2018年4期
關(guān)鍵詞:植物
吳一磊,孫清栩,路 璐,余 燕,曲繼鵬,彭正松,楊在君
(1.西華師范大學/西南野生動植物資源保護教育部重點實驗室,四川南充 637009; 2.西華師范大學環(huán)境科學與工程學院,四川南充 637009;3.西昌學院農(nóng)業(yè)科學學院,四川西昌 615013)
小麥傳統(tǒng)育種技術(shù)主要是基于雜交、回交和選種的品種選育過程,其缺點是費時和費力[1]。隨著基因工程和基因組學的發(fā)展,利用現(xiàn)代基因工程技術(shù)對小麥進行定向改造的研究發(fā)展迅速。其中,利用轉(zhuǎn)基因技術(shù)對小麥基因進行定向改變,可得到理想的小麥品種。但小麥轉(zhuǎn)基因研究明顯滯后于水稻和玉米,這在很大程度上歸結(jié)于小麥轉(zhuǎn)化體系的構(gòu)建難度大[2,4]。迄今為止,學者們在進行小麥組織培養(yǎng)的研究中采用了多種不同的外植體材料來進行愈傷組織的誘導,其中包括花藥、幼穗、幼胚和成熟胚等,不同的外植體材料對小麥的組織培養(yǎng)再生率和農(nóng)桿菌侵染效率都會有一定的影響[5]。成熟胚因取材方便、周期短、出愈率高、愈傷組織生長快等特點,已成為外植體的主要來源[6]。但采用成熟胚存在愈傷組織誘導率低的問題。影響愈傷組織誘導率的因素多而復雜,有學者指出基因型是影響愈傷組織誘導率的關(guān)鍵因素,其次是激素類型、含量及培養(yǎng)條件的影響[5,7]。小麥成熟胚愈傷組織誘導及分化的研究目前正處于起步階段,建立高效、穩(wěn)定的成熟胚植株再生體系,對于小麥基因工程育種和基因功能研究具有重要意義。
氨基酸是構(gòu)建生物機體的眾多生物活性大分子之一,是構(gòu)建細胞、修復組織的基礎材料。近20年來,直接以植物為材料的氨基酸生化研究越來越多,對植物氨基酸代謝的認識也更加深刻,這些研究不僅證實了絕大多數(shù)來自動物和微生物的氨基酸代謝過程和原理同樣適用于植物,如天冬氨酸代謝途徑[8]、芳香族代謝途徑[9]等,同時也發(fā)現(xiàn)了不少植物特有的氨基酸代謝途徑和特征[10]。愈傷組織誘導培養(yǎng)基主要使用酸水解酪蛋白作為氮源,但迄今為止尚無學者對各氨基酸成分在成熟胚愈傷組織誘導中的必要性進行討論。本研究結(jié)合酸水解酪蛋白的主要氨基酸成分及其在植物中的作用[11-12],分析了氨基酸缺失對小麥成熟胚的影響,以期找出小麥成熟胚愈傷組織誘導所必須的氨基酸,為優(yōu)化小麥成熟胚再生體系及研究小麥氨基酸代謝的生理過程提供參考。
1 材料與方法
1.1 植物材料
本研究選用的小麥成熟胚種子為2016年收獲的中國春(Chinese Spring,CS),實驗材料種植于四川南充西華師范大學生命科學學院試驗田。
1.2 培養(yǎng)基和處理方法
MS基本培養(yǎng)基作為成熟胚愈傷組織的誘導培養(yǎng)基,并根據(jù)實驗設計添加不同種類的氨基酸。滅菌前調(diào)節(jié)pH到6.5,于108 kPa、121 ℃高壓滅菌鍋內(nèi)滅菌20 min。當溫度降到40~55 ℃時,加入用0.22 μm針頭過濾滅菌的100 μmol·L-1乙酰丁香酮、250 mg·L-1羧芐青霉素、100 mg·L-1頭孢噻肟鈉和2.0 mg·L-1的2,4-D[13-18]。
1.3 培養(yǎng)方法
挑選成熟、飽滿、顏色和大小相對一致的種子,用75%乙醇震蕩滅菌30~60 s,然后用滅菌雙蒸水洗滌5~6次,再用質(zhì)量體積比為30%的過氧化氫浸泡3~5 min,于滅菌雙蒸水中洗滌5~6次后浸泡在雙蒸滅菌水中45 min。用手術(shù)刀對種子進行解剖,暴露出成熟胚,切去部分胚芽后盾片朝上接種于培養(yǎng)皿中暗培養(yǎng)一周(25 ℃,濕度60%),每皿20~25粒種子,當綠色結(jié)構(gòu)出現(xiàn)時,用手術(shù)刀對綠色結(jié)構(gòu)進行切割,并挑出完全白色(偏淡黃色)的愈傷組織,于新的培養(yǎng)基中培養(yǎng)3周以上(新培養(yǎng)基成分與上述培養(yǎng)基成分相同)[19-20],各實驗均重復3次,每次100粒種子。
1.4 數(shù)據(jù)分析方法
出愈率=(愈傷組織塊數(shù)/接種的成熟胚數(shù))×100%。利用SPSS 19.0進行正交實驗設計和相關(guān)數(shù)據(jù)分析。
2 結(jié)果與分析
2.1 滅菌方式及時間對出愈率的影響
利用控制變量法,分別探究酒精和雙氧水的作用效果與作用時間的關(guān)系。當用75%酒精浸泡時間控制在180 s、30%雙氧水滅菌時間在15 s以上時,成熟胚無染菌情況發(fā)生;當30%雙氧水消毒時間控制在180 s、75%酒精浸泡時間在15 s以上時,成熟胚不會出現(xiàn)污染。但隨著酒精浸泡時間延長,酒精對種子造成損害,出現(xiàn)成熟胚出愈率減少、生長狀態(tài)變差、愈傷組織變小等現(xiàn)象;而雙氧水滅菌時間延長對出愈率有微弱的促進作用(P<0.05)(圖1)。
2.2 氨基酸缺失對出愈率的影響
采用酸水解酪蛋白培養(yǎng)液(casein hydrolysate solution,CHS)[13]和對植物有明顯作用的9種氨基酸配制的氨基酸配合液(amino acid combination solution,AaCS)[12](甘氨酸300 mg·L-1、L-天冬酰胺10 mg·L-1、L-谷氨酰胺50 mg·L-1、亮氨酸3 mg·L-1、精氨酸10 mg·L-1、L-脯氨酸50 mg·L-1、絲氨酸50 mg·L-1、酪氨酸50 mg·L-1、L-色氨酸30 mg·L-1),分別對小麥成熟胚愈傷組織進行誘導。結(jié)果表明,CHS的出愈率為93.4%,而AaCS的出愈率為93.5%,二者差異不顯著(P=0.835),說明酸水解酪蛋白與氨基酸混合液在愈傷組織誘導過程中具有相近的功效。
通過控制變量法,分別將缺失1種氨基酸的培養(yǎng)基與含有9種氨基酸的AaCS培養(yǎng)基進行比較(表1),結(jié)果表明,當培養(yǎng)基缺乏甘氨酸、天冬氨酸、脯氨酸和亮氨酸時,成熟胚的出愈率顯著低于對照組,由此可以推測上述4種氨基酸是小麥成熟胚愈傷組織誘導的必需氨基酸。

圖1 滅菌時間對小麥成熟胚出愈率的影響
表1氨基酸缺失對小麥成熟胚出愈率的影響
Table1Effectofaminoaciddeficiencyonthecallusrateofmaturewheatembryo

缺失氨基酸Unaddedaminoacid出愈率Callusrate/%甘氨酸Glycine45±1.41dL?天冬酰胺Asparagine35±1.63eL?谷氨酰胺Glutamine95±0.81a亮氨酸Leucine65±2.16c精氨酸Arginine95±1.63aL?脯氨酸Proline5±1.41f絲氨酸Serine90±2.44ab酪氨酸Tyrosine85±1.63bL?色氨酸Tryptophan85±1.41b對照(AaCS)Controlgroup93.5±2.44a
數(shù)據(jù)后字母不同表示處理間差異顯著(P<0.05)。
Different letters following data meant significant difference among treatments at 0.05 level.
將甘氨酸、天冬氨酸 、脯氨酸和亮氨酸分別記作 A、B、C、D,進行四因素正交實驗,各因素均取3個水平,暫時不考慮其交互作用,選擇L9(34)表進行正交實驗(表2)。
表2L9(34)正交表和小麥成熟胚出愈率
Table2L9(34)Orthogonaltableandthecallusrateofmaturewheatembryo
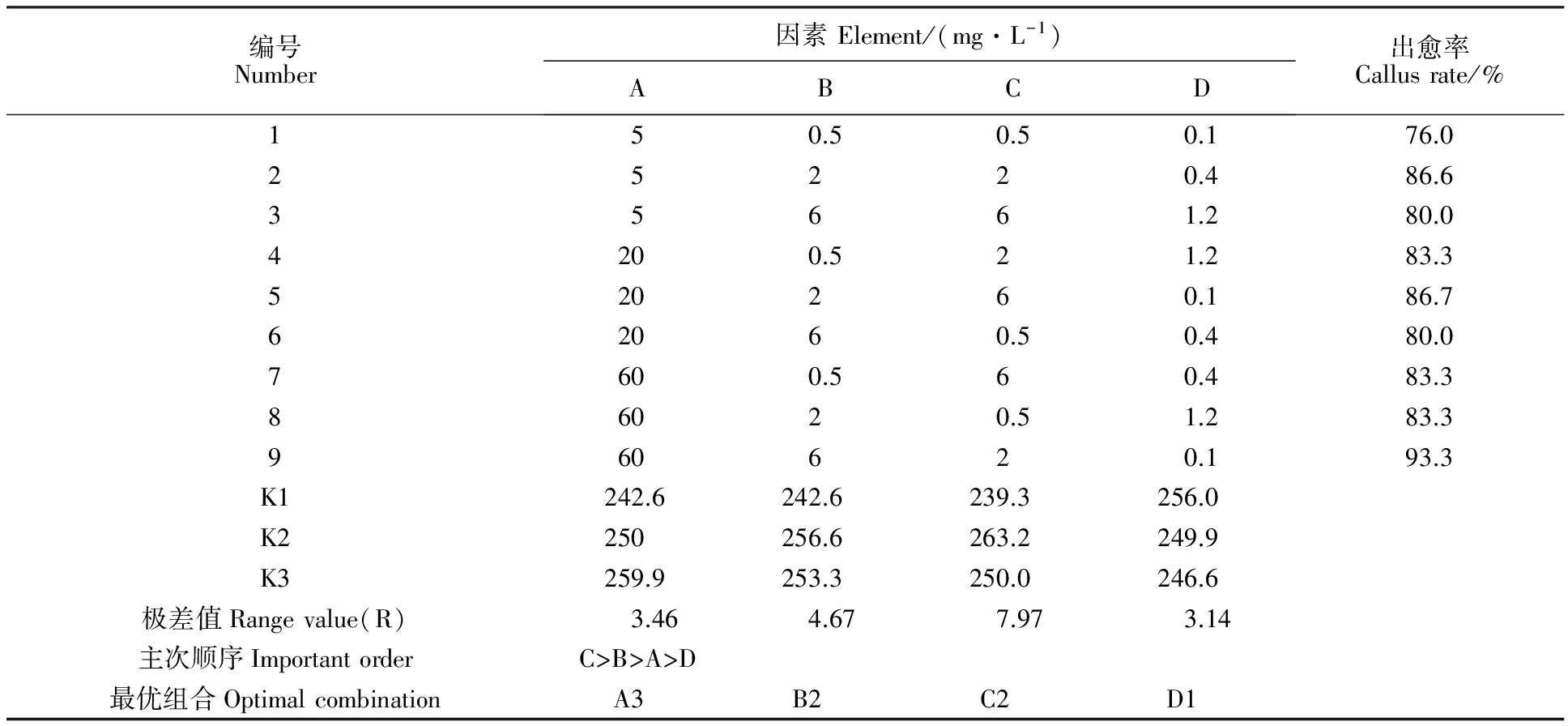
編號Number因素Element/(mg·L-1)ABCD出愈率Callusrate/%150.50.50.176.025220.486.635661.280.04200.521.283.3520260.186.762060.50.480.07600.560.483.386020.51.283.3960620.193.3K1242.6242.6239.3256.0K2250256.6263.2249.9K3259.9253.3250.0246.6極差值Rangevalue(R)3.464.677.973.14主次順序ImportantorderC>B>A>D最優(yōu)組合OptimalcombinationA3B2C2D1
正交實驗結(jié)果如表2所示,影響出愈率的各因素極差大小順序為C>B>A>D,即脯氨酸對出愈率影響最大,其次是天冬氨酸,甘氨酸和亮氨酸對其影響較小。但在設定范圍內(nèi),各個因素的濃度變化對出愈率的影響不大。根據(jù)各因素各水平的出愈率的平均值,確定出最優(yōu)的氨基酸配方組合為A3B2C2D1,即甘氨酸60.0 mg·L-1、天冬氨酸2.0 mg·L-1、脯氨酸2.0 mg·L-1和亮氨酸0.1 mg·L-1。利用該最優(yōu)配方組合,小麥成熟胚的出愈率達可達97% 。圖2顯示了利用最優(yōu)配方組合(圖2A)和利用AaCS(圖2B)培養(yǎng)14 d 的小麥成熟胚出愈情況,可以明顯地看出,利用本實驗優(yōu)化后的配方組合出愈率更高,愈傷組織長勢也更好。

A:利用最優(yōu)配方組合培養(yǎng)結(jié)果;B:利用AaCS培養(yǎng)結(jié)果。
A: Results of culture with optimal combination,B: Results of culture with AaCS.
圖2培養(yǎng)14d后的愈傷組織
Fig.2Callusesofembryoculturedfor14days
3 討 論
小麥種子消毒的研究不僅在小麥成熟胚愈傷組織誘導培養(yǎng)中有重要應用,而且在以小麥種子為基礎的未成熟胚研究和以幼苗等不同部位為外植體的其他研究中也有廣泛的應用。傳統(tǒng)的滅菌是利用100%酒精、10%雙氧水或0.1%升汞處理種子[13,17],通過長時間的浸泡達到對種子表皮滅菌的目的。氯化汞作為最常用的消毒劑,殺菌能力最強,但它卻是一種劇毒物質(zhì),極易進入人體,對眼睛有刺激性,是可疑致癌物。同時,汞污染是一個帶有地方性后果的全球性問題。從實驗人員安全和環(huán)境安全的角度考慮,在小麥種子滅菌的過程中應盡量避免或不使用氯化汞。雙氧水殺菌力強,性質(zhì)不穩(wěn)定,不易殘毒,是使用較多的殺菌劑之一,而且浸泡時間與小麥愈傷組織的誘導率呈微弱的正相關(guān)。因此本實驗采用相對于100%酒精具有更好的滅菌效果的75%酒精和30%雙氧水浸泡小麥種子,大大降低了浸泡時間,減少了對種子的潛在損傷,達到了更好的滅菌效果。
氨基酸在植物信號應激反應和二次代謝中具有重要的作用,不僅為植物生長發(fā)育提供能源或呼吸底物,還能提供合成其他氨基酸的前體物質(zhì),增強植物抗逆性,提高植物滲透壓調(diào)節(jié)能力,有助于組織持水,維持植物高效的生理反應作用,減弱脫落酸的效應,促進植物生長[21]。本研究通過氨基酸篩選實驗發(fā)現(xiàn)甘氨酸、天冬氨酸、脯氨酸和亮氨酸是小麥成熟胚誘導的必須氨基酸。
甘氨酸是唯一能夠同時增加過氧化物酶和苯丙氨酸氨裂解酶活性的氨基酸,能夠參與產(chǎn)生乙醛酸鹽,減少植物中H2O2含量從而降低脂質(zhì)過氧化,并且乙醛酸產(chǎn)生的NADPH和ATP能量分子將用于各種代謝過程[22]。同時,甘氨酸也可以增加植物對磷鉀元素的吸收,增強植物抗逆性。而天冬氨酸是高等植物中賴氨酸、蘇氨酸、甲硫氨酸和異亮氨酸的共同前體,處于轉(zhuǎn)氨基作用的中心地位,是植物中氮的運輸和儲存的關(guān)鍵化合物;此外,在潛在的競爭反應中天冬氨酸也可能轉(zhuǎn)化為天冬酰胺[23]。甘氨酸和天冬氨酸以及脫氨后形成的有機酸是重要的中間產(chǎn)物,轉(zhuǎn)化途徑多,代謝活躍,需要在植物體內(nèi)高濃度積累[21],在愈傷組織誘導早期大量添加甘氨酸和天冬氨酸具有重要意義。而脯氨酸能夠促進植物組織的生長[11],它的生物合成獨立于多胺調(diào)節(jié)和γ- 氨基丁酸生物合成[24],是水溶性最好的氨基酸,具有易于水合的趨勢或具有較強的水合能力,有助于細胞或組織持水,維持滲透壓和生物活性,對于除去胚乳的小麥胚,其持水能力是發(fā)育的巨大難題。由此可見,添加脯氨酸在小麥胚發(fā)育早期的必要性。亮氨酸是植物生長促進劑,而且富含亮氨酸的重復受體樣激酶(LRR-RLKs)在植物生長發(fā)育及應激反應中起重要作用,擬南芥中PnLRR-RLK27的異源表達提高了種子萌發(fā)和早期根生長過程中對鹽、脫落酸及氧化菌的耐受性,增強活性氧清除劑的活性并降低丙二醛和活性氧的水平[25],降低活性氧對幼嫩組織的毒害作用。
正交實驗結(jié)果表明,4種氨基酸的濃度對小麥成熟胚的出愈率影響不大。這可能是因為通過各種氨基轉(zhuǎn)換途徑,氨基酸發(fā)生相互轉(zhuǎn)化[12],所以自身的解釋力被其他控制變量稀釋,出現(xiàn)差異不顯著的現(xiàn)象,但具體原因還需要進一步深入研究。
參考文獻:
[1] SABETTA W,CROSATTI C,SOLTSZ A,etal.Screening auxin response,invitroculture aptitude and susceptibility toAgrobacterium-mediated transformation of italian commercial durum wheat varieties [J].Molecules,2016,21(11):1440.
[2]HIEI Y,OHTA S,KOMARI T,etal.Efficient transformation of rice(OryzasativaL.) mediated byAgrobacteriumand sequence analysis of the boundaries of the T-DNA[J].ThePlantJournal,1994,6(2):271.
[3]ISHIDA Y,SAITO H,OHTA S,etal.High efficiency transformation of maize(ZeamaysL.) mediated byAgrobacteriumtumefaciens[J].NatureBiotechnology,1996,14(6):745.
[4] SINGH R K,PRASAD M.Advances inAgrobacteriumtumefaciens-mediated genetic transformation of graminaceous crops [J].Protoplasma,2016,253(3):691.
[5] CHAUHAN H,DESAI S A,KHURANA P.Comparative analysis of the differential regeneration response of various genotypes ofTriticumaestivum,TriticumdurumandTriticumdicoccum[J].PlantCell,TissueandOrganCulture,2007,91(3):191.
[6] DING L,LI S,GAO J,etal.Optimization ofAgrobacterium-mediated transformation conditions in mature embryos of elite wheat [J].MolecularBiologyReports,2009,36(1):29.
[7] REPELLIN A,B?GA M,JAUHAR P P,etal.Genetic enrichment of cereal crops via alien gene transfer:New challenges [J].PlantCell,TissueandOrganCulture,2001,64(2-3):159.
[8] AZEVEDO R A,ARRUDA P,TURNER W L,etal.The biosynthesis and metabolism of the aspartate derived amino acids in higher plants[J].Phytochemistry,1997,46(3):395.
[9] AZEVEDO R A,LEA P J.Lysine metabolism in higher plants [J].AminoAcids,2001,20(3):261.
[10] 莫良玉,吳良歡,陶勤南.高等植物GS/GOGAT循環(huán)研究進展[J].植物營養(yǎng)與肥料學報,2001,7(2):223.
MO L Y,WU L H,TAO Q N.Research advances on GS/GOGAT cycle in higher plants [J].PlantNutritionandFertilizerScience,2001,7(2):223.
[11] 陳克貴,朱慶麟.脯氨酸在小麥愈傷組織培養(yǎng)中的作用初探[J].植物科學學報,1993,11(1):67.
CHEN K G,WU Q L.Preparation of proline in wheat callus culture [J].ChineseJournalofPlantScience,1993,11(1):67.
[12] 張夫道,孫 羲.氨基酸對水稻營養(yǎng)作用的研究[J].中國農(nóng)業(yè)科學,1984,17(5):61.
ZHANG F D,SUN X.Studies on the nutritional effects of amino acids on rice [J].ChinesAgriculturalSciences,1984,17(5):61.
[13] DING L,LI S,GAO J,etal.Optimization ofAgrobacterium-mediated transformation conditions in mature embryos of elite wheat [J].MolecularBiologyReports,2009,36(1):29.
[14] HIEI Y,ISHIDA Y,KOMARI T.Progress of cereal transformation technology mediated byAgrobacteriumtumefaciens[J].FrontiersinPlantScience,2014,5:1.
[15] PARMAR S S,SAINGER M,CHAUDHARY D,etal.Plant regeneration from mature embryo of commercial Indian bread wheat(TriticumaestivumL.) cultivars [J].PhysiologyandMolecularBiologyofPlants,2012,18(2):177.
[16] SABETTA W,CROSATTI C,SOLTSZ A,etal.Screening auxin response,in vitro culture aptitude and susceptibility toAgrobacterium-mediated transformation of italian commercial durum wheat varieties[J].Molecules,2016,21(11):1440.
[17] SHAH M M,KHALID Q,KHAN U W,etal.Variation in genotypic responses and biochemical analysis of callus induction in cultivated wheat [J].GeneticsandMolecularResearch,2009,8(3):783.
[18] BELLUCCI M,DE MARCHIS F,FERRADINI N,etal.A mutant synechococcus gene encoding glutamate 1-semialdehyde aminotransferase confers gabaculine resistance when expressed in tobacco plastids [J].PlantCellReports,2015,34(12):2127.
[19] MEDVECK E,HARWOOD W A.Wheat(TriticumaestivumL.) transformation using mature embryos [J].AgrobacteriumProtocols,2015,1:199.
[20] ISHIDA Y,TSUNASHIMA M,HIEI Y,KOMARI T.Wheat(TriticumaestivumL.) transformation using immature embryos [J].AgrobacteriumProtocols,2015,1:189.
[21] 楊 靖,孫海燕,李友勇.11 種非必需氨基酸對離體植物生長的脅迫作用[J].生物技術(shù),2008,18(4):72.
YANG J,SUN H Y,LI Y Y.Stress on the growth of plants in vitro of non-essential amino acids [J].Biotechnology,2008,18(4):72.
[22] TEIXEIRA W F,FAGAN E B,SOARES L H,etal.Foliar and seed application of amino acids affects the antioxidant metabolism of the soybean crop [J].FrontiersinPlantScience,2017,8(327):1.
[23] AZEVEDO R A,LANCIEN M,LEA P J.The aspartic acid metabolic pathway,an exciting and essential pathway in plants [J].AminoAcids,2006,30(2):143.
[24] MAJUMDAR R,BARCHI B,TURLAPATI S A,etal.Glutamate,ornithine,arginine,proline,and polyamine metabolic interactions:the pathway is regulated at the post-transcriptional level [J].FrontiersinPlantScience,2016,7(78):1.
[25] WANG J,LIU S,LI C,etal.PnLRR-RLK27,a novel leucine-rich repeats receptor-like protein kinase from the Antarctic moss Pohlia nutans,positively regulates salinity and oxidation-stress tolerance[J/OL].PloSONE,2017,12(2):e0172869.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
